
Abstract
Traumatic brain injury (TBI) causes significant neurophysiological deficits and is typically associated with rapid head accelerations common in sports-related incidents and automobile accidents. There are over 1.5 million TBIs in the United States each year, with children aged 0–4 being particularly vulnerable. TBI diagnosis is currently achieved through interpretation of clinical signs and symptoms and neuroimaging; however, there is increasing interest in minimally invasive fluid biomarkers to detect TBI objectively across all ages. Pre-clinical porcine models offer controlled conditions to evaluate TBI with known biomechanical conditions and without comorbidities. The objective of the current study was to establish pediatric porcine healthy reference ranges (RRs) of common human serum TBI biomarkers and to report their acute time-course after nonimpact rotational head injury. A retrospective analysis was completed to quantify biomarker concentrations in porcine serum samples collected from 4-week-old female (n = 215) and uncastrated male (n = 6) Yorkshire piglets. Subjects were assigned to one of three experimental groups (sham, sagittal-single, sagittal-multiple) or to a baseline only group. A rapid nonimpact rotational head injury model was used to produce mild-to-moderate TBI in piglets following a single rotation and moderate-to-severe TBI following multiple rotations. The Quanterix Simoa Human Neurology 4-Plex A assay was used to quantify glial fibrillary acidic protein (GFAP), neurofilament light (Nf-L), tau, and ubiquitin carboxyl-terminal hydrolase L1 (UCH-L1). The 95% healthy RRs for females were calculated and validated for GFAP (6.3–69.4 pg/mL), Nf-L (9.5–67.2 pg/mL), and UCH-L1 (3.8–533.7 pg/mL). Rising early, GFAP increased significantly above the healthy RRs for sagittal-single (to 164 and 243 pg/mL) and increased significantly higher in sagittal-multiple (to 494 and 413 pg/mL) groups at 30 min and 1 h postinjury, respectively, returning to healthy RRs by 1-week postinjury. Rising later, Nf-L increased significantly above the healthy RRs by 1 day in sagittal-single (to 69 pg/mL) and sagittal-multiple groups (to 140 pg/mL) and rising further at 1 week (single = 231 pg/mL, multiple = 481 pg/mL). Sagittal-single and sagittal-multiple UCH-L1 serum samples did not differ from shams or the healthy RRs. Sex differences were observed but inconsistent. Serum GFAP and Nf-L levels had distinct time-courses following head rotations in piglets, and both corresponded to load exposure. We conclude that serum GFAP and Nf-L offer promise for early TBI diagnosis and intervention decisions for TBI and other neurological trauma.
Introduction
Traumatic brain injury (TBI) is a common outcome in both sports-related incidents and automobile accidents, typically associated with rapid head accelerations, and leading to significant neurophysiological impairments such as cognitive, motor, and sensory dysfunctions. 1 It is estimated that there are more than 1.5 million cases of TBI in the United States each year. 2 Children aged 0–4 are particularly vulnerable to TBI with increased rates of brain injuries compared to adults and adolescents, 3 and TBI is the leading cause of morbidity and mortality in the pediatric population with nearly 100,000 fatalities in children and adolescents aged 0–19 years between 1997 and 2017. 4 TBI diagnosis is achieved through interpretation of clinical signs and symptoms using various neurophysiological assessments and neuroimaging modalities (e.g., computed tomography [CT] scans). 5 Acute TBI diagnosis and prognosis presents a significant challenge in young children who cannot express symptoms as effectively as adults. 6 Neuroimaging focuses on the more severe brain injury cases, is costly, and can be time consuming, 7 and avoiding ionizing radiation such as from CT scans is especially crucial in children, who experience increased risk of developing cancer and neurodegenerative diseases. 8,9 Therefore, there is increasing interest in minimally invasive fluid biomarkers to objectively detect and/or screen for follow-on diagnostic methods 10 and offer insight into the molecular and cellular changes associated with trauma across the injury severity spectrum. 7,11 The pediatric population is understudied, so biomarker levels in the developing stages of life are of particular interest, as many blood-based biomarkers increase with age. 12 –14 In contrast, GFAP has a U-shaped curve with age, with higher concentrations at <10 and >60 years old. 14
Serum biomarkers have significantly enhanced diagnostic and therapeutic approaches across various fields, including hematology, cardiology, oncology, and infectious diseases, 15 and the advent of immune assays in the last decades has enabled robust measurement of protein biomarkers from blood samples. 16 Numerous brain-specific proteins found in circulation have surfaced in the literature as potential biomarkers for TBI, and given the diverse cellular composition of the brain, measuring multiple biomarkers helps quantify the comprehensive spectrum of heterogeneous pathobiology associated with TBI. 17 For instance, glial fibrillary acidic protein (GFAP) is a filament protein exclusively found in astroglia in the brain; 18 neurofilament light (Nf-L) and tau are cytoskeletal proteins predominately found in myelinated and unmyelinated axons; 19 and ubiquitin carboxyl-terminal hydrolase L1 (UCH-L1) is an abundant, ubiquitinating enzyme predominately in neuronal bodies. 20 Multiplex immunoassays offer a pivotal leap forward enabling quicker, cost-effective measurement of biomarkers, smaller sample volumes, and development of objective TBI assessment scores. 17,21 The U.S. Food and Drug Administration (FDA) has granted approval for use of GFAP and UCH-L1 to rule out the need for head CT scans for patients with a suspected head injury, applying a conservative approach achieving a high sensitivity predicting a positive CT (97.5%) but limited specificity (36.4%). 22 In a cohort of TBI patients, there were differences in GFAP and UCH-L1 levels between CT+ and CT− patients achieving an AUC >75%; 23,24 however, the distinctive temporal profiles for both biomarkers over a span of 7–14 days emphasize the need to understand the time-courses and utility of blood-based biomarkers in the complex phenotypes and causes of TBI.
Human studies are challenged by participation attrition and obtaining blood samples at target time points. Pre-clinical models offer controlled conditions to evaluate TBI with known biomechanical conditions and standardized timing for sample collection. Swine serve as an exceptional large animal model for pediatric TBI, owing to their neuroanatomical similarities to humans with comparable structure and organization. 25,26 In addition, piglet brains exhibit developmental similarities to the pediatric brain in myelination timeline and white matter volume, aiding researchers in studying neurological conditions throughout development in both species. 27,28 Rapid nonimpact head rotation (RNR) models the swift acceleration and/or deceleration experienced by the head during falls or collisions and effectively reproduce the biomechanical mechanisms that cause TBI and resulting neuropathology. 25,29 The RNR TBI model of pediatric swine provides an optimal platform with which to study the time-course of blood biomarkers and relationships to severity of injury.
In summary, the objective of the current study was twofold: first, to establish healthy reference ranges (RRs) of specific biomarkers associated with glial, axonal, and neuronal injury—namely GFAP, Nf-L, UCH-L1, and tau—in 4-week-old Yorkshire porcine serum; and second, to analyze the acute time-course following a RNR injury in piglets. We hypothesize that the time-course in biomarker alterations after head rotation mimics those documented in humans after TBI. Repetitive head loading in sports is associated with long-term neurological effects and increased risk of concussion, 30,31 and the timing and magnitude of repeated head loading influence clinical and pathological outcomes. 32 We hypothesize that repetitive head loading will result in larger alterations in biomarker levels than single head rotation loads. Finally, unlike many common blood tests for general health, healthy RRs for TBI biomarkers have not been established; healthy RRs could be used for TBI screening in the absence of baseline testing or control groups. We hypothesize that healthy RRs for the 4-week-old Yorkshire porcine population can be established and that biomarker levels after injury are significantly altered from the healthy RRs and have reproducible time-courses which affect clinical utility for diagnosis.
Materials and Methods
A retrospective analysis was completed to quantify biomarker concentrations in porcine serum samples collected from 2011 to 2022. Animal subjects received experimental RNR, centered on the cervical spine, or were allocated to an anesthetized sham group. All protocols were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania (2011–2018) or Emory University (2019–2022).
Animals
Four-week-old female (n = 215) and uncastrated male (n = 6) Yorkshire piglets, whose level of brain development and myelination corresponds to 2- to 4-year-old human toddlers, 33 were used. Animals were received in cohorts of two to three littermates and socially housed for the duration of the study, with a 12-h light/12-h dark cycle and ad libitum access to pellets and water.
Study design
Animals and corresponding serum samples were selected through a convenience sample of available frozen serum (stored at −80°C) spanning various collection timepoints across distinct sham/RNR paradigms. Specifically, animals were categorized into one of the following experimental groups: sham, sagittal-single, sagittal-multiple, or baseline only (Table 1). Pre-clinical and physiological data from these animal subjects have been published in numerous articles for over a decade, but the serum obtained at the time of study has not been used in biomarker analyses previously. 29,32,34 –44 All animals were sedated with ketamine (4 mg/kg), xylazine (2 mg/kg), and midazolam (0.2 mg/kg) using intramuscular injection and subsequently anesthetized with 5% isoflurane and 1.5–2.0 L/min of oxygen using a nose cone. Sham animals experienced anesthesia to mirror the protocol for head rotation. Animals in the sagittal-single and sagittal-multiple groups experienced sagittal head rotations with kinematic details below and in Table 2. As previously described, the RNR model produces high-magnitude acceleration/deceleration with the animal securely mounted to a bite plate, producing a purely inertial injury with no impact. 29 The accelerations for each rapid (12–20 ms) sagittal head rotation are similar to those experienced in motor vehicle/high velocity trauma and head impacts in sports. 29,32 Animals in the baseline only group experienced various experimental procedures beyond the scope of the current study; however, only samples collected before any sham/RNR procedures were included for this study.
Number of Animals and Samples by Sex, Group, and Timepoint
Four animals were included in both the sagittal-single and sagittal-multiple (2 R100+) groups because samples collected between the two rotations spaced 24 h apart were included as sagittal-single samples.
F, female; M, male.
Sagittal RNR Angular Kinematics (Mean ± Standard Deviation)
For all other groups, serum samples were collected before sham/injury (pre) and at multiple timepoints postintervention: 30 min, 1 h, 6 h, 1 day, and 1 week. The 0-min timepoint was defined as the time of isoflurane anesthesia administration for sham animals and as the time of first rotation for sagittal-single and multiple groups. Samples collected at approximately 30 min post were defined as <45 min post, and the 1-h timepoint was defined as 45–90 min post. For sagittal-multiple animals with 5 head rotations 8 min apart, the 30 min post sample was collected after the last head rotation. All samples in the 1-day post timepoint were collected 23–30 h post, and samples in the 1-week post were collected at 6–8 days post. Table 1 summarizes the number of animals in each group and the number of subjects sampled at each corresponding timepoint.
RNR head injury model
A RNR head injury model was used to produce mild-to-moderate TBI in piglets following a single rotation and moderate-to-severe TBI following multiple rotations. 32 A single rotation (average peak: 126.8 ± 0.6 rad/s, 50 ± 2 krad/s2) resulted on average in intracranial hemorrhage (ICH) covering 27% of the cerebrum surface area at 3–8 h after injury and 3% at 5–6 days. 37 The sagittal-single injury group corresponds to a single RNR in the sagittal plane; subject group sizes, rotation paradigms, and peak angular velocities and accelerations are summarized in Table 2. The sagittal-multiple group comprised four distinct subgroups with a sequence of rotations: 2 R100+ (comprised two rotations with peak angular velocities >100 rad/s, 24 h apart); 4 R100+ (four 100+ rad/s rotations, 8 min apart); 1 R100+, 4 R60 (sequence of one 100+ rad/s rotation followed by four subsequent rotations with a targeted 60 rad/s peak angular velocity, each spaced 8 min apart); and 4 R60, 1 R100+ (sequence of four 60 rad/s rotations and ending with one 100+ rad/s rotation, each spaced 8 min apart). Rotation rates, or time between rotations, were determined from an adolescent soccer heading exposure dataset previously described. 32
Angular velocity data were captured using single-axis angular rate sensors in duplicate (Diversified Technical Systems, Inc., Seal Beach, CA; or Applied Technology Associates, Albuquerque, NM) attached to the side arm of the RNR device. Angular velocity was measured at 10 kHz and filtered at collection using a CFC 1000 filter. For calculating angular acceleration, angular velocity was filtered with a CFC 180 filter, and a two-point derivation was completed. Peak angular velocity was determined by the maximum absolute velocity corresponding to the first change in acceleration direction (i.e., start of the deceleration phase). Peak kinematics was determined for each sensor independently and averaged for each rotation. Values from the two sensors were averaged for each subject. After filtering, velocity waveforms were typically a half-sine, and acceleration was biphasic, with a prominent deceleration. After filtering, if spurious additional peaks occurred, recordings were excluded from analysis. If a single recording was removed from a rotation, then only data from the other sensor during the same rotation were used.
Serum samples
Whole blood was centrifuged at 8,500 rpm for three minutes (StatSpin VET Express, Iris International Inc., Chatsworth, Los Angeles, CA). Serum was extracted and diluted at 4X; then, the Quanterix Simoa Human Neurology 4-Plex A assay was used to quantify GFAP, Nf-L, tau, and UCH-L1.
A total of 481 serum samples were collected from 215 4-week-old female Yorkshire piglets and 12 serum samples from six uncastrated male piglets (Table 1). As described earlier, subjects were assigned to one of the three experimental groups (sham, sagittal-single, sagittal-multiple) or to the baseline only group, which consisted of subjects undergoing experimental procedures beyond the scope of the current study and only the baseline, or preintervention, serum sample was used. As noted in Table 1, four animals were included in both the sagittal-single and sagittal-multiple (2 R100+) groups. Specifically, serum samples were collected 24 h after a single sagittal RNR before a second sagittal RNR and were used as sagittal-single subjects at 1 day. Then, a second sample was collected 6 h after the second RNR or 30 h after the first RNR, and samples were used as sagittal-multiple subjects at 1 day.
Data processing
For each biomarker, duplicate samples were run, and the results were averaged. When discordance between duplicates was high, samples were unable to be quantified accurately because of a high coefficient of variance >25% (CV = σ/μ × 100). There were 21 total high CV readings (1 GFAP, 3 Nf-L, 5 UCH-L1, and 12 tau), which were removed from further analysis.
Sample values greater than the upper limit of quantification (>ULOQ) could not be accurately quantified within Quanterix’s calibrated range. One sample (sagittal-single, 30 min) was >ULOQ for GFAP and assigned the GFAP ULOQ limit value (3308 pg/mL) for analysis.
For samples less than the limit of detection (<LOD), the concentration could not be quantified accurately in Quanterix’s calibrated range; however, samples were confidently <LOD. Therefore, consistent with established practice, 45 samples determined to be <LOD were given the value of LOD/2 for analysis and for visual displays to ensure inclusion in the data set. Similarly, samples determined to be below the lower limit of quantification (<LLOQ) were given the value of LLOQ/2.
Two subjects’ baseline values were determined to be outliers, calculated as greater than four standard deviations away from group mean. All timepoints of that biomarker for those two subjects were removed from analysis (1 sagittal-single Nf-L, 1 baseline only UCH-L1). Five postinjury datapoints were also removed from analysis owing to being greater than four standard deviations above the group postinjury timepoint mean. All were in the sagittal-single group with one sample removed from each of the following groups: GFAP (30 min and 1 day), Nf-L at 1 day, and UCH-L1(1 day, 1 week). Statistical results run with and without outliers remain unaffected.
After removing the samples described earlier, the number and percentage of samples that were <LLOQ and <LOD are quantified in Table 3 by group and timepoint. As anticipated from a prior study indicating that the human tau antibody is nonreactive to swine and our personal communication with Quanterix, 46 a high percentage of tau samples were <LLOQ (>75% of values in nearly all timepoints). Therefore, total tau was not analyzed further in the current study.
Samples Below the Limit of Detection or Lower Limit of Quantification
GFAP, glial fibrillary acidic protein; LLOQ, lower limit of quantification; LOD, limit of detection; Nf-L, neurofilament light; UCH-L1, ubiquitin C-terminal hydrolase-L1; ULOQ, upper limit of quantification.
Statistical analysis
For female serum samples, the 95% healthy RRs (i.e., 2.5–97.5 percentile concentrations) were calculated from a random subset of female baseline (pre) samples from all groups (n = 149) and validated with the remaining subset of female baseline samples (n = 66) using Fisher’s exact tests (p < 0.05) and a criterion of >90% of validation samples within the healthy RR. Sham and injured animals had measurements taken at additional postintervention timepoints. The four sagittal-multiple paradigms were combined into one group for statistical analysis and comparison to other groups, but subgroup distinctions were noted in graphical displays to inform qualitative interpretation. Within-group serum concentrations were compared at each timepoint to their matched baseline values using Wilcoxon signed-rank test with Bonferroni correction for multiple comparisons (p = 0.05/5 timepoints yielding statistical significance at p < 0.01). As a second analysis, postintervention serum sample concentrations were converted to binomial distributions (inside or outside healthy RR) to evaluate significant group differences at each timepoint from the overall healthy RR binomial distribution using Fisher’s exact tests with Bonferroni correction (p < 0.01). Finally, serum levels were compared between groups (3) within a timepoint using Mann–Whitney U tests with Bonferroni correction (p = 0.05/3 groups yielding statistical significance at p < 0.017).
For male sagittal-single serum samples, male mean 1-day serum concentrations were compared to male pre using Wilcoxon signed-rank test (p < 0.05). Male samples were compared with female sagittal-single pre and 1-day samples using Mann–Whitney U tests (p < 0.05). Male samples at pre and 1 day were compared with the female healthy RR using Fisher’s exact tests (p < 0.05).
Results
Biomarker serum concentration female healthy RR
Values of GFAP (mean ± standard deviation) in the female healthy RR development group were 25.7 ± 15.8 pg/mL with a 95% healthy RR of 6.3–69.4 pg/mL (Fig. 1A). The validation group was not significantly different from the healthy RR with 94% of samples falling within the range (p > 0.999). For Nf-L, mean healthy RR development group levels were 28.0 ± 14.6 pg/mL with a 95% healthy RR of 9.5–67.2 pg/mL, and the validation group did not significantly differ with 97% of samples within the healthy RR (p > 0.727, Fig. 1B). For UCH-L1, mean healthy RR development group levels were 97.2 ± 232.2 pg/mL with a 95% healthy RR of 3.8–533.7 pg/mL, and the validation group did not significantly differ with 98% of samples within the healthy RR (p > 0.999, Fig. 1C). Therefore, healthy RRs were independently validated against separate healthy subjects.
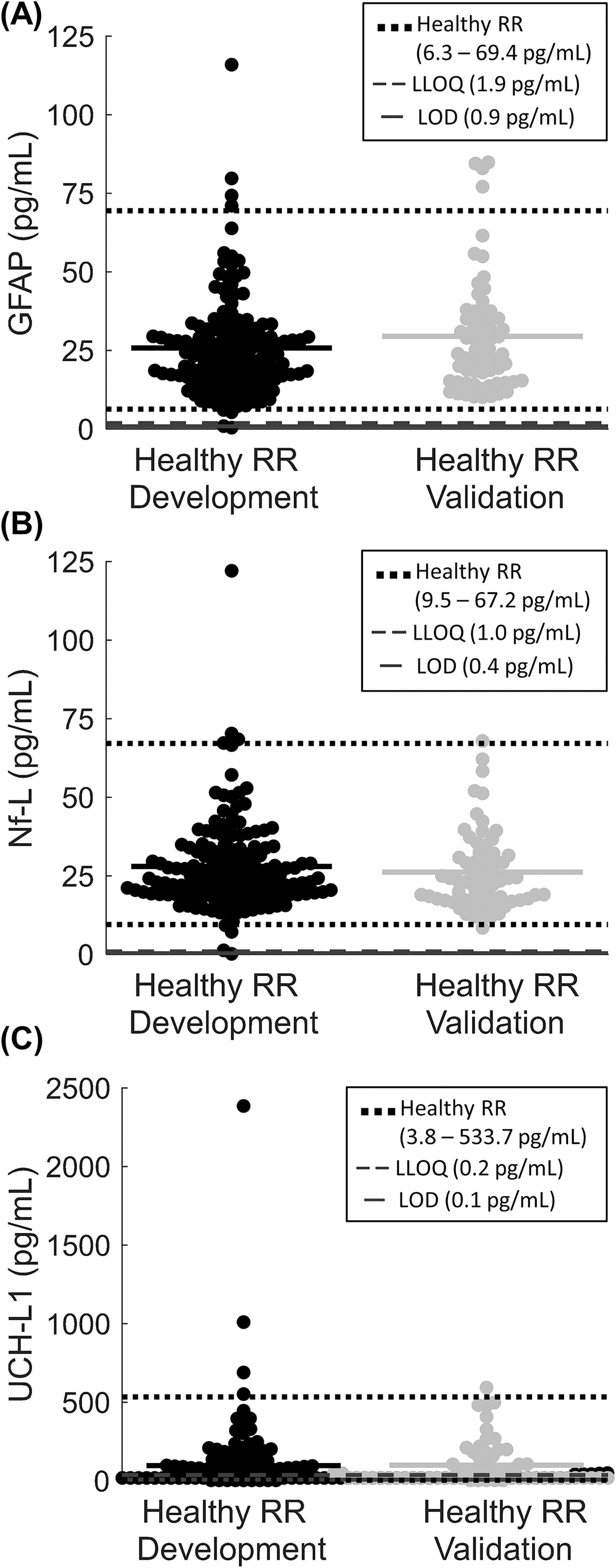
Establishment of serum biomarker healthy reference ranges in female piglets.
Biomarker time-courses after RNR injury
For female animals, mean GFAP levels did not differ for the sham group at any timepoint compared with their pre value (all p > 0.039), and sham levels did not differ from the healthy RR (all p > 0.122), with >80% within the healthy RR at any timepoint (Fig. 2A). In contrast, mean GFAP levels significantly increased above pre levels for sagittal-single animals at 30 min, 1 h, 6 h, and 1 day post (all p < 0.001, refer to Xs below plots) but not at 1 week post (p = 0.429). For sagittal-singles, GFAP levels were outside (above) the healthy RR at 30 min, 1 h, and 6 h post (all p < 0.001, refer to +’s below plots), but not at 1 day or 1 week post (all p > 0.125). In addition, the sagittal-single serum levels were significantly greater than time-matched sham animals at 30 min and 1 h (all p < 0.001, refer to bracket/# in plot) but similar to shams at pre, 1 day, and 1 week post (all p > 0.179). There were no sham samples at 6 h; therefore, no statistical comparisons to sagittal-single were made at 6 h post for any biomarker. Finally, sagittal-multiple GFAP concentrations were significantly higher compared with pre at 30 min post (p < 0.001) but not at 1 h, 1 day, or 1 week post (all p > 0.057). However, sagittal-multiple GFAP samples were outside (above) the healthy RR at 30 min, 1 h, and 1 day post (all p < 0.001) but not at 1 week (p > 0.999). Sagittal-multiple concentrations were also higher than time-matched sham samples at 30 min, 1 h, 1 day, and 1 week post (p < 0.015). Furthermore, sagittal-multiple GFAP levels were significantly higher than sagittal-single at 30 min and 1 day (all p < 0.007) but not 1 h or 1 week (p > 0.032). To summarize, GFAP presented as an early biomarker of injury, with increases for both sagittal-single and sagittal-multiple groups 30 min post compared with pre, healthy RR, and sham animals, and GFAP levels returned back to healthy RRs by 1-week postinjury. Importantly, GFAP levels at 30 min and 1 day post corresponded with load exposure, because sagittal-multiple resulted in values above the healthy RRs that were also significantly higher serum concentrations than sagittal-single.
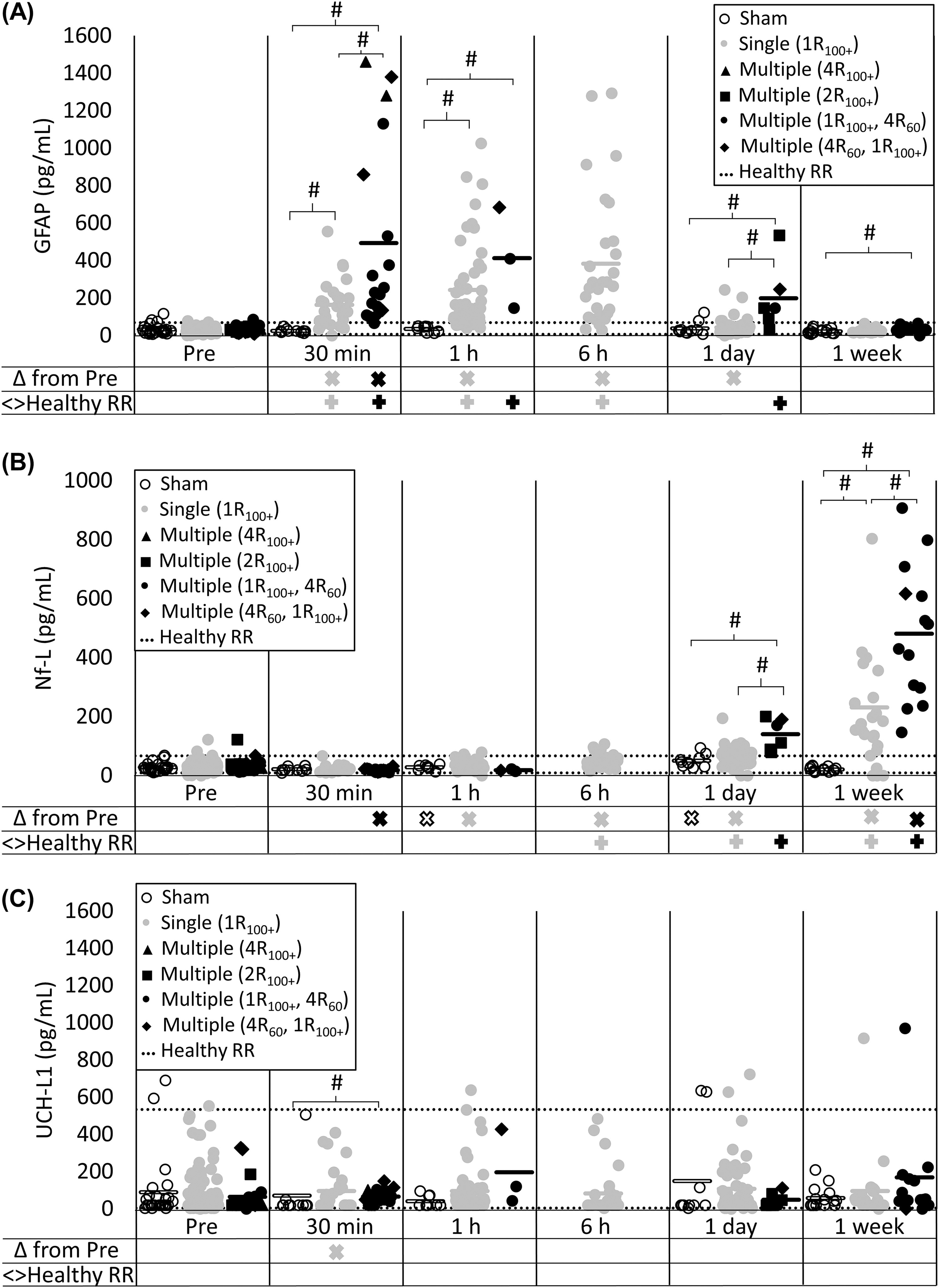
Serum biomarker time-course following rotational traumatic brain injury in piglets.
Mean Nf-L levels for the sham group were different at 1 h and 1 day post compared with pre (all p < 0.008) but not 30 min or 1 week (all p > 0.151); sham concentrations did not differ from the healthy RR at any timepoint (all p > 0.093), with >80% of sham concentrations within the healthy RR (Fig. 2B). For the sagittal-single group, Nf-L significantly increased at 1 h, 6 h, 1 day, and 1 week postinjury compared with their pre (all p < 0.001) but not at 30 min (p = 0.802). Sagittal-single samples were outside the healthy RR at 6 h, 1 day, and 1 week post (all p < 0.005) but not at 30 min or 1 h (p > 0.610). Similarly, sagittal-single levels were greater than time-matched shams at 1 week post (p < 0.001) but not at 30 min, 1 h, or 1 day (all p > 0.044). Mean sagittal-multiple Nf-L levels significantly decreased at 30 min post (p = 0.001), but 100% of samples were within the healthy RR (p > 0.999). Mean sagittal-multiple Nf-L levels significantly increased at 1 week (p < 0.001) but not at 1 h or 1 day (all p > 0.031). Sagittal-multiple samples were also significantly outside the healthy RR at 1 day and 1 week post (all p < 0.007) but not 30 min or 1 h (p > 0.999). Sagittal-multiple levels were significantly higher than shams at 1 day and 1 week post (all p < 0.001) but not at 30 min or 1 h (all p > 0.133). Importantly, sagittal-multiple levels were significantly higher than time-matched sagittal-single levels at 1 day and 1 week (all p < 0.001) but not at 30 min or 1 h (p > 0.033). Therefore, Nf-L presented as a delayed biomarker with sagittal-single and sagittal-multiple groups increasing compared with the healthy RR and sham animals by 1 day and at 1 week. Of consequence, serum Nf-L levels at 1 day and 1 week post corresponded with load exposure, with larger increases above the healthy RR in sagittal-multiple groups compared with sagittal-single.
Mean UCH-L1 levels for sham animals did not differ from pre at any timepoint (p > 0.219), and shams did not differ from the healthy RR at any timepoint (all p > 0.034), with >80% of sham concentrations within the healthy RR at all timepoints (Fig. 2C). Mean sagittal-single UCH-L1 levels were significantly lower than pre at 30 min (p < 0.001) but no other timepoints (all p > 0.013), and sagittal-single did not differ from the healthy RR at any timepoint (all p > 0.451). In addition, sagittal-single levels did not differ from shams at any timepoint (all p > 0.135). Mean sagittal-multiple UCH-L1 levels did not differ from pre (all p > 0.083) or the healthy RR (all p > 0.284) at any timepoint. Sagittal-multiple levels differed from sham at 30 min (p = 0.008) but no other timepoint (all p > 0.038). Sagittal-multiple did not differ from sagittal-single at any timepoint (p > 0.053). Therefore, UCH-L1 was consistent but insensitive to head rotational injury in piglets.
Ten animals (n = 10) were sacrificed before the scheduled end-point owing to poor clinical outcomes (e.g., did not meet standards to return to husbandry, seizure activity, nonambulatory; Table 4). Poor clinical outcome frequency is summarized in Table 5. Although three of the 10 (30%) early sacrifices were in the sagittal-single group, it was a relatively rare event (3% of sagittal-single animals). When focusing on the higher mortality sagittal-multiple groups (Fig. 3), GFAP levels at 30 min post were significantly higher for animals sacrificed early compared with animals that reached scheduled survival timepoints (p = 0.005); GFAP levels were similar at baseline and 1 day post (p > 0.117). Nf-L levels at 1 day trended toward higher for animals sacrificed early, but there were no significant differences at any timepoint (p > 0.058). There were no significant differences for UCH-L1 at any timepoint (p > 0.129). The 1 h timepoint was excluded from analysis owing to a small number of sagittal-multiple animals with available samples (n = 3 of 25). We concluded that GFAP should be investigated further as a potential early prognostic indicator.
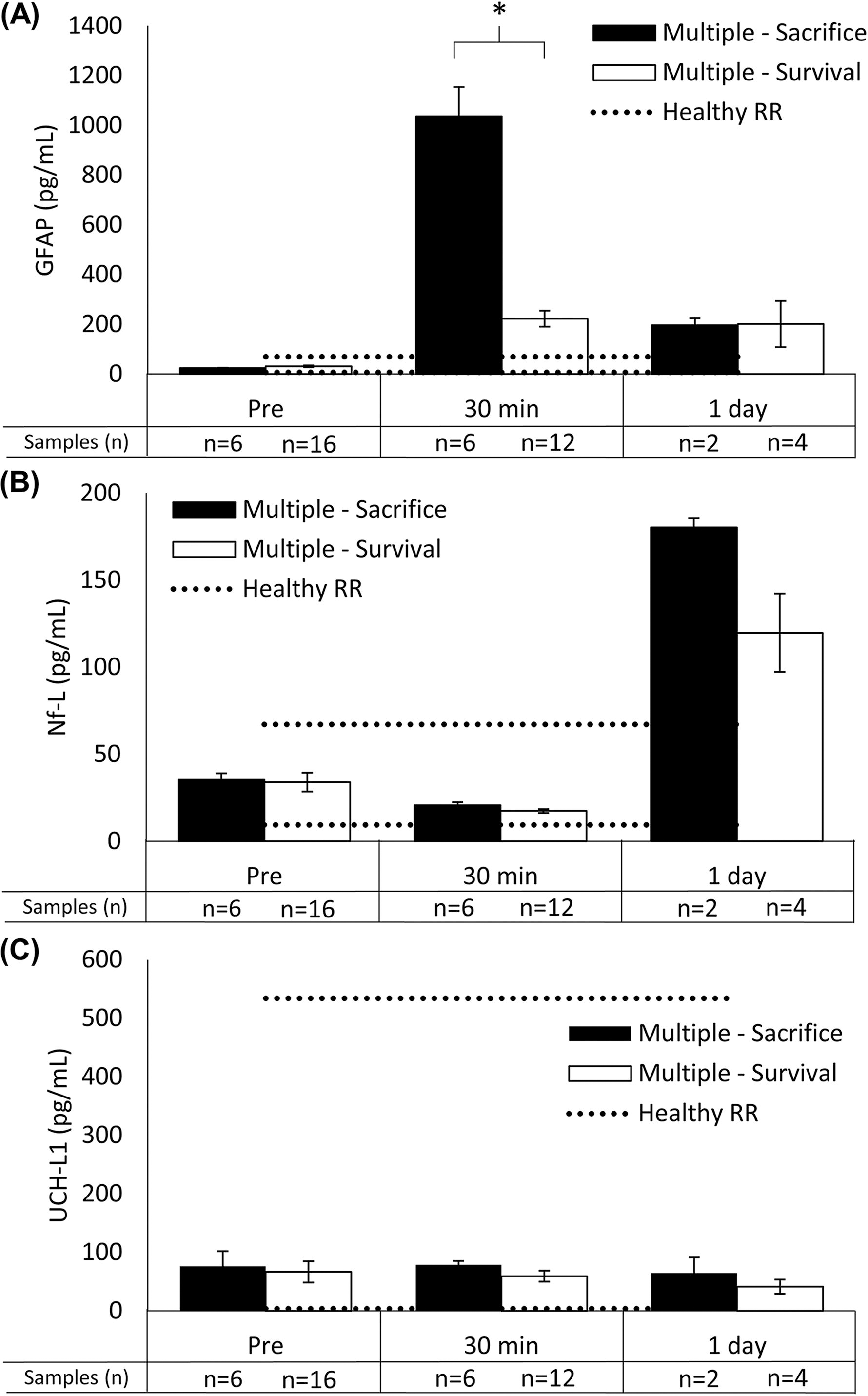
Animals with poor clinical outcomes were predictable using higher GFAP levels at 30 min postinjury.
Details and Medical Reasons for Early Sacrifice
Early Sacrifice Observation Frequency
Sex comparisons in sagittal-single RNR
Serum biomarker concentrations were measured for six additional piglets, uncastrated males who received single sagittal loads (Table 2). Male sagittal-single serum samples were measured at pre and 1 day postinjury and compared with the female sagittal-single data presented previously (Fig. 4). At baseline, males did not differ from female mean baseline GFAP, Nf-L, and UCH-L1 levels (all p > 0.079) or from the female healthy RRs (all p > 0.308), with >83% of values falling within the healthy RR.
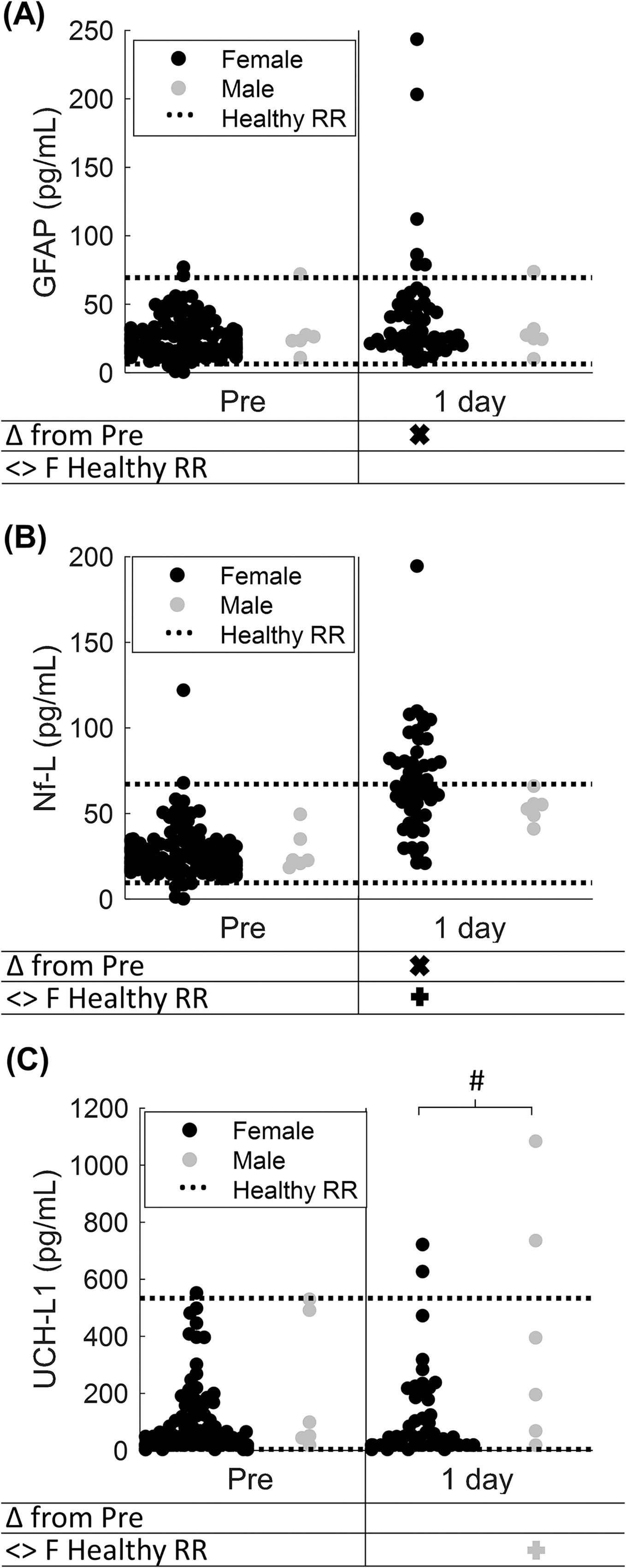
Sex comparisons of serum biomarker levels at baseline and 1 day after sagittal-single RNR.
One day after sagittal-single loads, male mean GFAP levels did not significantly differ from their pre levels (p = 0.563) or from the female healthy RR (p = 0.308, Fig. 4A). Recall that although GFAP in the female sagittal-single group was significantly increased at 1 day compared to pre (p < 0.001), values at 1 day did not differ from the healthy RR (p = 0.125). Finally, male 1-day levels did not significantly differ from female sagittal-single at 1 day (p = 0.706).
Unlike females, which had significant elevations in Nf-L at 1 day, male mean Nf-L levels at 1 day did not significantly differ from pre (p = 0.063) nor the female healthy RR (p > 0.999, Fig. 4B). However, male levels did not significantly differ from females at 1 day (p = 0.109).
For UCH-L1, similar to females, mean male levels at 1 day did not significantly differ from pre (p = 0.438). Unlike females, male samples were significantly outside (above) the female RR at 1 day (p = 0.013), and male levels were significantly greater than females at 1 day (p = 0.026).
In summary, before injury, male serum biomarker levels were similar to female baseline and healthy RR. Although female GFAP and Nf-L levels increased at 1 day post compared with pre and healthy RR for Nf-L, male levels were not altered significantly after injury and remained in the female healthy RR. After sagittal-single, UCH-L1 levels at 1 day were outside the healthy RR for males only and differed from females at 1 day. Importantly, we note that a small sample of males limited our conclusions. We found that sex differences were inconsistent and should be the topic of further study.
Discussion
TBI is common in sports and automobile accidents (>1.5 million in United States per year) causing neurophysiological deficits diagnosed through clinical interpretation of symptoms and brain imaging of hemorrhagic events. 5 Children aged 0–4 years are particularly vulnerable with elevated brain injury rates; 3 however, young children cannot express their symptoms as clearly as adults or older children. Many TBIs need to be treated immediately to relieve ICH and swelling, emphasizing the need for early detection/diagnosis to improve clinical outcomes, 47 but there can be delays, large costs, and long-term health risks associated with neuroimaging, which is the current gold standard for diagnosing ICH. Therefore, rapid, inexpensive, minimally invasive biomarkers such as fluid biomarkers can aid in avoiding unnecessary CT scans, TBI diagnosis, and treatment. Although neuroimaging was not completed in the current study, serum GFAP showed promise as an early injury biomarker in our experimental swine model, as GFAP levels increased substantially above healthy RRs within 30 min following a closed head nonimpact injury produced by a rapid sagittal head rotation. Second, serum Nf-L showed promise as a delayed biomarker with small increases beginning at 1 day post and substantial increases at 1 week. Third, both GFAP and Nf-L were sensitive to load “density,” as animals experiencing multiple head rotations had higher biomarker concentrations rapidly postinjury compared with a single RNR.
Healthy RRs
Our RRs, developed from over 200 subjects, are the first report of a porcine 95% healthy RR. The 95% healthy RRs were calculated for GFAP (6.3–69.4 pg/mL), Nf-L (9.5–67.2 pg/mL), and UCH-L1 (3.8–533.7 pg/mL) in 4-week-old female Yorkshire piglets. In addition, we found that a small group of uncastrated male 4-week-old Yorkshire piglets did not differ at baseline from females. Most published values (mean ± standard deviation) fell within our healthy RR. Owing to the many different types and availability of assays, we limited our quantitative comparisons to other studies using Quanterix Simoa assays where possible. Specifically, in 4-week-old female Yorkshire piglets, 48 mean serum levels for GFAP (7.46 pg/mL) and Nf-L (3.75 pg/mL) were below or within our healthy RR. Similarly, a prior study reported median serum GFAP (10.1 pg/mL) and Nf-L (10.6 pg/mL) levels for young female Yorkshire pigs (35–45 kg) that were within our healthy RR. 49 Mayer et al. reported healthy mean ± standard deviation levels in plasma for GFAP (7.10 ± 4.26 pg/mL), Nf-L (8.03 ± 3.27 pg/mL), and UCH-L1 (175.00 ± 22 pg/mL) in sexually mature Yucatan male and female swine. 46 All levels were below or within our calculated healthy RR despite plasma levels expected to be more concentrated than serum levels. 50 This comparison highlights the current variability in the TBI biomarkers space in processing (i.e., plasma vs. serum), and there may be age and species differences with higher levels of GFAP and Nf-L in Yorkshire piglets compared with adult Yucatan pigs. Next, we compared our porcine healthy RR to published human values. In adult humans, median (interquartile range [IQR]) serum GFAP concentrations of 60.2 (46.4–80.1) pg/mL in male and female control subjects were within or slightly above our porcine healthy RR. 51 Median (IQR) Nf-L levels of 6.3 (3.6–9.2) pg/mL in adult humans were below our porcine healthy RR, and median (IQR) UCH-L1 levels of 8.3 (5.5–11.7) pg/mL were within our healthy RR but at the extreme lower end with substantially less variability. 51 In a study spanning age groups, plasma Nf-L levels (2.6–11.3 pg/mL) were similar in children and young to middle-aged adults; however, GFAP levels (47–226.3 pg/mL) were higher in children aged 3–10 compared with ages 10–60 (21.2–91.9 pg/mL). 14 In a study spanning a broad pediatric age range, serum GFAP levels were higher for children aged 1–3.5 years (80–601 pg/mL) than 11- to 18-year-olds (26–119 pg/mL) and Nf-L levels were higher for children aged 1–9 years (3–21 pg/mL) than 10- to 18-year-olds (2–7 pg/mL). 52 In summary, prior studies in human and swine reported mean baseline GFAP and Nf-L within or below our healthy RR; for UCH-L1, our healthy RR agreed with previous adult porcine studies and were higher and more variable than those observed in adult humans. Baseline serum biomarker levels were validated in an independent group of controls. Moreover, serum biomarker values were stable over time when sampled in sham subjects on multiple occasions. Owing to age and species-specific effects of baseline levels, our study contributes important data for pediatric Yorkshire piglets, and there should be a concerted effort to establish similar healthy RRs across age and species for research and clinical applications. Injured animals demonstrated significant departures in biomarker values from healthy RRs, demonstrating potential utility of healthy RRs as a screening tool for follow-on diagnostics.
GFAP as an early biomarker for TBI diagnosis
GFAP is a protein found exclusively in astroglia in the brain, and therefore, increased GFAP levels indicate glial injury in the brain. 18 However, GFAP also correlates with biomarkers of blood–brain barrier breakdown, and hemorrhagic shock can further exacerbate changes in GFAP. 46 For nonimpact head rotations in the current study, serum GFAP showed promise as an early biomarker of TBI as GFAP increased significantly within 30 min postinjury for sagittal-single (mean 26 → 164 pg/mL) and sagittal-multiple groups (mean 29 → 494 pg/mL) compared with preinjury, sham, and healthy RRs. Furthermore, at 30 min postinjury, mean GFAP levels within the sagittal-multiple group were substantially higher for animals that were sacrificed early owing to poor clinical outcomes (mean 1036 pg/mL) compared with those that reached full survival (mean 222 pg/mL). In addition, GFAP levels began returning to baseline levels by 1-day postinjury and were within the healthy RR by 1-week postinjury. These results showing early GFAP increases were similarly documented in other porcine TBI models. In a model of rotational TBI + hemorrhagic shock (blood loss) in adult pigs, Mayer et al. reported a significant increase in mean plasma GFAP within 30 min of injury (6.7 ± 7.4 → 69.1 ± 63.1 pg/mL). 53 Furthermore, plasma GFAP levels continued increasing through 5 h postinjury (147.7 ± 135.6 pg/mL), which was the study end-point (for Mayer et al. 2021). A similar trend of serum GFAP increases beginning at 1 h postinjury and continuing through 6 h was observed following central fluid percussion injury (cFPI) in adult male Yucatan micro pigs 54 and in the current study. In a focal brain injury model (controlled cortical impact, CCI) in 4-week-old female piglets, median GFAP levels increased by 1 h postinjury (7.5 → 16.3 pg/mL) but continued increasing through 48 h postinjury (89.5 pg/mL). 48 Similarly following CCI in female 4-week-old piglets, plasma GFAP levels increased significantly by 8 h postinjury, peaked at 1 day postinjury, and returned to baseline by 10 days postinjury. 55 Therefore, GFAP translates as an early biomarker across multiple porcine TBI pre-clinical models; however, the loading conditions are critical to magnitude of changes and the time-course of circulating GFAP. Similarly, early GFAP increases were also seen in humans. In human adult patients being treated at a hospital for mild TBI, GFAP began increasing immediately after injury (within 3 h) compared with baseline (∼260 pg/mL), peaked at 20 h (∼1200 pg/mL), and returned to baseline by 72 h postinjury. 23 In sports concussion in collegiate athletes, GFAP levels increased (mean 50 → 90 pg/mL) at the acute timepoint (mean 3.4 h) compared with baseline, uninjured controls, and the current study’s healthy RR. 56 GFAP remained elevated at 24 h postinjury (∼75 pg/mL) before returning to baseline at asymptomatic and 7 day postinjury timepoints. 56 Serum GFAP and UCH-L1 collected within 12 h of suspected head injury have been FDA approved to rule out the need for CT neuroimaging in human subjects with a TBI, such that mean serum GFAP levels from blood drawn within 3 h of injury were substantially higher in CT positive (172 pg/mL) patients compared with CT negative (45 pg/mL) patients. 57 In summary, the current study supports GFAP as an early diagnostic measure with clear increases in the first few hours after injury. Because higher GFAP levels corresponding to animals experiencing multiple head rotations and highest levels were reported amongst animals with poor clinical outcomes, the current study also supports follow-on investigations to explore GFAP as a prognostic measure.
Nf-L as a late biomarker for TBI
Nf-L is a cytoskeletal protein predominately found in myelinated axons serving as a marker of axonal injury. 19 In the current study, serum Nf-L showed promise as a delayed biomarker of TBI as Nf-L increased significantly 1-day postinjury for sagittal-single (mean 27 → 69 pg/mL) and sagittal-multiple groups (mean 34 → 140 pg/mL) compared with sham and healthy RR, and continued increasing through 1 week post for sagittal-single (mean 231 pg/mL) and sagittal-multiple (mean 481 pg/mL). Our results in Nf-L were supported by other porcine TBI models. In a CCI model (in female 4-week-old piglets, median serum GFAP levels increased at 1 day post (3.8 → 14.5 pg/mL) and continued increasing through the study end-point at 5 days postinjury (43.9 pg/mL). 48 Similarly in a CCI model in 4-week-old piglets, Nf-L levels increased significantly at 1 day, peaked at 5 days postinjury, and returned to baseline by 30 days postinjury. 55 Following rotational TBI + hemorrhagic shock in adult pigs, plasma Nf-L increased at 6 h postinjury (6.2 ± 1.6 → 49.5 ± 11.9 pg/mL), 53 which was similar to the start of Nf-L increases in the current study. In a study of Swedish ice hockey players, median (IQR) serum Nf-L levels at 6 days postinjury (12 [10–17] pg/mL) were higher than preseason measurements (9 [6–13] pg/mL), and higher Nf-L levels predicted players to more likely return to play >10 days. 58 In a cohort of collegiate athletes, concussed athletes did not show differences from athlete and nonathlete controls at any timepoint; however, subjects suffering loss of consciousness had increased Nf-L levels at the asymptomatic and 7 days after return to play timepoints. 56 Therefore, Nf-L may serve as a delayed biomarker to inform diagnosis common for secondary concussion clinics that may not see patients until 3–14 days postinjury. To summarize, Nf-L could be explored further as a prognostic measure given increased Nf-L levels with multiple head rotations (compared with sagittal-single) and corresponded to longer return to play periods.
UCH-L1 levels unaffected by pure rotational head injury
UCH-L1 is an abundant, ubiquitinating enzyme with various roles in protein degradation and maintenance expressed predominately in neurons and providing a marker of neuronal injury. 20 In the current study, UCH-L1 did not differ after injury compared with pre, sham, or the healthy RR. Similar results were reported in other porcine TBI models. Following a rotational head injury + hemorrhagic shock in adult pigs, mean plasma UCH-L1 levels did not differ from baseline at 30 min (169.7 ± 169.1 → 390.5 ± 550.0 pg/mL) nor 6 h (103.3 ± 82.7 pg/mL), despite our previous reports of ICH in the RNR model at 3–8 h after injury and at 5–6 days. 37 Similarly following cFPI in adult male Yucatan micropigs, serum UCH-L1 levels were unchanged from baseline at 1 min, 30 min, 3 h, and 6 h postinjury. 54 In general, baseline UCH-L1 levels had higher variability and ranges than previously reported in human subjects. In humans, results were mixed. At approximately 4 h postinjury, serum UCH-L1 did not differ between concussed adolescent patients (190 ± 200 pg/mL) and orthopedic injury controls (260 ± 30 pg/mL). 59 In pediatric patients <12 years old, serum UCH-L1 levels collected acutely (median 3.9 h with a range of 0.5–43.7 h) were significantly higher for patients experiencing a moderate and severe TBI but not mild TBI, compared with controls. 60 The FDA approved UCH-L1 as an early biomarker to help rule out the need for CT neuroimaging in conjunction with GFAP. 22 In human adult patients being treated at a hospital for mild TBI, UCH-L1 peaked within the first 3 h after injury (∼1000 pg/mL) and stabilized by 48 h after injury <250 pg/mL; however, UCH-L1 was also elevated in trauma patients without mTBI (∼400 pg/mL) within the first 3 h after injury. 23 There were also stark differences within 3 h postinjury between TBI subjects with intracranial lesions on CT (mean ∼2500 pg/mL) and those without lesions (∼800 pg/mL). 23 In summary, UCH-L1 is not consistently specific to inform all TBI diagnosis alone, but it may have specific applications at very acute timepoints.
Prepubescent sex differences
At the prepubescent stage in Yorkshire piglets (4-week-old), there were no differences between uncastrated males and females in baseline GFAP, Nf-L, and UCH-L1 levels. Importantly, there were few male subjects, and this analysis is underpowered. We found that there were no sex differences for sagittal-single animals at 1 day post for GFAP and Nf-L; however, response to injury (i.e., GFAP and Nf-L changes from baseline) in females and males differed. After injury, UCH-L1 for males was outside the female healthy RR and differed from female levels at 1 day. Numerous prior pre-clinical and human studies have collected serum pre- and postinjury from males and females; however, results are typically presented together. 51,53,56,60 Therefore, potential sex differences could not be determined, but robust differences between controls and TBI patients were present for multiple biomarkers allowing detection even without sex-specific information. Because the current study had a relatively small male sample size limiting the strength of conclusions, future studies should continue to report results for male and female subjects to determine potential baseline or injury response differences.
Limitations
There are several limitations to the current study. First, a retrospective analysis was completed using cohorts of convenience, from a variety of load conditions and survival end-points, collected over many years from a single laboratory, using consistent methods. This type of analysis did not permit us to study neuroimaging and confirm ICHs in our animal model and establish relationships with our biomarker findings. As a positive, the retrospective analysis allowed the creation of larger sample sizes for greater confidence in establishing healthy RRs and understanding multiple rotation loading conditions. Second, owing to the various protocol end-points and sample timelines, not all serum timepoints were captured for all animals; therefore, statistical analysis focused on changes from baseline levels to capture individual animal variability, as well as the healthy RR of the larger group. A third limitation is that Quanterix (Billerica, MA, USA) developed antibodies for human quantification; however, their assay has been successfully applied to swine previously. Finally, potential species and age differences may limit translation of healthy RRs to humans.
Conclusions
In conclusion, porcine healthy RRs were calculated and provided for 4-week-old Yorkshire female piglets for GFAP (6.3–69.4 pg/mL), Nf-L (9.5–67.2 pg/mL), and UCH-L1 (3.8–533.7 pg/mL); these ranges can be used for reference in pediatric Yorkshire studies, not limited to TBI, such as neurodegeneration, stroke, and cardiac arrest, but also the veterinary science field. GFAP showed utility as an early biomarker (within minutes to hours) of rotational head injury to aid in diagnosis and prognosis with an acute time-course sensitive to the early phases of injury. Nf-L showed increases beginning at 1 day postrotational injury that corresponded with head loading magnitude and shows potential as a delayed biomarker for diagnosis and prognosis at secondary clinics that may see patients 3–14 days after injury. UCH-L1 did not change after mild-to-moderate rotational head injury; however, prior human subject studies have shown that UCH-L1 increases for more severe injuries, especially amongst patients suffering ICHs. Overall, serum biomarkers are a minimally invasive tool that can aid in diagnosis of TBI and other neurological trauma. Further development of minimally invasive measurements could facilitate early diagnosis and intervention decisions.
Transparency, Rigor, and Reproducibility Summary
This study was not formally registered because it was a retrospective analysis using a convenience sample of available serum collected from prior studies in the Margulies Injury Biomechanics Lab. Animal experimental and serum collection methods have been published previously with pre-clinical and physiological data. 29,32,34 –44 The sagittal-single animals (n = 106) were included from multiple prior studies, and 41 received saline vehicle injections for comparison to treated animals. 29 No animals included in the current study received any therapeutic treatments. The analysis plan was not formally preregistered, but the lead author certifies that the analysis plan was prespecified. Sagittal-multiple subgroups were combined before analysis owing to the limited power achieved by small individual injury paradigm subgroup sample sizes. Based on GFAP levels at 30 min postinjury and Nf-L levels at 1 week, an effect size >0.9 was achieved for comparisons between experimental groups.
Footnotes
Acknowledgments
The authors thank the current and past members of the Margulies lab at Emory University, Georgia Institute of Technology, and University of Pennsylvania for their contribution to study design, data collection, collaboration, and support.
Authors’ Contributions
C.M.H.: Conceptualization (equal), data curation (lead), formal analysis (lead), review and editing (equal), investigation (supporting), methodology (equal), project administration (supporting), visualization (lead), writing—original draft (lead), and writing—reviewing and editing (equal). A.D.T.: Conceptualization (equal), investigation (lead), methodology (equal), project administration (lead), visualization (supporting), writing—original draft (supporting), and writing—reviewing and editing (equal). R.A.O.: Conceptualization (equal), methodology (equal), project administration (supporting), supervision (supporting), visualization (supporting), and writing—reviewing and editing (equal). S.S.M.: Conceptualization (equal), funding acquisition (lead), investigation (lead), methodology (equal), project administration (lead), resources (lead), supervision (lead), visualization (supporting), and writing—reviewing and editing (equal).
Author Disclosure Statement
The authors have no conflicts of interest to disclose.
Funding Information
This study was supported by the Georgia Research Alliance and the National Institutes of Health (National Institute of Neurological Disorders and Stroke) awards (R01NS097549, U01NS069545, R01NS039679).