
Abstract
Significance:
The crisis of antimicrobial resistance (AMR) increases dramatically despite all efforts to use available antibiotics or last resort antimicrobial agents. The spread of the AMR, declared as one of the most important health-related issues, warrants the development of new antimicrobial strategies.
Recent Advances:
Antimicrobial peptides (AMPs) and AMP dendrimers (AMPDs), as well as polymer dendrimers are relatively new and promising strategies with the potential to overcome drug resistance issues arising in ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species) colonizing chronic wounds.
Critical Issues:
AMPs–AMPDs suffer from limited efficacy, short-lasting bioactivity, and concerns of toxicity. To circumvent these drawbacks, their covalent coupling to biopolymers and/or encapsulation into different drug carrier systems is investigated, with a special focus on topical applications.
Future Directions:
Scientists and the pharmaceutical industry should focus on this challenging subject to either improve the activity of existing antimicrobial agents or find new drug candidates. The focus should be put on the discovery of new drugs or the combination of existing drugs for a better synergy, taking into account all kinds of wounds and existing pathogens, and more specifically on the development of next-generation antimicrobial peptides, encompassing the delivery carrier toward improved pharmacokinetics and efficacy.

Scope and Significance
Microbial infection has become a major global threat due to the emergence of antimicrobial resistance (AMR). This spurred the need for innovative strategies to fight multidrug-resistant (MDR) bacteria.
This review summarizes the most relevant available antimicrobial agents related to topical therapy. We discuss antiseptics and antibiotics commonly used in wound care and summarize the shortcomings of their application in certain patients. Furthermore, we review innovative strategies relying on linear and dendrimeric antimicrobial peptides (AMPs), including the drug delivery approaches that may improve their efficacy against AMR.
Translational Relevance
Treatment of infections resulting from bacteria belonging to the ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species) collection is a clinical challenge. This article describes commercially available topical antimicrobials and antiseptics, emphasizing their bacterial activity and limitations. Key AMPs tested in clinical trials are presented. To cope with the urgent need for innovative solutions, novel dendrimer peptides, chemical conjugates, and nanocarriers are discussed, which may allow for an improved activity against MDR bacterial infections.
Clinical Relevance
The care and management of infected wounds is a burden not only for patients, who suffer from severe pain, but for clinicians as well. AMR remains a critical issue and novel AMPs and AMP dendrimers (AMPDs) offer solutions for eradicating MDR bacteria, allowing healing to occur.
Background
Skin has essential roles in maintaining homeostasis, preventing microbial invasion, and providing a barrier between body tissues and the external environment. However, burns, and traumatic or chronic wounds, such as venous, diabetic, and pressure ulcers, compromise the protective barrier that skin offers and subsequently facilitate the risk of bacterial infections. 1 Microbial infection due to AMR is one of the major global threats that continues to worsen despite the efforts in finding solutions. Around 0.7 million people die each year due to the emergence and persistence of MDR bacteria or so-called superbugs. 2 It is estimated that by the end of 2050, AMR could lead to more than 10 million deaths annually, compared with more than 8.2 million deaths attributable to cancer. 2
MDR hinders the healing process in wounds, as most of the wounds would develop infections at some point. 1 One of the most challenging MDR bacteria is P. aeruginosa, which is part of the problematic bacterial collection called ESKAPE. Among other virulence factors such as adhesion, quorum sensing, or toxin production, the success of ESKAPE pathogens in escaping from the antibiotic treatment lies in the different posttranslational modifications found in their proteome. 3 With the emergence of multidrug-resistant bacteria and reemerging infectious diseases, development of new antibacterial agents has become an urgent task.
The World Health Organization (WHO) called on member states to act against AMR since 1998. WHO calls for improvement of surveillance and testing systems, appropriate use of antibacterial agents, and strengthening countermeasures against infectious diseases. WHO has recently listed nosocomial ESKAPE among the 12 families of MDR bacteria. Moreover, AMR negatively affects the global economy and especially poor countries where the health care system is underdeveloped. 4 Recent calculations estimate that by the end of 2050, global MDR costs will go above $100 trillion. 2
Clinical and Laboratory Standards Institute (CLSI) guidelines established a list of available antibiotics specifically active against ESKAPE bacteria (Table 1). Surprisingly, many antibiotics and combination of antibiotics have been removed from the list due to their side effects or lack of efficacy. Even for relatively new antibiotics, incidences of resistance are reported and for some bacteria, such as A. baumannii, K. pneumoniae, and P. aeruginosa, resistance has not yet been overcome. These latter were generally kept in the CLSI guideline. Some antimicrobial agents were added since 2010 due to evidence of absence of resistance against specific strains.
Antimicrobial agents against ESKAPE added, removed, or kept from the CLSI guidelines over the period 2010–2019
CLSI, Clinical and Laboratory Standards Institute.
Lastly, AMPs have attracted high interest, since they are less likely to induce MDR. AMPs are short peptides consisting of 10–50 residues and produced by all living forms, including protozoa, bacteria, animals, and humans. They are important mediators of innate immune defense. Their amino acid sequence and secondary structure are diverse, but most of the antibacterial peptides are amphipathic with both basic and hydrophobic clusters, and bacterial cell membranes rich in acidic lipids are used for ATP production. Most AMPs possess cationic properties. They have shown a broad activity against a wide range of pathogens, including MDR bacteria by lysing cell membrane through electrostatic interactions.
However, AMPs are rapidly degraded once in contact with human serum (short plasma half-life), losing their activity and most of them are toxic, besides having high production costs. 16 Another promising approach is the use of AMPDs, which showed better activity than conventional AMPs, but still show quite high toxicity and poor stability in human serum of only a few hours. 17 There is therefore an urgent need to develop new strategies of application of these AMPs and AMPDs avoiding their degradation, while reducing their toxicity at therapeutic concentrations. Chemical conjugation of these potent molecules to different polymers may offer a solution to overcome these drawbacks. Still, the exact antimicrobial mechanism of the polymer–peptide conjugate needs to be further evaluated, as there is a lack of clinical studies describing the healing of infected wounds upon application of AMPs or AMPDs.
Discussion
Infection and biofilm formation
In general, microorganisms colonize all open wounds, although not all wounds will show clinical signs of infection. 18,19 Upon infection, microbes create a cytotoxic environment, which often leads to chronic wounds and eventually to gangrene with successive amputation of the infected limb or even to the death of the patient due to sepsis. 20 However, the likelihood that a wound will be infected is not only related to the presence of the microorganisms, but to the depth, size, and location of the wound, as well. For example, purulent secretions or local expressions of inflammation are clear indications that an infection has occured. 18 Nevertheless, the inflammation may be caused by conditions unrelated to a wound, such as diabetic neuropathy, venous insufficiency, or ischemia. 20
As a rule, wounds can be classified as acute or chronic. Acute wounds are a result of an injury, surgery, or the use of intravascular devices. Acute wounds heal within a very short period of time, following successive phases of inflammation, proliferation, migration of keratinocytes and fibroblasts, and final tissue maturation. 21 Wounds that fail healing within 3 months through the normal healing process are categorized as chronic. 22 This type of wounds show a persistent inflammation phase, which is characterized by a continuous influx of polymorphonuclear neutrophils leading to impaired wound healing. 23
Moreover, as wounds grow deeper and become more complex, they can infect the underlying tissue and bone causing osteomyelitis. 24 For instance, surgical-site infections, such as superficial incisional, deep incisional, organ, or interorgan space infections, will show postoperative signs of infection typically within the first 10 days, occasionally only after month(s). 25
Diabetic wounds, such as foot ulcer or venous ulcer, result from uncontrolled glycemia, leading to microvascular complications (retinopathy, nephropathy, and neuropathy) and very high incidence of infection. 20,26 The potential of the wounds to be infected or to heal depends on the surrounding skin and mucous membranes. Actually, the longer the wounds are exposed to the bacteria, the easier it is for the bacteria to proliferate and colonize.
Wounds are ideal hosts for bacterial colonization, providing a warm environment and nutrients. This may lead to local or dangerous systemic infections. 27 The propensity for a wound to become infected is directly proportional to the pathogenicity or virulence of the microorganism, and inversely related to both local and systemic resistance of the host. 28 Local factors refer to wound size and depth, degree of chronicity, contamination, type of wound, presence of necrotic tissue, anatomic location, and compromised sterilization of the materials; while systemic factors relate to diabetes, obesity, smoking, age, alcoholism, malnutrition, radiation, medication (with steroids, chemotherapy), or inherited neutrophil defects. Usually, infected wounds are accompanied by foul odor, necrotic tissue, wound pain, and impaired healing. It is generally considered that wound colonization occurs at bacterial loads <105 bacteria per gram tissue and infection when >105 bacteria per gram tissue are found. 23
Interestingly, critically ill patients have higher rates of MDR microorganisms compared with other patients. 29 Most common MDR bacteria are methicillin-resistant S. aureus (MRSA), vancomycin-resistant enterococci, and MDR Gram-negative bacteria (i.e., extended-spectrum β-lactamases; AmpC-type β-lactamases, and metallo-β-lactamase). 30,31 Importantly, a rational administration of topical antimicrobial agents should be considered to prevent any resistance development. For this, the use of systemic antibiotics is indicated only when infection is evident or in case of ascending limb infection, sepsis, or incision wounds spreading cellulitis. 32 Bite wounds, depending on their severity, should be treated with oral antibiotics. 33 Care should be taken to limit the duration of antibiotic administration than to the recommended one, to avoid the development of AMR. 34
Bacteria within biofilms are 100–1,000 times more tolerant to antibiotics, disinfectants, or mechanical stresses; thus impeding conventional antibiotic therapy and delaying wound healing in chronic infections. 35 Pathogenic bacteria amplify the AMR issue by creating a 3D bacterial biofilm network, which can strongly enhance the chronicity of the wounds. Biofilms are formed of communities with a high bacterial cell density that are enclosed in a self-produced matrix of extracellular polymeric substance. This matrix composed of exopolysaccharides, proteins, and DNA confers additional resistance to bacteria.
The formation of biofilm (Fig. 1) begins when planktonic (free swimming) cells find their way to a surface to which they attach, followed by their rearrangement to form and maturate the biofilm. Biofilm dispersion, also referred as cell detachment, is followed by release of planktonic bacteria that will restart formation of a new biofilm at distant sites.
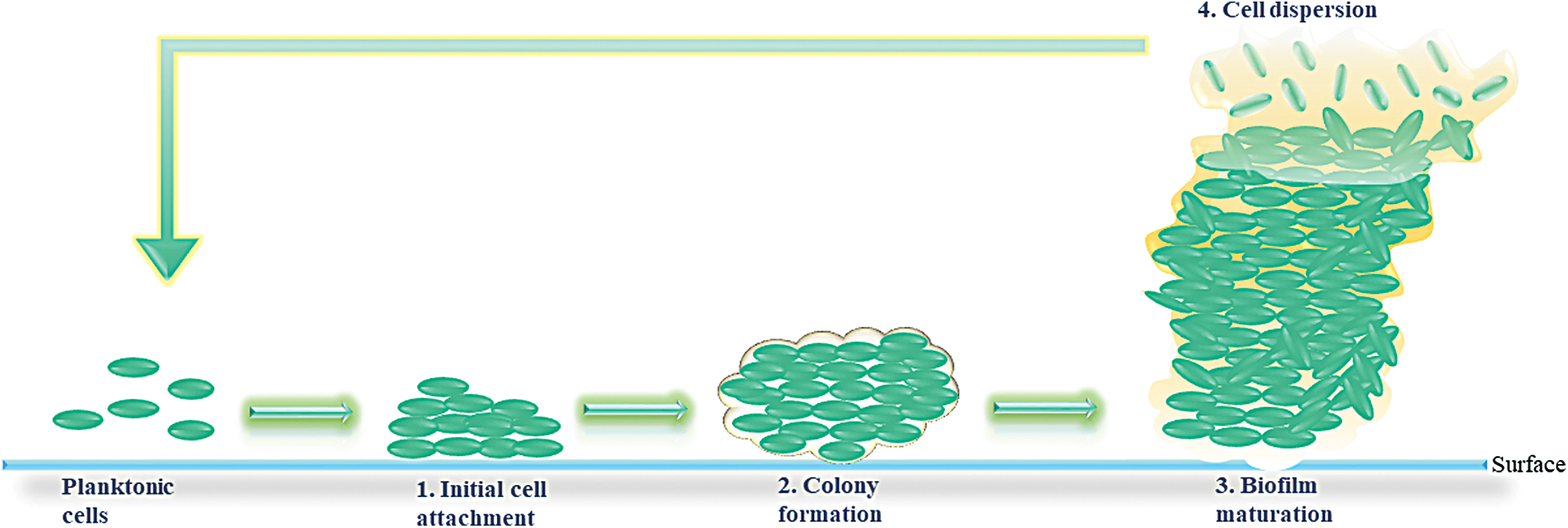
Biofilm formation, including the four stages: (1) Cell attachment, (2) colony formation, (3) mature biofilm, and (4) dispersion of the cells. Color images are available online.
Examples of biofilm-related infections include colonization of almost any surface, including abiotic surfaces (hospital walls, medical devices, implants, catheters, etc.), as well as of biotic surfaces (surgical sites, wounds, lungs, urinary tract, cardiac tissues, bones, etc.). 36
Several mechanisms have been proposed to understand the tolerance of bacteria in biofilms to antibiotic treatment
37
: Limited antibiotic penetration into the biofilm Gene mutation Reduced metabolic rate, growth rate, and division rate Presence of slowly growing persister cells that could reactivate after the antibiotic treatment Overexpression of bacterial efflux pump in biofilm, which leads to increased resistance to antibiotics Protection by the self-secreted matrix of extracellular polymer substance.
There are few antimicrobial agents in clinical use to specifically target biofilms, probably due to the poor understanding of biofilm formation. Resistance ability can be explained if combining the before-mentioned mechanisms. For instance, P. aeruginosa, which frequently leads to biofilm-associated infections, adapts easily to the hostile habitat by producing adapted phenotypes and mutations. On the other hand, the geometry of P. aeruginosa's colonies in a shape of tall ridges or wrinkles (referred as colony rugosity), facilitates their oxygen supply and allow them to grow taller. 35
Current biofilm-related infection diagnosis are based on (i) clinical wound characteristics: edema, erythema, warmth, and purulence; (ii) laboratory-based analyses: microbiological tests of wound swabs; and (iii) technical methods: scanning electron microscopy, gas chromatography–mass spectrometry, epifluorescence microscopy, colorimetric methods, and metabolic and biomass assays. 38 In the context of AMR, adequate diagnosis and design of efficient treatments against biofilm-associated infection is critically needed.
Topical antimicrobial and aseptic agents
Despite recent advances in wound management, very few topical therapies (Fig. 2) proved their efficacy in promoting wound healing. They led to a better understanding of factors influencing the process of wound healing and protection against bacterial infection. However, these methods have met with challenges, as well. 39,40
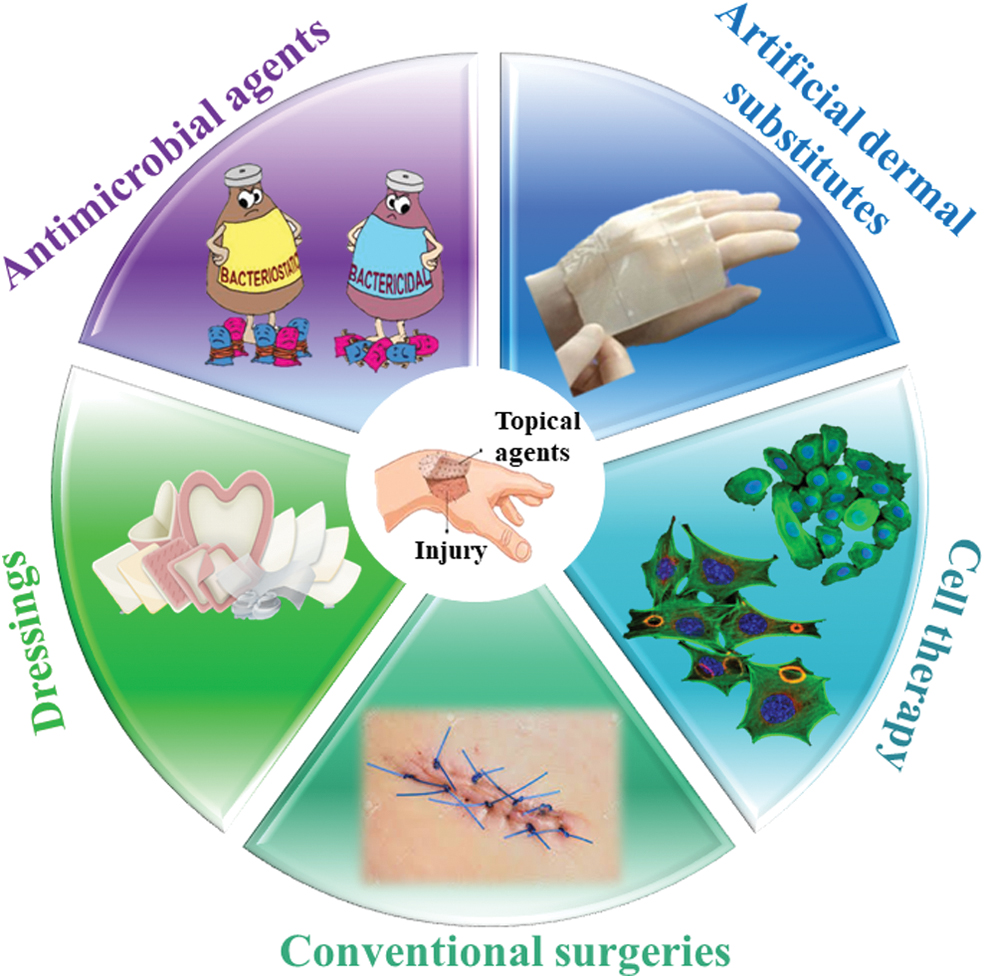
Current treatment methods for skin regeneration. Color images are available online.
Many wound dressings have been developed in an attempt to combine anti-infective properties and promotion of wound healing. Hydrophilic hydrogels and foams absorb wound exudates and keep the wound hydrated. However, they can act as a perfect breeding ground for bacterial growth. Bruises and scrapes result from frequent bandage change, which may lead to new skin injuries. 41 Cotton gauze dressing has been considered for years as the standard of wound care, along with many other commercially available products, such as alginates, collagen dressings, regenerated cellulose, and honey, among others. However, the gold standard for treatment of chronic wounds is yet to be identified. 42 Integration of antibiotics into conventional wound dressings has been of high interest.
Topical antimicrobial agents, including disinfectants, antiseptics, and antibiotics, have the ability to kill microorganisms, inhibit their growth, or reduce their number. Disinfectants are very potent against most microorganisms; however, their high toxicity toward all healthy tissues limits their application to inanimate objects and materials, such as surgical instruments and surfaces. Antiseptics have a broad antimicrobial activity and may be used even for some open wounds, but toxicity was reported as well. 43
In contrast, antibiotics have high bactericidal activity through a specific cell-targeting mechanism—still limited by the AMR, which is very common nowadays. Moreover, administration of some antibiotics is accompanied by several side effects, such as pain, rash, cytotoxicity, toxic effects to kidneys, liver, and other organs. 44 Several studies reported toxic encephalopathy-induced nonconvulsive status epilepticus, 45 seizures, 46 chronic myeloid leukemia, and renal failure, 47 in the case of cephalosporin use.
Chronic wounds are treated using different topical antibacterial or antiseptic formulations with or without antibiotics as listed in Table 2, depending on the severity of the wound. Besides topical agents, treatment with dermal substitutes has shown effective to heal specific wounds. However, these substitutes generally lack the antimicrobial properties required for a durable outcome.
Commercially available topical antimicrobials and antiseptics, their bacterial activity and limitations
+, positive; −, negative; AgNO3, silver nitrate; H2O2, hydrogen peroxide; MRSA, methicillin-resistant S. aureus; PHMB, polyhexa-methylene biguanide; TAA, topical antimicrobials and antiseptics; VRE, vancomycin-resistant enterococci.
Alternative agents to antibiotics
Nowadays, we face an urgent need to identify new antibacterial drugs to overcome AMR of different microorganisms.
Antimicrobial peptides
The AMPs, also referred as host defense peptides, were identified as good candidates to limit resistance-induced microorganisms. They are abundant in prokaryotes (produced by Gram-positive and Gram-negative bacteria) and in all eukaryotic organisms (fungi, algae, plants, insects, and mammals) and well distributed in cells and tissues as the front fighting line against pathogens. 81 The first AMP, a tyrothricin compound was extracted from Bacillus strain by Dubos and Gause in 1939 independently from each other in their respective laboratories. It proved to be effective against pneumococci infection in mice. 82 Soon after, it was found that the first AMP contained two different molecules: 80% tyrocidine and 20% gramicidin. Tyrocidine was very effective against both Gram-positive and Gram-negative bacteria, despite being highly toxic against mammalian cells. 83 Gramicidin was applied for the treatment of infected wounds and especially ulcers during the Second World War.
Inspired by natural AMPs, many synthetic or semisynthetic analogs were recently developed. The main focus is on synthetic AMPs with higher antimicrobial activity and lower risks of toxicity toward host cells than their natural analogs. An online antibacterial peptide database (APD3) lists more than 3,130 AMPs originating from all species, 84,85 out of which 134 are identified as human host defense peptides with more than 100 of those exhibiting antibacterial activity. 86 Their classification depends on the charge, length, sequence of amino acids, and their secondary structure as shown in Table 3. They may have either amphiphilic or cationic domains, for example, human AMPs have a net charge range from anionic (rare) to cationic (most often), which ranges from −3 to +20. 86
The four antimicrobial peptide families with their type of conformation and examples
hBD, human β-defensin; Pg1, protegrin-1.
Considering the critical situation of ESKAPE pathogens, AMPs are used to date as an effective therapy. They have the advantage of fast acting, bactericidal, multifunctional (stimulate the immune system and inhibit bacterial growth), and anti-inflammatory and/or wound healing promotor. AMPs, such as human β-defensins (hBD-1, hBD-2, and hBD-3) 90 and cathelicidin LL-37, 91 originating from epithelial tissues are factors of the innate immune system. They protect skin from infections caused by several microorganisms, such as K. pneumoniae, MRSA, P. aeruginosa, Escherichia coli, and Neisseria gonorrhoeae. 92
Moreover, LL-37 is reported as a safe agent for clinical use as it successfully showed promotion of wound healing in hard-to-heal venous leg ulcers during short-term treatment. 93 Histatin 5, human salivary peptide, has a strong antibacterial activity (≥70%) against five out of six ESKAPE pathogens, except K. pneumoniae. It also showed a strong in vitro antibiofilm formation in P. aeruginosa (60% killing) but less in A. baumannii and S. aureus. 94
Cellular distress is usually noticed when exposing bacteria to antibiotics, although no bacterial adaptation nor resistance development was shown when treating E. coli for several hours with AMPs, such as cecropin A, melittin, magainin II, pexiganan, and LL-37 at 50% minimal inhibitory concentration (MIC). Also, treatment with these AMPs did not show any changes in mutation rate nor differential expression of genes related to stress-induced mutagenesis, while ampicillin, ciprofloxacin, and kanamycin antibiotics increased the mutation rate by threefold to fourfold. 95
Bacterial resistance to AMPs and virulence was noticed in case of AMP proteolytic degradation. For instance, S. aureus together with an aureolysin metalloprotease could degrade LL-37 AMP by cleaving C-terminus bonds of the peptide and in turn contributing to resistance. 95 ZapA metalloprotease could inhibit bacterial activity of LL-37 and hBD-1 against Proteus mirabilis, responsible for urinary tract infections, by at least 7- and 30-fold, respectively. Surprisingly, the same protease did not inhibit the activity of hBD-2, which has differences in amino acid sequence than hBD-1. 96 P. aeruginosa, Enterococcus faecalis, and Streptococcus pyogenes use a common mechanism to escape α-defensins by secreting extracellular proteoglycans, which releases dermatan sulfate. Released compound further binds to α-defensins by completely inhibiting its activity. 95
The process of biofilm formation is another mechanism of AMR to escape AMPs. It was reported that the DNA found in P. aeruginosa biofilm induces resistances to both polymyxin B and colistin by inducing lipopolysaccharide (LPS) modification. 95 On the contrary, no resistance was found when treating protease-resistant P. aeruginosa biofilms with LL-37. 97 In spite of these few reported resistance to pathogens, AMPs remain a promising tool to fight resistance, benefiting from their broad activity spectrum and the variety of their mechanisms of action.
Antimicrobial peptides mechanism of action
Bacteria fall into two main categories depending on their cell wall structure: Gram-negative and Gram-positive bacteria. Gram-positive cells have an outer bacterial cell wall; whereas Gram-negative possess an additional outer membrane adapted with several porins and LPSs. These differences in the cell membrane will confer different susceptibility to various antimicrobial agents. 98 For instance, some of the AMPs were reported to be more effective against Gram-negative than Gram-positive species.
Even though the mechanisms of action are currently debated among scientists, most agree on the key role of electrostatic forces between positively charged AMPs and negatively charged bacterial membrane, which in turns leads to bacterial leakage and death. 99 Other proposed mechanisms are: (i) disruptive, such as “barrel-stave” and “toroidal pore” models of pore formation in the bacterial membrane; “carpet–detergent,” by which peptides can form micelles with the membrane components, (ii) nondisruptive, for example, bacterial membrane thinning, depolarization, or aggregation, and (iii) mediated by the “stringent response,” which is the stress response by the bacteria, involving secondary messenger metabolites. 98 A detailed description of the modes of action is given by Bahar and Ren 100 In addition, the AMP mechanisms of action depend on their concentration, pH, or temperature. 99,101
Antimicrobial peptides toxicity and efficacy: preclinical and clinical data
Despite their high and broad antimicrobial activity, AMPs may suffer from their toxicity toward mammalian cells. Toxicity against red blood cells (RBCs), or the ability of AMPs to lyse RBCs, also referred as hemolysis, is another major concern. Therefore, the selectivity toward bacterial cells is generally defined by the ratio of HC50/MIC, where HC50 is the concentration necessary to lyse 50% of RBCs and MIC is the minimal concentration to inhibit the growth of a given microorganism, for example, to obtain a bacteriostatic effect. 102 Another important parameter is the minimal bactericidal concentration (MBC) of AMPs, which indicates the ability to eliminate (kill) bacteria. Antibacterial agents are usually regarded as bactericidal if the MBC is no more than four times the MIC. 103
For the clinical use of AMPs, one should consider their mechanisms of action, stability under physiological conditions, and the balance between their efficacy and toxicity. Currently, only some AMPs with the ability to combat MDR bacteria have been approved by the FDA and already routinely used, such as Gramicidin (date of approval: 2005), 104 Micafungin (2005), 105 Anidulafungin (2006), 106 Telavancin (2009), 106 Ceftaroline (2010), 5 Dalbavancin (2014), 106 Oritavancin (2014), 106 Caspofungin (2017), 106 Ozenoxacin (2017), 5 Tedizolid Phosphate (2015), 5 and Omadacycline (2018). 5 Most of them are against bacterial infection and administered either I.V. or topically. 107,108 Some AMPs, not yet FDA approved, are being tested in clinical trials and listed in Table 4.
Novel antimicrobial peptides for topical application tested in preclinical and clinical trials to eradicate multidrug-resistant bacteria
hLF, human lactoferrin fragment; HNC, head and neck cancer; I.M., intramuscular; I.T., intrathecal; I.V., intravenous injection; PC, preclinical.
Most of the AMPs cannot reach the clinical phase due to their systemic toxicity, fast degradation, short half-life, and/or reduced activity in the presence of salts or divalent cations. 122 Several AMPs in clinical trials failed to show a better activity than conventional antibiotics or exhibited adverse side effects, although in general one should consider the trade-off between toxicity and efficacy. Some examples are Iseganan (withdrawn after phase III) intended for the treatment of oral mucositis 117,123 and Omiganan (withdrawn after phase III), a topical gel for prevention of catheter infections, acne and rosacea. 122,124 These failures have spurred the development of encapsulation strategies of AMPs into different delivery systems, such as nanoparticles and liposomes, to enhance their stability and half-life.
Besides clinically approved antimicrobial and antiseptic agents, synthetic dendritic polymers and peptide dendrimers have recently shown promising developments, detailed in the dedicated paragraph below. In addition, other approaches such as encapsulation into nanocarriers or chemical coupling to other molecules are used to reduce AMPs toxicity and increase their efficacy.
Polymer dendrimers for topical application
Poly(amidoamine)
Synthetic poly(amidoamine) (PAMAM) dendrimers are available up to the 10th generation and mostly studied for their possible antimicrobial efficacy. The most common polymeric dendrimers are PAMAM, polypropyleneimine, poly-
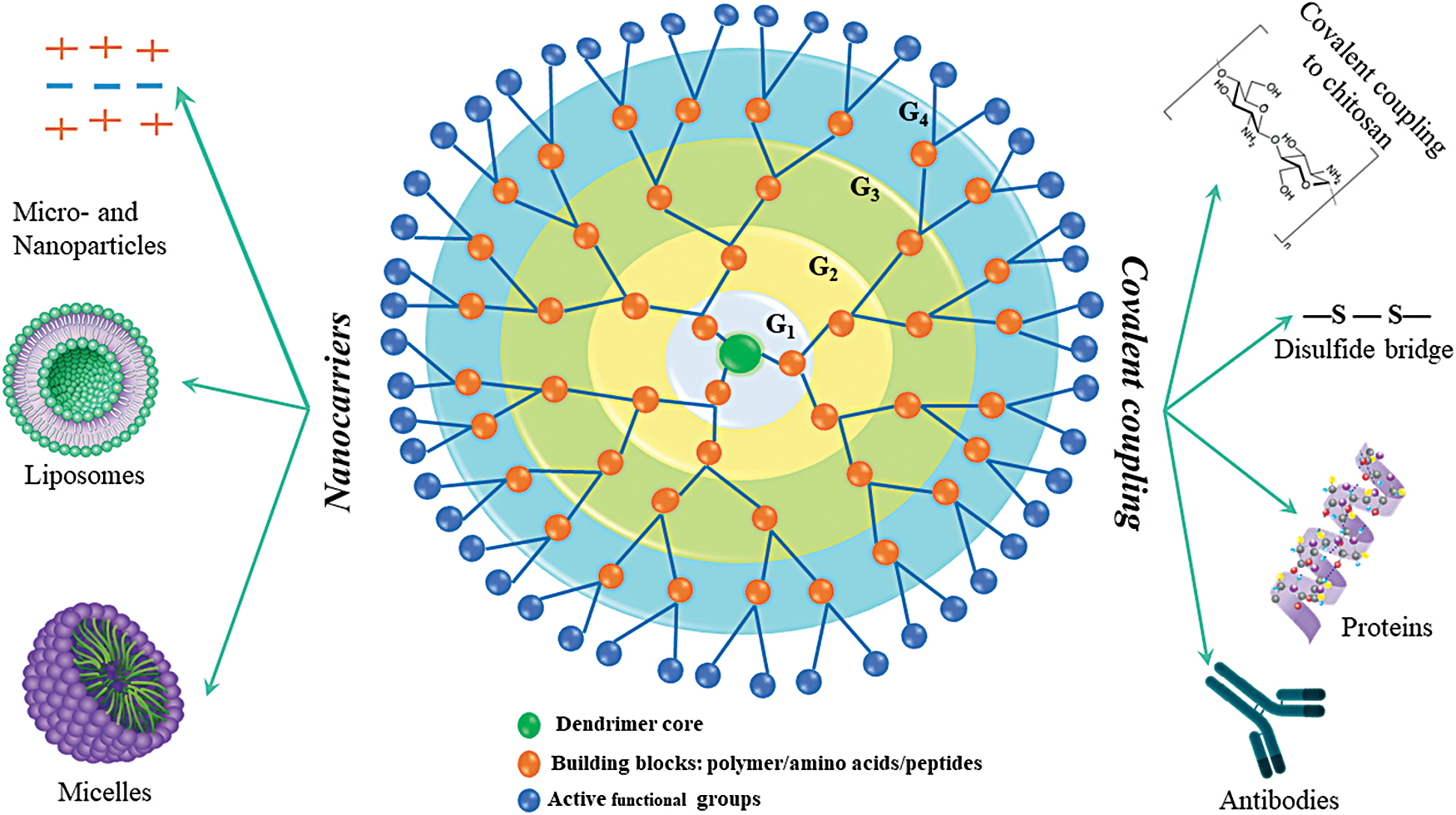
Schematic representation of branched dendrimers and their delivery systems. Color images are available online.
The modification of one unit will affect the properties of the whole dendrimer construct. Calabretta et al. have reported for the first time the effectiveness of fifth-generation (G5) amino-terminated PAMAM dendrimers against both P. aeruginosa and S. aureus at very low concentrations (MIC of 1.5 and 20.8 μg/mL, respectively). 126 However, G5 PAMAM exerted higher toxicity (25% survival at 10 μg/mL) toward human corneal epithelial cells compared with LL-37 (significant toxicity at 25 μg/mL), potentially due to the highly branched cationic dendrimers.
Interestingly, a smaller PAMAM generation (G3) was found to have an enhanced activity against P. aeruginosa and S. aureus when compared with G5 PAMAM (G3: 6.3 μg/mL vs. G5: 12.5 μg/mL), or LL-37 (1.3–12.5 μg/mL). 127 This suggests that the number of amino groups displayed by the dendrimers—higher for G5 than for G3—is not the sole factor for the antimicrobial activity observed.
PEGylation of the functional groups in PAMAM dendrimers was reported to reduce both the toxicity and bactericidal activity, while complete polyethylene glycol replacement of functional groups inhibited the activity of the dendrimer against P. aeruginosa, most probably due to decreased number of amino groups. 126,127 Another approach to reduce the toxicity of PAMAM dendrimers is to modify the functional groups into amino-, hydroxyl-, and carboxyl-terminated G4-PAMAM. 128
The antibacterial activity against E. coli in vitro was found to decrease from G4-PAMAM-NH2 to G4-PAMAM-OH to G4-PAMAM-COOH (IC50 of 3.8 μg/mL, 5.4 mg/mL, and 22 mg/mL, respectively). Topical vaginal and cervical application of G4-PAMAM-OH in a pregnant guinea pig model of chorioamnionitis (intrauterine infection by E. coli) lead to major changes to the outer membrane of E. coli, while G4-PAMAM-NH2 induced changes to both inner and outer bacterial membranes. 128 However, G4-PAMAM-NH2 was dropped for further potential application due to its very high toxicity, while G4-PAMAM-OH was barely transported across placental membrane model, suggesting safety for pregnant women. 129
Lower generation, G1 PAMAM-disaccharide galabiose modified exhibited a 3,000-fold increased potency against Streptococcus suis with an MIC of 0.3 nM. G1 dendrimer was able to inhibit the adhesion of S. suis. 130 Actually, increasing the number of generations in amino-PAMAM from G3 to G7 significantly decreased in vitro viability and inhibited differentiation of human neural progenitor cells and damaged DNA at a concentration of 5 μg/mL. In contrast, G0, G1, and G2 at the same concentration did not show any cytotoxicity. 131 Therefore, lower generation PAMAM hold promise to improve the efficacy–toxicity ratio of the dendrimers, paving the way to clinical applications.
AMPDs for topical applications
Compared with linear AMPs, AMPDs show a three-dimensional, regularly branched structure built by covalent bonds, a very low polydispersity and a higher density of surface groups. 17 Their structure is very similar to the polymeric dendrimers, except that they have only one-side branches, which makes them more flexible for chemical coupling or incorporation into different delivery systems (Fig. 4). The synthetic flexibility and high density of the functional groups found in AMPDs make them very attractive for use as delivery systems for drugs and bioactive principles. Designing peptide-based agents is strongly supported by the high potency of the AMPDs not only to kill bacteria, but also to reduce the toxicity against mammalian cells. 132
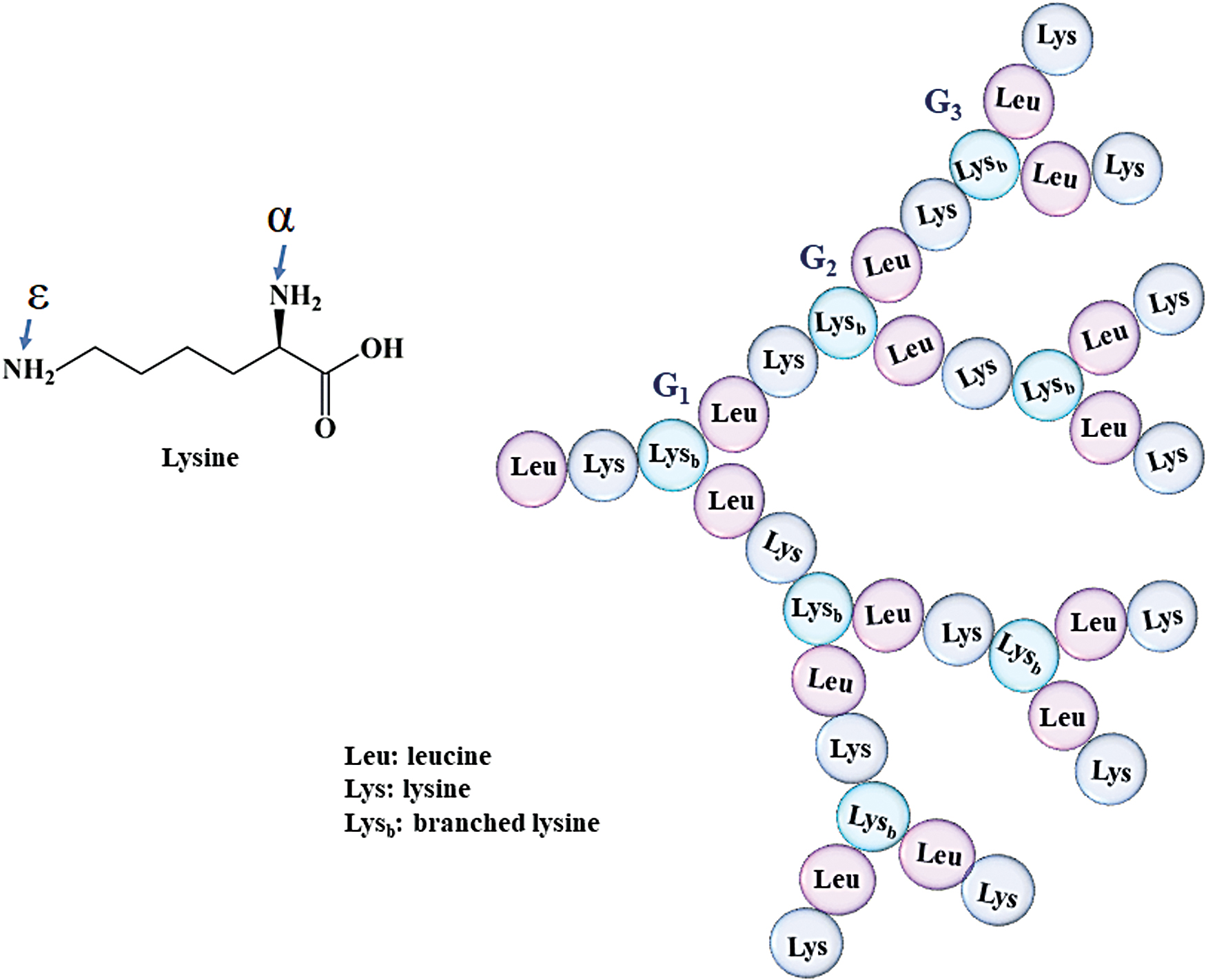
Amino acid sequence in AMPD (e.g., G3KL), which is based on a divalent lysine core whose α and ɛ amines along with leucine double geometrically with each ramification building up a new generation. AMPD, antimicrobial peptide dendrimer. Color images are available online.
The displayed functional groups will govern the mechanism of bacterial killing. AMPDs, which bear charged ends, are believed to act by penetrating the cell membrane inducing leakage of intracellular materials resulting in bacterial death. Therefore, the AMPD mechanism of action against bacteria is related to the number of functional surface groups and their ability to cross the cell membrane. 132
AMPDs show increased activity, which is usually attributed to the higher local concentration of bioactive units in such assemblies, and to their greater stability against peptidases and proteases. For instance, dendrimeric peptides were shown to be selective for microbial surfaces with a broad antimicrobial and low hemolytic activity. A family of AMPDs based on R4 tetrapeptide (RLYR) and R8 octapeptide (RLYR-KVYG), were tested against 10 different microbial strains. Both R4 and R8-based dendrimers of fifth and eighth generation exhibited high activity with MICs <1 μM against Gram-positive and Gram-negative bacteria as well as fungi. 133
Besides, these AMPDs were resistant to proteolytic degradation or to protease inhibition, which has been attributed to their dendrimeric structure. A lipodimeric peptide, SB056, was investigated for its antimicrobial activity against a range of bacteria, including Gram-positive and Gram-negative strains. The in vitro assays showed high antimicrobial activity with MIC in the range of 2–32 μg/mL against A. baumannii, Enterobacter cloacae, E. coli, K. pneumoniae, and P. aeruginosa, which is comparable with the activity of polymyxin B.
The SB056 AMPD showed strong activity against E. coli and S. aureus strains as well as strong Staphylococcus epidermidis biofilm inhibition. 134 Further improvements on the amphipathic part of the SB056 resulted in more ordered β-strands with a stronger antimicrobial activity against both Gram-positive and Gram-negative bacteria. 135 A study on series of tryptophan-ending dendrimers showed that amphiphilic AMPDs can be an effective therapy of E. coli infections. Most of the tryptophan-anchored AMPDs were able to inhibit the growth of antibiotic-resistant E. coli strains, sometimes better than polymyxin B or even indolicidin, besides showing stability in plasma along with low hemolysis and genotoxicity. 136
Recently, a novel G3KL (containing repetitive units of lysine and leucine) AMPD showed high potency at low MIC against 35 strains of P. aeruginosa (8–32 μg/mL), 32 strains of A. baumannii (16 μg/mL), E. coli (8 μg/mL), and K. pneumoniae (16–64 μg/mL). 17 G3KL is a peptide dendrimer of third generation, which acts as a membrane-disrupting agent against bacteria. G3RL (with repetitive units of arginine and leucine) showed lower efficiency than G3KL against P. aeruginosa (8–32 μg/mL) and Bacillus subtilis (11 μg/mL). Once in contact with the serum, the biological activity of G3RL is inhibited. 132
Moreover, both G3KL and G3RL within biological bandages have shown high efficacy against P. aeruginosa, absence of toxicity, and no gene alteration in progenitor fibroblast cells at a concentration of 100 μg/mL. Especially G3KL showed enhanced angiogenesis in human umbilical vein endothelial cells and chorioallantoic membrane assays, as a proof of further potency to enhance wound healing. 137 A second-generation AMPD, such as TNS18, has the same activity as G3KL against Gram-negative bacteria, except K. pneumoniae and against Gram-positive MRSA (MIC = 8–16 μg/mL). 17
Moreover, D-enantiomeric dendrimers dG3KL and dTNS18 have shown high killing effect against different P. aeruginosa biofilm strains (90.2–100%) in vivo on larvae. 138 Therefore, the topology and the sequence of the dendrimers can not only affect their antimicrobial potential, but can also alter their proangiogenic effect, as well. Moreover, the same group of Reymond have developed two different glycopeptide dendrimers: a fucosylated peptide dendrimer (FD2) and two galactosylated dendrimer (GalAG2 and GalBG2), which proved to be potent against P. aeruginosa biofilm formation in vitro. 139
The AMPDs show strong potency against multiple bacterial strains and biofilms. Further research is warranted to optimize their delivery to the wounded site, for a potential clinical translation.
Delivery strategies for AMPs
The limitations of AMPs in terms of efficacy, fast degradation, or toxicity require adequate delivery strategies to tackle these challenges. As AMPDs are a relatively new class of antimicrobials, most of the research has been performed on AMPs, which were covalently coupled to biopolymers or encapsulated into nanoparticles or liposomes. The design of AMP nanocarriers could serve as an example of how to render AMPDs even more effective while preserving their bioactivity.
Covalent coupling of AMPs to chitosan–chitosan derivatives
AMPs can be favorable drug candidates to be coupled to biopolymers, such as chitosan or chitosan derivatives to reduce hemolytic effects and/or enhance antibacterial activity, also benefiting from chitosan's bacteriostatic properties. These conjugates have the advantage of increased stability against proteases and peptidases, low immunogenicity, high efficiency and selectivity, and relatively small size that allows AMPs to disrupt bacterial wall. 140
Anoplin–chitosan: Anoplin (derived from wasp venom) covalently coupled to chitosan showed enhanced in vitro bioactivity and absence of hemolysis. The activity of anoplin–chitosan conjugates against S. aureus and E. coli increased proportionally with their degree of substitution (MIC of anoplin peptide of 1.9 μg/mL against E. coli). 141
HHC10–chitosan: Cysteine-HHC10 AMP coupled to chitosan showed enhanced bioactivity against S. aureus and S. epidermidis; almost no hemolysis and lower toxicity than HHC10 alone. 142
hLF1–11–chitosan: Human hLF1–11 covalently coupled to a thiol-derivatized chitosan film lead to a significant increase in S. aureus adhesion against implant-related infections. 143
Dhvar-5–chitosan: This peptide was immobilized to chitosan films for S. aureus elimination. 144
These studies suggest a potential for chitosan–peptide conjugates to improve activity and decrease toxicity compared with the parent peptide. Still, we need further investigations to validate this experimental approach and reveal the mechanisms behind this improvement.
Nanocarrier systems
Encapsulation of peptides into nano- or microcarriers systems can be an efficient approach to lower cytotoxicity, preserve activity by reducing their degradation and enhance their selectivity. 145 Among these, liposomes, micelles, polymer nanoparticles, and microparticles have met success for drug delivery. This approach has been applied to some AMPs, potentially improving their pharmacokinetic profile.
Liposomes
Liposomes are self-assembled colloidal systems composed of one or more phospholipid bilayers. They have been studied in the last decades as suitable vehicles for drug delivery due to their encapsulation ability and biocompatibility. 145 Liposomes have the advantage of encapsulating both hydrophobic and hydrophilic compounds. Moreover, these drug release systems may protect AMPs against degradation, decrease cytotoxicity, and enhance their stability and bioactivity. For example, Yang et al. incorporated a WLBU2 peptide (24-amino acids) using a modified liposome delivery system with high efficiency against both Gram-positive and Gram-negative bacteria (P. aeruginosa and S. aureus, respectively) and against Chlamydia trachomatis.
Furthermore, WLBU2-modified liposomes were safe to human skin fibroblasts and the activity of the peptide was preserved even in the presence of human serum and blood. 146 This AMP-modified liposome system could be potentially used for local infections. An I.V. injection of tuftsin-loaded liposomes in infected animals resulted in site-specific delivery of AMP and stimulation of liver and spleen macrophage functions against antibacterial–antiparasitic infections, such as tuberculosis and leishmaniasis. 147
Vancomycin and chitosan-loaded liposomes were shown to not only improve the pharmacokinetic profile of the peptide, but to also reduce nephrotoxicity in mice. Injectable vancomycin liposomes showed high antibacterial efficacy against Gram-positive bacterial infections, a sustained release profile, and prolonged systemic circulation. 145 This prevented a vancomycin burst release, which could lead to different side effects.
Micelles
A DP7 (12-amino acids) cationic and hydrophilic AMP, incorporated into a micellar system, showed potent therapeutic benefits in different in vivo disease models and proved to be safe through I.V. injection in mice. The AMP–micellar construct showed reduced hemolysis and high antibacterial activity against S. aureus both in vitro (MIC of DP7-micelles >1,024 μg/mL against P. aeruginosa, S. aureus, and E. coli) and in vivo.
In P. aeruginosa-infected zebrafish embryos and S. aureus-infected mice, DP7-micelles showed high efficacy and therapeutic safety comparable to vancomycin. After an I.V. (80 mg/kg body weight) administration of DP7-micelles, all mice survived and no liver bleeding or pulmonary hemorrhage was observed. 148 These AMP-micelle formulations may potentially be used for bacterial infections (in both Gram-positive and Gram-negative species) as they were shown to significantly stimulate defensive immune reactions in vivo as well.
Micro- and nanoparticles
Vancomycin has been loaded into polycaprolactone polymer microparticles to minimize the side effects of vancomycin. These microparticles were coadministered with calcium phosphate bone substitutes for preventing postsurgery infection. The encapsulation of vancomycin into microparticles resulted in prolonged peptide release in vitro over several weeks. 149 Vancomycin was successfully encapsulated into poly(lactide-co-glycolide) (PLGA) polymers to prevent external–internal ocular bacterial infections. 150 Thus, vancomycin encapsulated into nanodelivery systems may successfully be used as an alternative treatment of infections caused by MDR bacteria.
Piras et al. could formulate an efficient nanoparticles system against S. epidermidis by ionic gelation method. Peptide LLPIVGNLLKSLL-amide (called TB) was added to chitosan to form NPs. The encapsulation of TB peptide exhibited high bactericidal properties against S. epidermidis strains and significantly reduced the toxicity against mammalian cells. 151 Another RBRBR peptide was encapsulated into chitosan NPs by a similar method resulting in an enhanced activity against S. aureus and significantly reduced hemolysis and cytotoxicity. 152 Thus, chitosan may act as an antimicrobial activity enhancer and/or significantly limit the toxicity of the AMPs.
d'Angelo et al. engineered a colistin-loaded PLGA nanoparticles system for sustained delivery of the peptide against P. aeruginosa in lung infection. Colistin encapsulated into PLGA NPs could easily penetrate an artificial mucus layer during the first 6 h and successfully eliminated P. aeruginosa biofilm in vitro within 72 h at 7.5 and 15 μg/mL. 153
These few studies of AMPs covalently coupled or noncovalently associated to different biopolymers suggest that AMPs are potent candidates to eradicate MDR bacterial infections at an enhanced antimicrobial activity and lower toxicity. Additionally, achieving optimum drug–AMP loading, using the right and safe polymer, storing the new formulation while preserving the bioactivity and stability of the AMP are definitely to be further evaluated.
Summary
The occurrence of AMR has changed the landscape of the drugs used in clinics, more specifically to treat ESKAPE-related infections. In addition, the complexity of the factors affecting wound healing renders the choice of adequate antimicrobial agents difficult. In this context, alternative strategies to overcome AMR are proposed.
Besides clinically approved antimicrobial and antiseptic agents, synthetic dendritic polymers and novel peptide dendrimers (AMPDs) have recently shown promising results in preclinical models of infection. Further strategies are also available to improve their activity and decrease toxicity compared with the parent peptides: the conjugation with (bio)polymers, or the incorporation into carriers such as liposomes, nano- or microparticles. These strategies may allow for a sustained pharmacokinetic profile and improve the activity against MDR bacterial infections, paving the way toward their use in a clinical setup.
Take-Home Messages
The occurrence of AMR has changed the landscape of the drugs used in clinics, more specifically to treat ESKAPE-related infections.
Antimicrobial peptidic agents are highly potent with a broad activity against Gram (+) and (−) bacteria and microorganisms.
AMPs are good candidates to limit resistance-induced microorganisms, benefiting from their broad activity spectrum and the variety of their mechanisms of action.
The performance of AMPs can be further enhanced by several strategies such as: chemical conjugation to biopolymers or organization in a dendritic structure.
Nanocarrier technology may further improve pharmacokinetic profile to increase antimicrobial effect and reduce toxicity.
AMPDs show strong potency against multiple bacterial strains and biofilms. Further research is warranted to optimize their delivery to the wounded site, for a potential clinical translation.
Footnotes
Acknowledgment and Funding Sources
No external funding sources were used in the preparation of this article.
Author Disclosure and Ghostwriting
All authors confirm no conflict of interest and no ghost writers were used to write this article.
About the Authors