
Abstract
We revisit the hypothesis that there is life in the venusian clouds to propose a life cycle that resolves the conundrum of how life can persist aloft for hundreds of millions to billions of years. Most discussions of an aerial biosphere in the venusian atmosphere temperate layers never address whether the life—small microbial-type particles—is free floating or confined to the liquid environment inside cloud droplets. We argue that life must reside inside liquid droplets such that it will be protected from a fatal net loss of liquid to the atmosphere, an unavoidable problem for any free-floating microbial life forms. However, the droplet habitat poses a lifetime limitation: Droplets inexorably grow (over a few months) to large enough sizes that are forced by gravity to settle downward to hotter, uninhabitable layers of the venusian atmosphere. (Droplet fragmentation—which would reduce particle size—does not occur in venusian atmosphere conditions.) We propose for the first time that the only way life can survive indefinitely is with a life cycle that involves microbial life drying out as liquid droplets evaporate during settling, with the small desiccated “spores” halting at, and partially populating, the venusian atmosphere stagnant lower haze layer (33–48 km altitude). We, thus, call the venusian lower haze layer a “depot” for desiccated microbial life. The spores eventually return to the cloud layer by upward diffusion caused by mixing induced by gravity waves, act as cloud condensation nuclei, and rehydrate for a continued life cycle. We also review the challenges for life in the extremely harsh conditions of the venusian atmosphere, refuting the notion that the “habitable” cloud layer has an analogy in any terrestrial environment.
1. Introduction and Overview
Life on Venus has been a topic of speculation for more than half a century, with published papers ranging from science-fiction-like to invalid conjecture to legitimate hypothesis (Morowitz and Sagan, 1967; Grinspoon, 1997; Cockell, 1999; Schulze-Makuch and Irwin, 2002, 2006; Schulze-Makuch
Almost all previous work on life in venusian clouds does not specify what exactly “life in the clouds” means (for one exception see Schulze-Makuch
We argue that life, if it exists in Venus' atmosphere, must reside inside cloud liquid droplets for the majority of its life cycle (Section 2.1). Life engulfed by cloud droplets will be protected from a fatal net loss of liquid to the atmosphere, an unavoidable problem for any free-floating microbial life-forms. But a droplet habitat implies a lifetime limitation. As liquid droplets coalesce and grow, they eventually reach a size that, due to gravity, settles out of the temperate layers of the atmosphere at an appreciable rate. Over time, the population of inhabited droplets should therefore decline to zero. (Droplet fragmentation—which would reduce particle size—does not occur in venusian atmosphere conditions [Section 3.4].) The conundrum is that there is no way for liquid droplets—and hence life inside of them—to persist indefinitely in Venus' temperate atmosphere layers.
In this article, we describe this problem, and a solution to it. We hypothesize how life escapes being “rained out” down to inhospitably hot atmosphere layers by cycling between small, desiccated spores and larger, metabolically active, droplet-inhabiting cells. Venusian life escapes settling to the surface by forming a resistant, spore-like form that survives the evaporation of the inevitable downward droplet flow to atmosphere layers of high temperature. The desiccated spores become suspended in the Venus atmosphere lower haze layer, which we thus call a “depot” for desiccated microbial life. Because the dynamics of the relatively stagnant lower haze layer are not well known, the main uncertainty in our life cycle hypothesis is how the spores are transported back up into the clouds again. The spores most likely travel upward by vertical mixing induced by gravity waves, and once in the cloud layer they form the nucleus of a new droplet. The depot is “leaky,” that is spores will also vertically mix downward to atmosphere layers with fatally high temperatures. Our proposed life cycle includes cell division that occurs in the larger droplets, and sporulation for individual cells, enabling cell numbers to be replenished against loss.
In this work, we use the terms “microbial life” or “microbes” for microscopic life, without intending to imply that hypothetical venusian microbes might in any way be taxonomically related to microbial life on Earth. We use the term “spore” to denote a cell in a dormant state of long-term metabolic inactivity, which is further resistant to (and protected from) environmental stresses.
We begin with a highlighted review on the very harsh and inhospitable conditions in the venusian atmosphere and related, required assumptions for life to exist (Section 2). We next present our venusian life cycle hypothesis (Section 3), which optimistically assumes that the challenges described in Section 2 can be met. We put the hypothesis in the context with Earth's aerial biosphere and other characteristics of the venusian hypothesized aerial biosphere in Section 4. We conclude with a summary in Section 5.
2. Challenges and Assumptions for Life in the Venusian Clouds
The venusian cloud decks are often described as a potentially habitable environment. The severe and unique environmental challenges, however, are often insufficiently explored. In this section, we review and emphasize the incredibly harsh conditions in the venusian temperate clouds and cloud layer—far more extreme than any on Earth. Major assumptions must be made to envision life existing in such harsh conditions. After arguing why life must be confined to the inside cloud droplets, we review the challenges in order of severity.
2.1. Arguments for why microbial life “outside the droplets” is implausible
The requirement for a liquid environment is one of the general attributes of all life regardless of its biochemical makeup. If life's requirement for a liquid environment is universal, then on Venus the only conceivable stable habitat that meets this criterion is inside of cloud droplets.
Our main argument for why Venus atmosphere life outside liquid droplets is not possible is that free-floating life outside of droplets would rapidly desiccate (by net loss of liquid to the atmosphere) in the very dry atmosphere of Venus. Specifically, a free-floating cell will lose water until its internal water activity is the same as the vapor pressure of the atmosphere around it, which is more than an order of magnitude lower than the driest environments on Earth (Section 2.3). A non-desiccated cell will either have a water activity that is lower than that of the atmosphere around it, in which case water will condense onto it and it will then be in a droplet, or have a water activity that is higher than that of the atmosphere around it, and so will dry out. Only cells in a droplet will be stable. Dried out cells cannot actively divide.* To withstand the dry atmosphere of Venus, microbial life must use the protective environment of the inside of the droplet. In our proposed hypothesis, only one small part of the life cycle has desiccated cells in the form of spores that are dormant.
On Earth, free-floating metabolically active cells outside of cloud water droplets are known to exist, but they are a small fraction of the overall aerial biomass. Free-floating cells are, however in danger of severe desiccation and death, if not deposited back on the surface within a few days. In addition, if such free-floating cells are swept up into the stratosphere, where the stellar UVC becomes the main sterilizing factor, they also die in a matter of days (Bryan
Published papers do not discuss the exact habitat for the hypothesized life in the venusian clouds. One paper does link particles in the lower venusian atmosphere (called Mode 3) to microbial life but only implicitly implies that life resides inside the Mode 3 droplets (Schulze-Makuch
2.2. The high concentrated sulfuric acid environment
The very high concentration of H2SO4 and extreme acidity is a unique challenge for life on Venus. Prior speculations about life on Venus often emphasize that terrestrial polyextremophiles can tolerate very low pH, high temperature, and low water activity environments with a “high concentration” of sulfuric acid. The implication is that the venusian sulfuric acid cloud conditions have similarly low water activity and low pH, and hence are conducive to life (Morowitz and Sagan, 1967; Grinspoon, 1997; Cockell, 1999; Schulze-Makuch and Irwin, 2002, 2006; Schulze-Makuch
This is an incorrect implication. We cannot emphasize enough that the venusian sulfuric acid clouds are much more acidic than even the most harshly acidic conditions found on Earth—the Dallol Geothermal Area, within the Danakil Depression in Northern Afar (Ethiopia) (Cavalazzi
Preliminary studies suggest that even in such harsh polyextreme hydrothermal conditions, life can survive and possibly even thrive; there are examples suggesting that the Dallol acidic pools are inhabited by very small (∼0.2 μm cell size) Nanohaloarchaea and other phyla of archaea (Belilla
The “pH” of venusian clouds defined in a conventional way (−log10[H+]) is meaningless because the conventional pH scale refers only to dilute aqueous solutions. The Hammett acidity value is a measure of acidity that naturally continues the pH scale up to concentrated acids such as sulfuric acid (Liler, 2012). The Hammett Acidity of 85% sulfuric acid is about −11.5 (Yates
There is no Earth-based analogy of life adapting to or living in sulfuric acid concentrations as high as those in venusian cloud droplets. It is quite impossible for terrestrial metabolism to function in concentrated sulfuric acid where the majority of terrestrial biochemicals would be destroyed in seconds. There is extensive literature on the reactions of classes of molecules with concentrated sulfuric acid. Crucial biochemicals are unstable in sulfuric acid and include sugars (including nucleic acids, RNA, and DNA) (Krieble, 1935; Dische, 1949; Long and Paul, 1957), proteins (Reitz
One might suggest that venusian life is insulated from the sulfuric acid environment by elemental sulfur shells (Schulze-Makuch
2.3. Very low water content
If venusian life is water based, then the extremely dry venusian atmosphere is a major challenge for life. Global water vapor mixing ratios in the venusian atmosphere average around 40–200 ppm (Donahue and Hodges Jr, 1992; Barstow
Venusian water activity is much lower than any analogous habitat on Earth. On Earth, some species of filamentous fungi and yeasts are capable of growth at a water activity (
The habitat inside liquid droplets has a higher water content than the atmosphere. The droplets are mostly H2SO4 (85% by volume, on average) with a much smaller component of liquid water (15%). The droplets in the clouds of Venus do have a varying composition from ∼75% H2SO4 at high altitudes to ∼110% (
2.4. Nutrient scarcity
Permanently confined to an aerial biosphere, life must get all its nutrients from the atmosphere. We first review and comment on the availability of carbon, hydrogen, nitrogen, oxygen, phosphorus and sulfur (CHNOPS) biogenic elements before turning to the main nutrient scarcity, metals.
CHNOPS elements are all present in the venusian atmosphere habitable layer as, for example, CO2, N2, SO2, and H2O, in modest amounts. Even phosphorus compounds are measured to be present in the venusian atmosphere. The abundance of phosphorus in the venusian atmosphere and on the surface has only been measured by one of the Venera descent landers (although this X-ray-based detection determined the elemental P abundance it did not determine which P-bearing chemical species the phosphorus was in). In the altitude range of 52 and 47 km, the abundance of phosphorus appears to be on the same order as the abundance of sulfur (Vinogradov
There are examples on Earth of microbes that obtain all their C and N from the atmosphere. The chemolithoautotrophic and acidophilic bacterium
Venus is heavily depleted in hydrogen with respect to Earth. This is illustrated by the increased ratio of deuterium to hydrogen in the venusian atmosphere. The ratio is ∼100 times higher as compared with the rest of the solar system planets (Donahue
Metals are scarce in the venusian atmosphere, and the low metal abundance is likely to be a major growth-limiting factor. Metal ions are required for many biological functions (
Many metals are soluble in H2SO4, including many silicate ores and other metal salts (although glass and silica species are not). There is extensive literature dating many decades back about the study of metal salts in concentrated sulfuric acid, reviewed in part in Liler (2012). It may be worth acknowledging that although Earth-like biochemistry requires metals for biochemistry this may not be a universal requirement for all life. In principle, many biochemical functions of metals can be substituted by other specialized molecules built exclusively from biogenic CHNOPS elements (Hoehler
The scarcity of non-volatile nutrients, including metals, could, in principle, be mitigated by meteoritic delivery. The degree to which meteoritic delivery might be efficient is unknown. Although Venus is volcanically active, the planet lacks the global tectonic activity seen on Earth (
Life on Earth can inhabit environments where nutrients are extremely scarce. A recent example is the discovery of microbial ecologies in the lower oceanic crust (Li
2.5. Energy requirements and the assumption of photosynthesis
Living in the nutrient-depleted incredibly harsh conditions of the venusian cloud decks—much more extreme than any conditions found on Earth—is likely incredibly energy intensive. For example, significant energy would be expended on such cellular processes as: active transport of substances through the cell membrane against the concentration gradient (
We must assume that venusian aerial microbial life is photosynthetic, so that energy capture is not a limiting factor. First, this is because there is little chemical energy potential within the atmosphere, and any redox disequilibria on the surface (the type of disequilibria exploited by terrestrial chemotrophs) are inaccessible to cloud-based life. Second, it is because of the abundance of solar energy on Venus and noting that the conversion of light energy to chemical energy through photosynthesis is the fundamental process in which life on Earth harnesses energy (the other being redox reactions). In fact, it has been speculated that the “unknown UV absorber” in the clouds of Venus, a chemical species of unknown identity, absorbing more than half of all UV that the planet receives could be a manifestation of a wide-spread energy-capture process by an aerial biosphere (Limaye
Photosynthesis need not be oxygenic, or even for carbon fixation. Life on Earth uses several strategies to photosynthetically fix carbon, with oxygenic photosynthesis being only one of the possibilities (reviewed in Seager
There are a number of terrestrial examples for utilization of sunlight for metabolic processes other than fixing carbon. (In principle, any chemical reaction or metabolic activity can be coupled to the light-capturing process.) Aphids, for example, have a layer of carotenoids under the cuticle that forms a sunlight-harvesting system coupled to the synthesis of adenosine triphosphate (ATP) (Valmalette
For a specific example of energy requirement and satisfaction, consider the assumption that venusian life uses a water-based solution for the cell interior. In this case, the life would have to employ special strategies aimed at water capture and retention. The energy required to extract one mole of water from 85% sulfuric acid is ∼25 kJ mol−1 (depending on temperature). The solar energy available (solar flux between 380 and 740 nm) in the temperate cloud region depends on the altitude and varies from 636 J m−2 s−1 at 48 km to 952 J m−2 s−1 at 60 km (as calculated from models; P. Rimmer, private communication 2020). Considering a cell with a radius of 1 μm has a volume of about 4.2 fL, the energy needed to extract the corresponding 2.3 × 10−13 moles of water is about 6 × 10−9 J. The visible-wavelength solar energy available at 52 km is 760 J m−2 s−1, translating to about 9.5 × 10−10 J for a 1 μm cell area. There is, therefore, plenty of light energy incident on the cell surface area. Energy from on the order of seconds to minutes of illumination for a microbe 1 μm in radius would be sufficient for life to extract sufficient water from its sulfuric acid environment to fill the organism.
Ultraviolet radiation should not be a problem for any assumed venusian life because of UV-protecting pigments, such as melanins. It is possible that melanins can harness high-energy electromagnetic radiation (a phenomenon called radiosynthesis) for useful metabolic activity. Some species of fungi (
3. A Proposed Cycle for Venusian Aerial Microbial Life
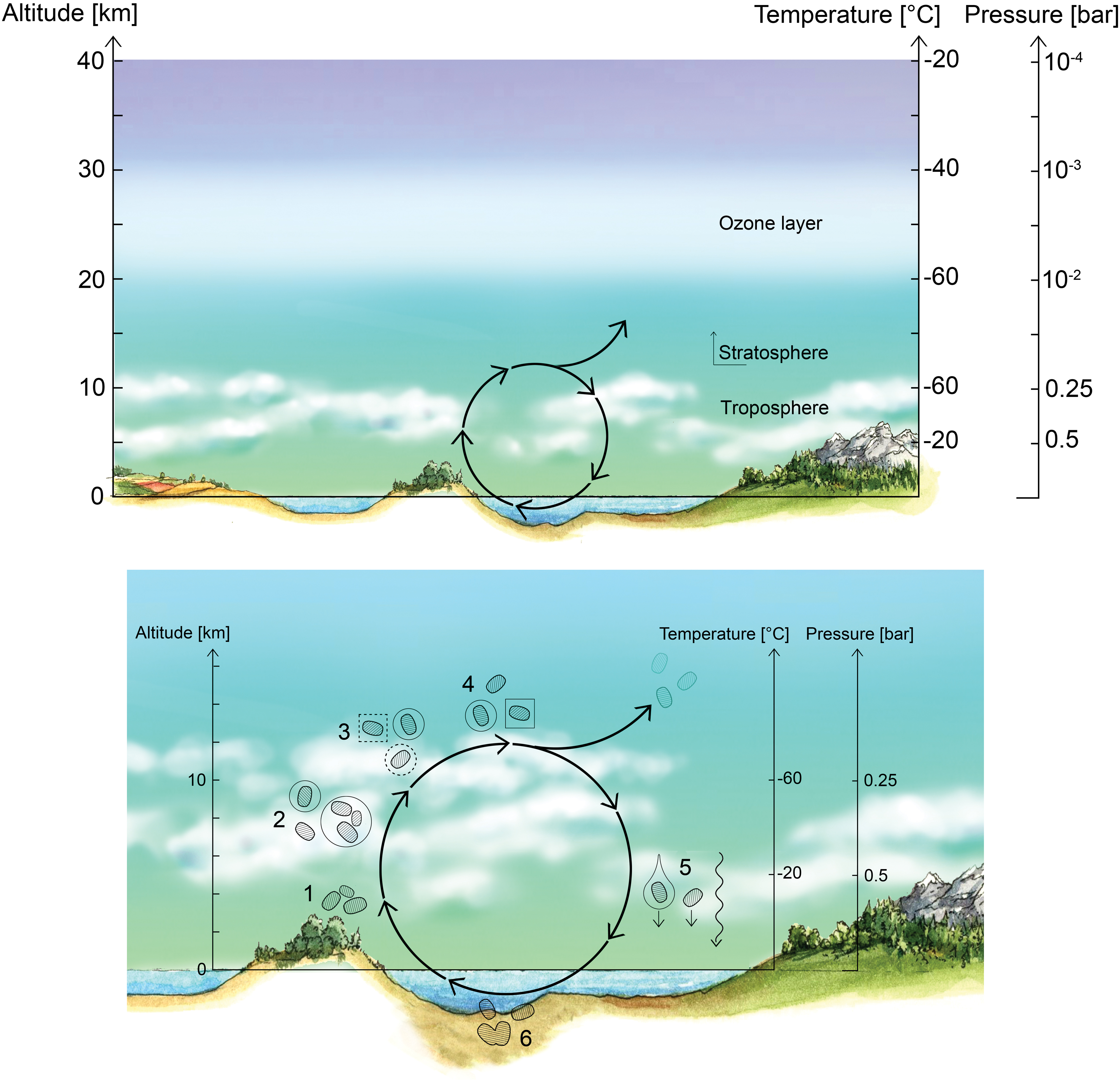
We propose a life cycle for venusian microbes (Fig. 1) that begins in the lower haze layer where desiccated spores reside in a dormant phase. The spores are transported upward by vertical mixing induced by gravity waves to a habitable layer of temperate conditions. Acting as CCN, the spores become encased in a liquid droplet (mostly H2SO4 with some H2O) and germinate. During months aloft, the bacteria metabolize and divide. The cloud droplets, meanwhile, collide and grow to a size large enough that gravity forces them to settle downward. On the downward journey, triggered by changes in the environment (increasing temperatures and the concomitant evaporation of liquids), the bacteria sporulate, preserving themselves as desiccated spores. Once reaching the stable, long-lived stagnant lower haze layer “depot,” the spores remain dormant until the life cycle can begin again. In this section, we describe each step in more detail.
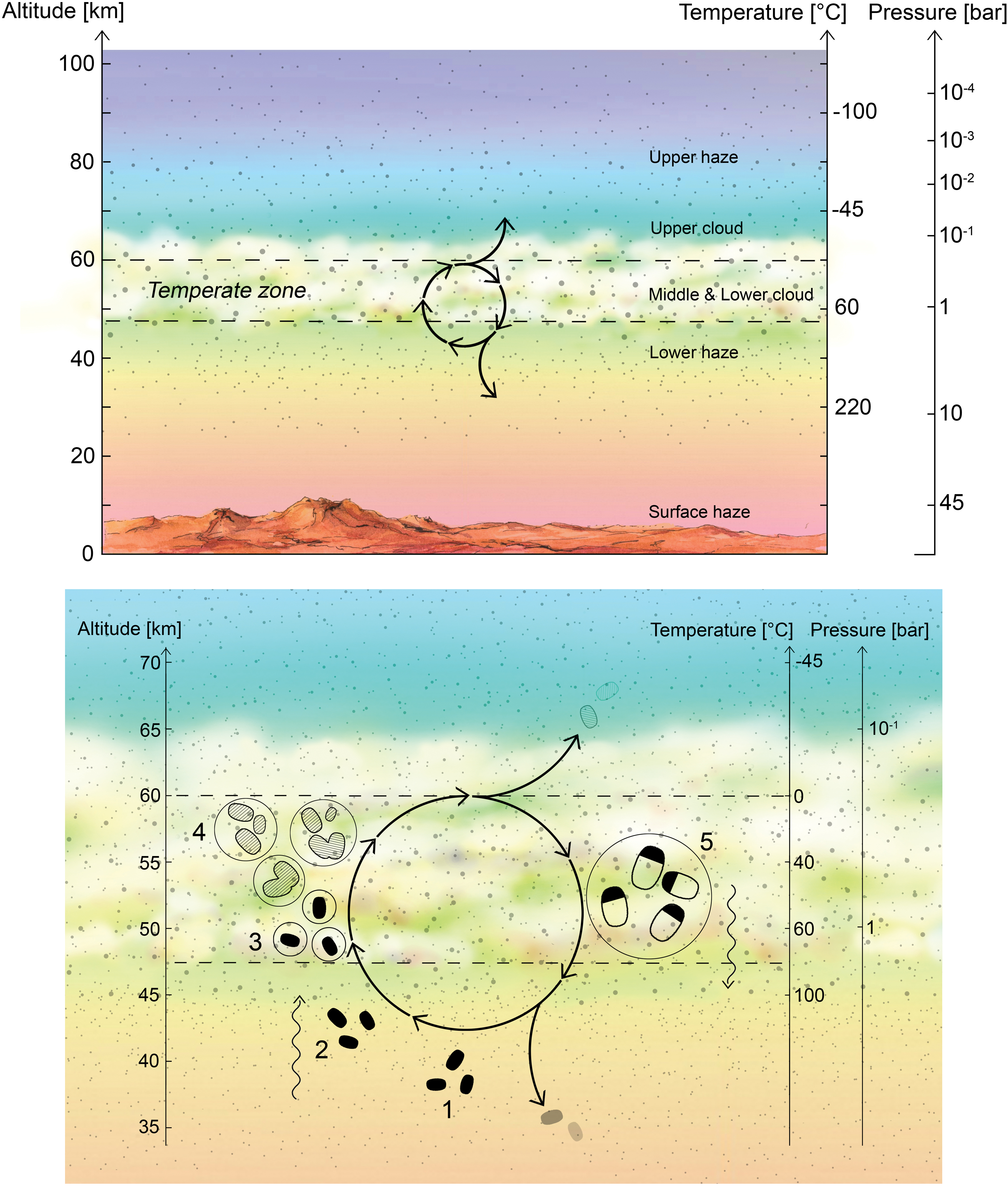
Hypothetical life cycle of the Venusian microorganisms. Top panel: Cloud cover on Venus is permanent and continuous, with the middle and lower cloud layers at temperatures that are suitable for life. Bottom panel: Proposed life cycle. The numbers correspond to steps in the life cycle as described in the main text. (1) Desiccated spores (black blobs) persist in the lower haze. (2) Updraft of spores transports them up to the habitable layer. (3) Spores act as CCN, and once surrounded by liquid (with necessary chemicals dissolved) germinate and become metabolically active. (4) Metabolically active microbes (dashed blobs) grow and divide within liquid droplets (solid circles). The liquid droplets grow by coagulation. (5) The droplets reach a size large enough to gravitationally settle down out of the atmosphere; higher temperatures and droplet evaporation trigger cell division and sporulation. The spores are small enough to withstand further downward sedimentation, remaining suspended in the lower haze layer “depot.” CCN, cloud condensation nuclei. Color images are available online.
3.1. Step 1: Desiccated spores populate the lower Venus atmosphere haze layer, a depot of hibernating microbial life
Venus has a lower haze layer of relatively low mass and unknown composition (Titov
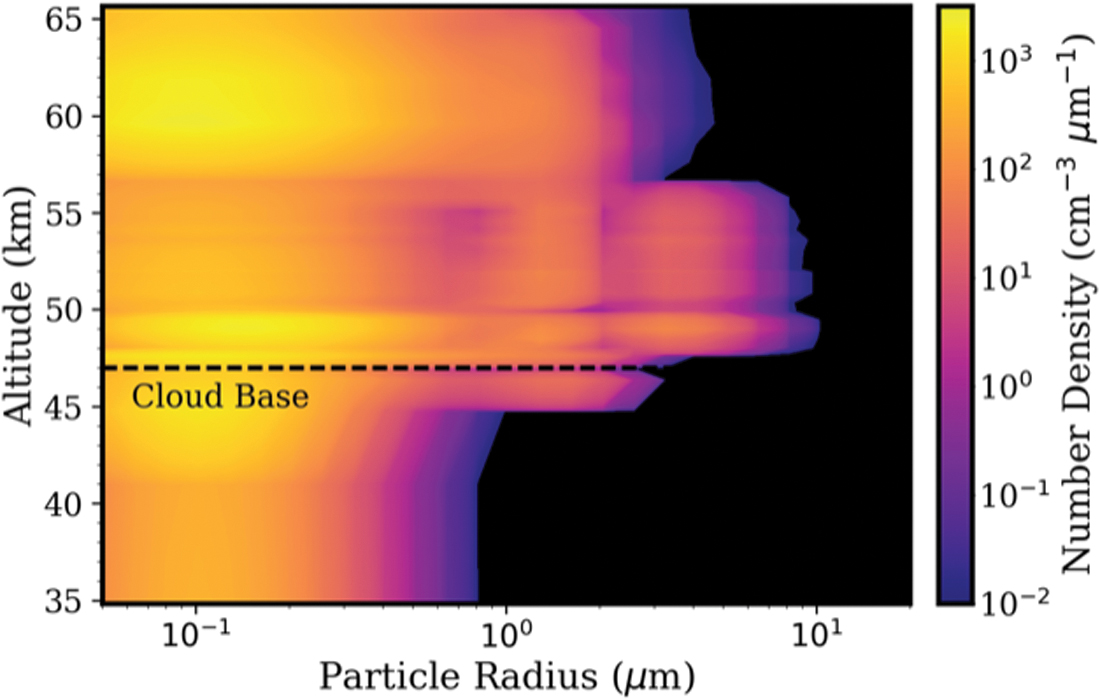
Cloud and haze particle size distribution as a function of altitude. The contour colors show the number density of particles. Particles in the lower haze layer (33–47.5 km) are predominantly smaller than 1 μm (relevant to Step 1 of the proposed life cycle). Particles in the lower and middle clouds (47.5–56.5 km) can extend in radius to well more than 1 μm (relevant to Step 3 of the proposed life cycle). Data from Knollenberg and Hunten (1980), Table 4. Color images are available online.
Desiccated spore sizes must be consistent with the small particle radii in the lower haze layer. This motivates the question of whether spores of that small size are large enough to contain all the required “cell machinery.” Although the question of minimum cell size has only been considered for a hydrated free-living cell, it is likely similar for a spore. Cells must have sufficient space to accommodate metabolic machinery. Based on terrestrial life, a cell is unlikely to be much smaller than 0.2 μm in diameter based on the actual volume of genetic material, enzymatic complexes required for replication, transcription, and translation, in addition to a set of other proteins that contribute to basic physiological processes (Luef
Although these minimal cell sizes are established for Earth's microbial life, it is unlikely that the hypothetical venusian aerial life can be much smaller. Any complex metabolic activities would require complex biochemical machinery, which would require enough cell volume to function properly, even if the chemical basis of venusian life is different than on Earth. Interestingly, the minimal recorded bacterial spore size on Earth is 0.25 μm diameter (Staley, 1999), with a more general range of 0.8 to 1.2 μm diameter (Krieg and Holt, 1984; Ricca and Cutting, 2003). For comparison, mean particle radii in Venus' lower haze layer generally range from 0.2 to 0.5 μm, but with radii as large as 2 μm at the top of the lower haze layer (Knollenberg and Hunten, 1980) (Fig. 2). Therefore, the particle size distribution in the venusian atmosphere lower haze layer is compatible with the known range of cell and spore sizes of Earth's microorganisms.
We refer to the lower haze layer as a depot due to its relative stagnation compared with the rest of the Venus atmosphere, allowing desiccated bacteria to persist for a prolonged period.
For example, the sedimentation timescale,
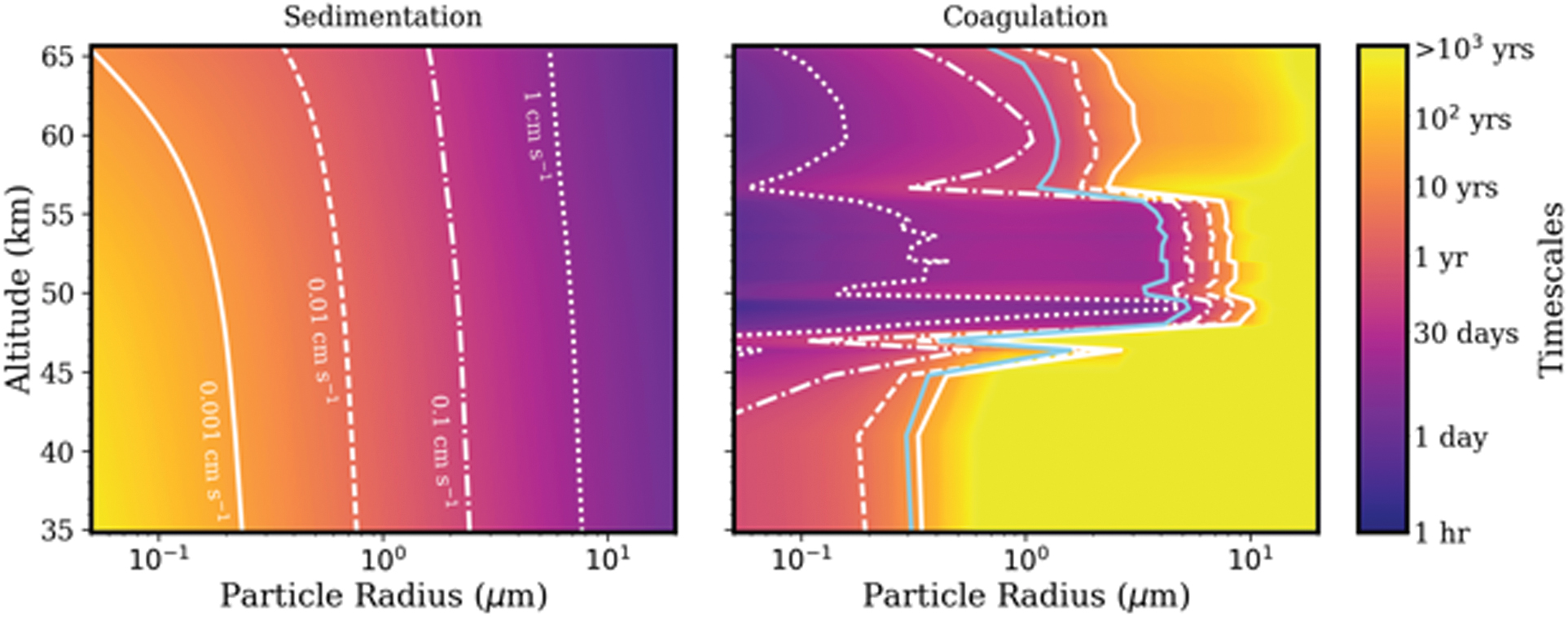
Sedimentation velocities (left) and growth by coagulation timescales (right) as a function of particle radius and altitude. The contour colors show timescales, whereas the white curves show sedimentation velocities on the left, and sedimentation velocities needed to quench coagulation on the right. The blue curve on the right shows where the sedimentation timescale equals the coagulation timescale. Small particles in the lower haze layer (35–47.5 km) are long lived against sedimentation with timescales of tens to thousands of years for particles smaller than 1 μm (relevant to Step 1). Particles in the lower and middle cloud layers (47.5–56.5 km) coagulate on the timescale of hours to months, and particles on order 1 to a few microns in size may be stable against sedimentation for months to years (relevant to Step 4). Sedimentation and coagulation timescales are calculated by using data from Knollenberg and Hunten (1980), table 4, and the Venus International Reference Atmosphere (Seiff
where
Venus has a Hadley cell flow that moves upward at the equator and downward at the poles, but not enough is understood to know what altitude particles trapped in the flow may be lofted to or deposited in. In addition, the location of the returning branch is not known and thus may not impact the lower haze (Sánchez-Lavega
3.2. Step 2: Desiccated spores in the lower Venus atmosphere haze layer travel up to the lower clouds by mixing via gravity waves, followed by convective entrainment
The spores in the lower haze layer must be transported upward to continue their life cycle, but the relative stagnation of the lower haze layer, as described in Section 3.1, creates a challenge. One possible solution is the action of gravity waves, which appear to be present in the lower haze layer due to the layer's static stability. Although gravity waves can only lead to the net transport of energy and momentum and not matter, they can compress atmosphere layers as they travel and contribute to atmospheric mixing.
The gravity waves could be launched by convective plumes arising in the adjacent (50–55 and ∼18–28 km) convective regions (Schubert
We hypothesize that gravity waves mix some of the spores in the lower haze into the lower clouds, where they can participate in cloud formation. We parameterize the mixing by using eddy diffusion, which approximates turbulent transport in planetary atmospheres. Particles, if small enough, can follow the gas in eddy diffusion (Lilly, 1973). The eddy diffusion coefficient (
where
Gravity waves operate in both vertical directions, meaning that some desiccated spores will be lost to lower, hotter layers. Thus, the lower haze layer may be more of a “leaky” depot for desiccated spores.
In the lower haze, where temperatures are too warm for condensational growth, coagulation may play an important role. The coagulation timescale,
where
In summary, although the dynamics of the lower haze layer are highly uncertain, upward (and downward) transport of haze particles is likely accomplished through mixing via gravity waves. Once transported upward to the bottom of the lower cloud layer, particles may continue to efficiently move upward into the clouds by convective entrainment.
3.3. Step 3: The desiccated spores act as CCN and once surrounded by a liquid droplet germinate to a metabolically active life-form
We have argued in Section 2.1 that active microbial life must live inside a droplet, so the spores must be only part of the life cycle. This section describes how the spores become engulfed in cloud droplets.
Once transported to the Venus lower cloud layer, the spores must act as CCN. CCN are “cloud seeds,” a small solid surface needed for vapor to condense. Unlike the relatively high temperature at lower haze altitudes, the lower cloud layer has temperatures where liquid and vapor sulfuric acid and water can coexist. Sulfuric acid (H2SO4) vapor is produced photochemically from SO3 and water vapor, with more H2SO4 being produced at higher altitudes because of the higher flux of UV radiation.
There is a precedent for spores acting as CCN. Aerial bacteria on Earth act as CCN for ice nucleation (Morris
The venusian spores are likely to have a hydrophilic and hygroscopic exterior so they can attract and absorb both sulfuric acid vapor and water vapor. We note that terrestrial bacteria capture water by using hygroscopic biosurfactant polymers. Many microbial polysaccharides and amphipathic lipopeptides, such as syringafactin, from
We envision that the venusian spores only constitute a small fraction of CCN, not large enough to affect prior CCN concepts and calculations. The CCN have previously been suggested to be photochemically generated polysulfur compounds (Toon
3.4. Step 4: The cellular life-form lives in the droplet for months to years, depending on the path of the droplet—during this time, the droplet grows by coagulation
Once in the Venus lower clouds the droplets grow and circulate around the atmosphere. The particles will collide, and each collision for liquid particles results in coagulation, leading to further droplet growth. Coagulation timescales [Eq. (3)] are on the order of days to months in the temperate cloud layers (Fig. 3) and particles may grow to sizes greater than 1 μm. Note that at the same time, zonal flow or Hadley cell motions can carry the droplets around the planet with a timescale of days to months (Schubert
Once the droplet reaches a large enough size (∼1 μm), the cell residing inside the droplet has room to grow and divide. The timescales on which droplets persist in the habitable layer depend on particle size and altitude and are controlled by droplet growth by coagulation and sedimentation due to gravity (Fig. 3). The range can be hours to months to years.
The critical motivating fact for our life cycle description is that droplets will continue to grow by coagulation until the sedimentation timescale becomes shorter than the coagulation timescale, and the particles fall rapidly into deeper, hotter layers of the atmosphere. (Falling much faster than the diffusive transport.) A key question is then: Do the microbes have enough time to metabolize and divide before their droplet home falls to an altitude where they must form spores to survive? The answer is yes. For example, a 3 μm-radius particle in the venusian lower clouds persists for about 6 months, which should be more than enough time for cellular life to germinate from the spore, metabolize, grow, and divide within the same droplet.
Another valid question is whether the droplets in Venus' temperate cloud decks provide enough habitable space. We argue yes. The most numerous droplets in the venusian middle and lower clouds are 2 μm in diameter (1 μm in radius) (Fig. 2). As an example, majority of free-living soil bacteria and archaea have a cell diameter smaller than 0.5 μm, with some as small as 0.2 μm diameter (Hahn, 2004; Portillo
The small size of some species of microorganisms does not mean that they are biochemically simple, or primitive from the evolutionary standpoint. For example, the smallest known free-living photosynthetic organism is the prokaryote
As an interesting aside,
To close out this part of the life cycle, there are two additional peripheral issues. Some particles might updraft up out of the habitable part of the clouds (above 60 km) by being trapped in the Hadley cell flow or by upward diffusion. We note, however, that surviving freezing temperatures in the Venus upper clouds is less challenging than surviving the scorching heat of the lower altitudes. Synthesis of a variety of cryoprotectants is a common strategy employed by life on Earth, including aerial bacteria in Earth's atmosphere, to mitigate extreme cold (Amato
A second peripheral issue is that of droplet fragmentation. If fragmentation could occur, many droplets could avoid growing to sizes with large corresponding sedimentation velocities. More importantly, fragmentation would increase the microbial population in the habitable layer by creating new droplet habitats, some already populated with microbes from prior cell division. By populating the aerial biosphere by fragmentation, the lower haze depot would not be needed.
However, fragmentation of droplets should not occur in Venus' atmosphere. Fragmentation is quantitatively captured by the Weber number,
3.5. Step 5: The bacteria settle down out of the clouds as the droplet reaches a maximum size that can stay aloft against gravity—the decreasing liquid activity triggers cell division and sporulation
Venusian life, trapped living inside of liquid droplets, must adapt to the eventual downward droplet migration to the lower, hotter parts of the venusian atmosphere. Temperatures during the downward fall increase, and this would result in a gradual loss of liquid activity. In other words, the droplets begin to evaporate, and conditions become inhospitable for physiologically active cells. We hypothesize that in response to degrading conditions (high temperature and low liquid activity), microbes begin metabolic preparations for sporulation and deposition of desiccated spore cells. Desiccated to very small sizes, the spores remain in the lower haze, dormant until upward eddy diffusion caused by mixing induced by gravity waves brings them back to the habitable layer.
Sporulation is the ability of various organisms on Earth to form small, desiccated, and metabolically inactive cells, called endospores, or more generally spores (Setlow, 2006). Sporulation is an adaptation to detrimental environmental conditions that aims at preserving the genetic material of the cell when the surrounding environment is inhospitable or lethal for metabolically active cells. The triggers for sporulation on Earth are stress factors, including dehydration, nutrient limitation, and high cell density (Setlow, 2006; Hutchison
The final step in our propsed venusian life cycle is when the dessicated spores settle out into the lower haze layer. As the spores lose liquid and become less massive, their downward settling times slow further. The individual cells must have a coating that prevents cells clumping together as they dry out, such that each individual spore is a single microbial cell. This critical step is the only way to sustain stable numbers of cells in the cloud decks, as it is the only step in the life cycle where the number of distinct atmospheric particles that contain cells can increase (the number of cells increases during the growth phase in the clouds, but the number of particles containing cells remains the same because the cells all remain in one droplet). For example, fungal, bryophyte, and other spores produced in a dense clump in a sporangium (or equivalent) scatter on release because of the properties of the spore-forming body (Sundberg and Rydin, 1998). Each spore remains dormant until mixing brings it back to the beginning of the life cycle (Section 3.2).
Spores could remain viable in the Venusian atmosphere lower haze layer for long periods, based on analogy with Earth life. On Earth, some bacterial spores can survive in extremely harsh conditions for many thousands of years (Nicholson
4. Discussion
With our hypothesized life cycle articulated, we now turn to a discussion for further context.
4.1. Earth's aerial biosphere
Clouds on Earth harbor a diverse species of microbial life, including bacteria, archaea, eukaryotes, and viruses (Amato
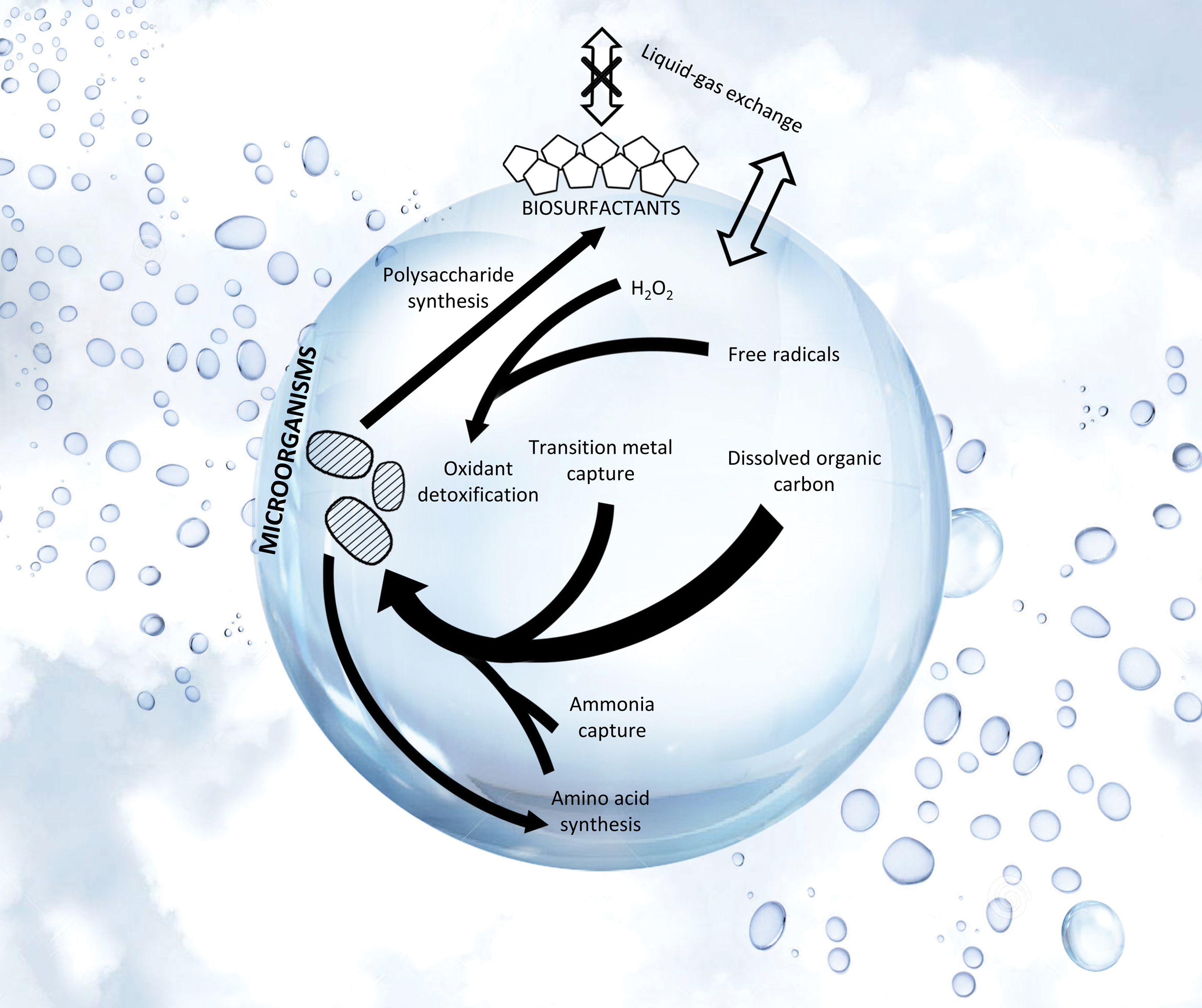
Major metabolic processes for life residing in Earth cloud liquid water droplets. Single-celled microorganisms are shown by the dashed blobs. Selected key adaptations for microbes to survive inside the droplet are depicted by black arrows. Thicker arrows represent more important pathways. The processes were identified from the metatranscriptomics studies of Earth's aerial cloud biosphere (Amato
The transport of microbes from Earth's surface up into the clouds is now known to be a common phenomenon (Vaïtilingom
The life cycle of Earth's aerial biosphere is intimately connected with the habitable surface. Top panel: The cloud cover on Earth is transient and fragmented and is, therefore, not a permanent habitat for Earth's aerial biosphere. (This is in contrast to Venus where the cloud cover is permanent and continuous.) Bottom panel: Life cycle of Earth's aerial biosphere. (1) Updraft of metabolically active microorganisms (dashed blobs) from the surface. (2) Microbial cells are metabolically active both within water cloud droplets (solid circles) and in the free-floating form. (3) Cells likely act as CCN (dashed circle) and promote IN (dashed square) in the atmosphere, promoting droplet formation. (4) Metabolically active cells transiently persist in the atmosphere, are transported over long distances until (5) deposition onto the surface by precipitation or downdraft. (6) On colonization of the new surface habitat, active cell division commences. There is, as of yet, no evidence for cell division in the clouds. Note that Earth's microbial aerial biosphere is metabolically active at every step of the life cycle, and survival is not limited to microorganisms capable of sporulation (Bryan
Microbes are eventually deposited to the surface by precipitation (Vaïtilingom
Earth bacteria swept up from the surface have been postulated to act as CCN of water clouds (Sattler
Earth's clouds are a challenging ecological niche for permanent habitation because of their transient and fragmented nature (in contrast to Venus' permanent and continuous cloud cover). In addition to the fact that Earth's surface is habitable, there is no evolutionary pressure exerted on the microbial biosphere on Earth to adopt a life cycle permanently sustained in the clouds. Rather, the evolutionary selection has likely been focused on temporary survival in the clouds (including cloud-specific complex metabolic functions) in anticipation for the eventual deposition on the habitable surface of the planet. For clouds to be a permanent habitat, active cell division would have to occur in the clouds. Metatranscriptomics studies on Earth's aerial biosphere have not identified transcripts related to active DNA replication and cell division, suggesting that DNA replication and cell division are not performed
Although there is no direct evidence of active cell division available
The characteristics that enable certain microorganisms to retain viability in clouds are likely attributed to the selective advantage of being able to survive long-distance transport to new surface habitats that is provided by temporary cloud droplet colonization (Fig. 5). The habitability of the Earth surface likely squashed any potentially significant evolutionary advantages that might be gained from permanent cloud colonization.
In contrast to Earth, the lack of habitable surface on Venus would force hypothetical microbes to live in the clouds permanently. Therefore, the natural selection pressure on Venus would be directed toward evolutionary strategies that allow life to colonize clouds permanently and not just transiently as it is on Earth. This forms the basis for our life cycle hypothesis: that the entire life cycle of venusian life, including genetic material replication and cell division, must occur in the only temperate region of Venus—the clouds.
4.2. Active mechanisms for remaining aloft?
Throughout the article, we have assumed that the passive mechanisms of air movement drive our proposed life cycle, enabling microbes to remain aloft in the venusian temperate zone for part of their life cycle. Active mechanisms might evolve but are beyond the scope of this article. Active mechanisms could perpetuate life in the clouds of Venus in four ways. First, life could be part of the life cycle proposed here, actively propelling the desiccated spores upward over many kilometers.
Second, life could maintain falling droplets in the cloud layer (
4.3. On biomass and fluxes in the venusian lower haze layer
We are far from being able to construct a plausible ecosystem with biomasses and fluxes, due to lack of information. One estimate, focusing on the abundance of particles (Mode 2 and Mode 3) in the Venus lower and middle clouds, is that the biomass could be 0.1 to 100 mg m−3, which is comparable to Earth's aerial 44 mg m−3 (Limaye
For the context, Earth is estimated to have more than 1030 free-living single-celled microbes (Whitman
For our proposed life cycle to be viable, microbes within droplets must reproduce in large enough numbers (
The venusian atmosphere dynamics, especially in the lower haze layer, are not well enough understood to have any certainty to work out mass or flux balance. All we can say is that upward mixing by gravity waves is a way to move small particulate biomass upward, and would have to approximately equal the planet-wide but slow sedimentation of biomass trapped inside droplets. Vertical transport and fluxes, including Hadley cell transport at the equator and poles, definitely require further study.
5. Summary
Life in the venusian clouds has long since been a popular if speculative topic. The clouds decks of Venus themselves are often described as conductive to life. We reassessed this notion and reviewed the severe and unique environmental challenges that life in the aerial biosphere of Venus would have to overcome. The challenges include: an extremely acidic environment, far more so than any known environment on Earth; very low water content; and nutrient scarcity.
We also highlighted the assumption that life would have to reside inside protective cloud droplets (sulfuric acid mixed with water), and that any life would have to be photosynthetic to have enough energy for a variety of cellular processes.
The main new point of this work is to present a life cycle concept. Assuming that life must reside inside cloud droplets, we resolve the subsequent conundrum of gravitationally settling droplets reaching hotter, uninhabitable regions by proposing a venusian life cycle where a critical step is microbes drying out to become spores on reaching the relatively stagnant lower haze layer, which we call a leaky “depot.” The dried out spores would reside there until some of them can be transported back up to the temperate, habitable cloud layers, where they would act as CCN to promote cloud formation, becoming enveloped in cloud droplets to continue the life cycle.
Footnotes
Acknowledgments
The authors thank Joanna Petkowska-Hankel for the preparation of Figs. 1, ![]() , and 5. They also thank Daniel Koll for useful discussions.
, and 5. They also thank Daniel Koll for useful discussions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank the Change Happens Foundation, the Heising-Simons Foundation, and the MIT Professor Amar G. Bose Research Grant Program for funding. P.G. acknowledges support from the 51 Pegasi b Fellowship funded by the Heising-Simons Foundation.
Abbreviations Used
Associate Editor: Norman Sleep