
Abstract
Hypertensive small vessel disease is a major cause of vascular cognitive impairment (VCI). Spontaneously hypertensive/stroke prone rats (SHR/SP) with unilateral carotid artery occlusion (UCAO) and a Japanese permissive diet (JPD) have white-matter (WM) damage similar to that seen in VCI. We hypothesized that WM injury was due to hypoxia-mediated, blood–brain barrier (BBB) disruption. Twelve-week-old SHR/SP had UCAO/JPD and were studied with immunohistochemistry, biochemistry, multimodal magnetic resonance imaging (MRI), and Morris water maze (MWM) testing. One week after UCAO/JPD, WM showed a significant increase in hypoxia inducible factor-1α (HIF-1α), which increased further by 3 weeks. Prolyl hydroxylase-2 (PHD2) expression decreased at 1 and 3 weeks. Infiltrating T cells and neutrophils appeared around endothelial cells from 1 to 3 weeks after UCAO/JPD, and matrix metalloproteinase-9 (MMP-9) colocalized with inflammatory cells. At 3 weeks, WM immunostained for IgG, indicating BBB leakage. Minocycline (50 mg/kg intraperitoeally) was given every other day from weeks 12 to 20. Multimodal MRI showed that treatment with minocycline significantly reduced lesion size and improved cerebral blood flow. Minocycline improved performance in the MWM and prolonged survival. We propose that BBB disruption occurred secondary to hypoxia, which induced an MMP-9-mediated infiltration of leukocytes. Minocycline significantly reduced WM damage, improved behavior, and prolonged life.
Keywords
INTRODUCTION
Small vessel disease is the major risk factor producing white matter (WM) damage in vascular cognitive impairment (VCI).1–3 Glutamate-mediated excitotoxicity is thought to cause oligodendrocyte death in acute focal ischemia, but less is known about the delayed damage to cells in the WM.4,5 In normotensive rats, bilateral carotid artery occlusion produces inflammatory disruption of the blood-brain barrier (BBB) by matrix metalloproteinases (MMPs) associated with demyelination.6–8 Lowering oxygen induces hypoxia inducible factor-1α (HIF-1α), which leads to expression of a large number of genes. 9 Increased levels of HIF-1α and MMPs are seen pathologically in the WM in VCI patients, suggesting that tissue under hypoxic stress expresses proteolytic enzymes.10,11 In acute stroke, hypoxia initiates a neuroinflammatory response with production of free radicals and proteases, BBB disruption, vasogenic edema, and myelin damage. 12 Hypoxia inducible factor-1α, which is a transcriptional activator and major regulator of the adaptive cellular response to hypoxia, is upregulated after focal ischemia in rat brain. 13 The HIF-1α expression and stability are regulated by HIF prolyl-4-hydroxylases (PHDs). 14 One of the PHD isoforms, PHD2, is expressed in brain tissue and is believed to be the primary regulator of HIF-1α expression. 15 In brain trauma, HIF-1α has a key role in brain edema formation and BBB disruption via a molecular pathway involving MMP-9. 16 A recent study showed that hypoxia-induced MMP-9 expression leads to vascular leakage, which could be reduced by MMP inhibition. 17 Pharmacological blockage of MMP-9 or MMP-9 gene deletion confers neuroprotection in traumatic brain injury and stroke. 18 Thus, there is considerable evidence to suggest that hypoxia-induced MMPs mediate the damage to the BBB that is related to damage to the WM in acute stroke, but little is known about the role of hypoxia in BBB opening and WM injury secondary to chronic hypertension.
Spontaneously hypertensive/stroke prone rats (SHR/SP) show many of the pathologic features of patients with VCI, including fibrosis of small blood vessels, injury to the WM, and behavioral deficits. 19 Normally, SHR/SP can live up to 9 months without magnetic resonance imaging (MRI) or pathologic evidence of brain damage. 20 We have shown that feeding SHR/SP a diet consisting of low protein and high salt, the Japanese Permissive Diet (JDP), along with unilateral carotid artery occlusion (UCAO), produces an accelerated injury to the WM that is associated with MMP expression, BBB disruption, death of mature oligodendrocytes, and expression of oligodendrocyte precursor cells; the extent of injury can be monitored noninvasively with MRI and the behavioral changes with Morris water maze (MWM). 21 Using an oxygen-sensitive crystal with electron paramagnetic resonance, we showed an increase in oxygen extraction in the WM at 12 weeks of life with a drastic decrease in oxygen from the thirteenth to the sixteenth week when the animals died. 22 The present report describes the molecular events leading from hypoxia to WM damage and death, which we hypothesize is a multistep process proceeding from chronic hypoxia to MMP-9-mediated BBB disruption, resulting in myelin loss and behavioral changes. 23 In addition, we treated the SHR/SP/JPD/UCAO with the antiinflammatory agent, minocycline.24,25 Minocycline is a tetracycline derivative that was shown in early studies to reduce infarct size in acute stroke. 26 More recently, it has been studied in a number of neuroinflammatory conditions, and relevant to this study, it was shown to downregulate the expression of inducible nitric oxide synthase in a model of vascular dementia involving bilateral carotid artery occlusion in normotensive rats.27,28
MATERIALS AND METHODS
Animal Groups and Unilateral Common Carotid Artery Occlusion The animal protocol was approved by the University of New Mexico Health Sciences Center Institutional Animal Care and Use Committee (IACUC), and followed the NIH Policy on Humane Care and Use of Laboratory Animals. Male SHR/SP (Charles River Laboratories, Wilmington, MA, USA) were purchased at 6 weeks of age and monitored with blood pressure measurements and weights until 12 weeks of age when the blood pressure has reached a maximum, where it remains unless they are fed the JPD with added salt, which causes the blood pressure to continue to rise. Unilateral carotid artery occlusion was performed in SHR-SPs at 12 weeks of age. The methods for the carotid surgery, specialized diets, and blood-pressure measures have been reported previously and will be described briefly. 21 SHR/SP were divided into four groups (random number sequence generated by: www.randomizer.com): (1) UCAO/JPD; (2) carotid sham-operated without diet; (3) UCAO/JPD DMSO vehicle-treated; and (4) UCAO/JPD minocycline-treated (50 mg/kg) in DMSO. Japanese permissive diet, UCAO surgery, sham surgery, drug or vehicle administration all began on the start of the twelfth week of the animal's life according to which group they had been assigned (end points were determined based on the experimental designs). 21 The vehicle for the minocycline study consisted of 30% DMSO with 25% Solutol (BASF Corp., Florham Park, NJ, USA) in normal saline, which was needed to dissolve drug. At day 1 of UCAO, rats were given the first injections intraperitoneally of drug or vehicle, followed by injections every other day for 9 weeks. All injections were done similarly to avoid bias. The JPD consists of 16% protein, 0.75% potassium, 4% sodium (Ziegler Bros, Inc., Gardeners, PA, USA) with 1% sodium chloride added to drinking water. Our earlier study shows that at 4 weeks after UCAO/JPD (16 weeks of life), neurologic symptoms appear and WM lesions are seen on MRI; they die within 4 weeks of diet and surgery. 21 In the sham-operated group, the right carotid artery was isolated and rats were fed with regular rodent diet with tap water after this procedure. In the present study, rats in the UCAO/JPD and sham-operated groups were killed at 1, 2, and 3 weeks after UCAO/JPD. In the drug treatment study, vehicle-treated and minocycline-treated groups were killed at week 8 or 9 after UCAO/JPD (nineteenth or twentieth week of life) because of the prolonged life with treatment. Females were not used in this study to avoid the effects of hormonal cycling.
Tissue Preparation for Immunohistochemistry and Immunoblotting
Rats were anesthetized with pentobarbital (50 mg/kg, intraperitoneally) and transcardially perfused with 0.1% procaine in cold phosphate buffer followed by 2% PLP (2% paraformaldehyde, 0.1 mol/L sodium periodate, 75 mmol/L lysine in 100 mmol/L sodium phosphate buffer, pH 7.4). Brains were removed and postfixed in 2% PLP for 24 hours at 4°C. For cryo-sectioning, tissues were cryoprotected in 30% sucrose/2% PLP at 4°C, and placed in a Peel-A-Way histology mold (Ted Pella Inc., Redding, CA, USA) containing Tissue-Tek embedding medium (OCT; Sakura Finetek, Torrance, CA, USA). Tissue was frozen in 2-methylbutane cooled in liquid nitrogen. Brains were kept at −80°C until use. Brain tissues were sectioned at 10 μm before histologic analysis. For immunoblotting, rats were perfused with ice-cold saline, brains were removed quickly, snap-frozen in 2-methylbutane chilled to −80°C, then kept at −80 °C until use.
Immunohistochemistry
Brain tissues were prepared for histology as previously described, and tissue was sectioned with a cryostat. 21 Brain sections were stained with antibodies to MMP-9 (1:175, Chemicon, Billerica, MA, USA), glial fibrillary acidic protein (GFAP; 1:400; Sigma, St Louis, MO, USA), CD4 (1:250; Santa Cruz, Dallas, TX, USA), myeloperoxidase (MPO; 1:50; Santa Cruz), HIF-1α (1:200; Abcam, Cambridge, MA, USA), prolyl hydroxylase-2 (PHD2; 1:250; Santa Cruz), RECA 1 (1:300, Abcam), and Cy-3-conjugated Affinity Pure Goat Anti-Rat IgG antibodies (1:500; Jackson, West Grove, PA, USA). Secondary antibodies (1:500), conjugated to FITC (Alexafluor 488; Invitrogen, Grand Island, NY, USA) or Cy-3 (Alexafluor 546; Invitrogen), were applied for 90 minutes at room temperature. 4′-6-Diamidino-2-phenyllinidole (DAPI; Invitrogen) was used to label cell nuclei.
Immunoblotting
Frozen coronal sections were cut at 400 micrometer thickness and micropunched biopsies were collected from both brain hemispheres in WM areas of external capsule (EC), corpus callosum (CC), and internal capsule (IC) at −25°C. 21 Western blotting followed standard techniques, using tissue obtained by microbiopsy punches taken from WM regions. Primary antibodies were against HIF-1α (1:1,000, Abcam), PHD2 (1:5,000, Santa Cruz), and MMP-9 (1:1,000; Abcam). In rat brain tissue lysates, the anti-HIF-1α antibody detected two protein bands of ~ 120 kD (corresponding to post-translationally modified HIF-1α) and ~ 93 kD (predicted molecular weight of native HIF-1α) molecular weight. Since the 93-kD band was constantly detected and appeared as most prominent on all the immunoblots, we considered it as a major protein band and used it to perform quantification analysis for HIF-1α expression.
Magnetic Resonance Imaging
We used a 4.7 -T small animal MRI (Bruker, Biospin, Billerica, CA, USA) to measure WM lesion size with T2-weighted imaging, apparent diffusion coefficients (ADCs), fractional anisotropy (FA), and arterial spin labeling (ASL) weekly after treatment with either minocycline or vehicle. Detailed descriptions of the MRI methods were reported previously. 7
Morris Water Maze
Seven weeks after minocycline and vehicle treatments, cognitive function was assessed as reported previously. 21 We measured latency to reach the hidden platform, and calculated histograms of the probe trial. The cue test was used to show visual impairment. Mean velocities (cm/s) were measured during the acquisition trial.
Statistical Analysis
Data are expressed as means ± s.e.m. Statistical significance was set at P < 0.05. Data were analyzed by two-way ANOVA followed by Bonferroni t-test analysis, and unpaired Student's t-tests, using Prism 5.0 (GraphPad Software Inc., La Jolla, CA, USA).
RESULTS
Western blot analysis showed that at 1 week after UCAO/JPD, HIF-1 a protein level was increased in all examined WM regions in the occluded side (right, R), as compared with the same regions in sham-operated animals. Similar increase in HIF-1α was observed in all regions of the nonoccluded side (left, L), which is consistent with the observation that the UCAO/JPD model presents WM damages at both occluded and nonoccluded sides. 21 The same pattern of HIF-1α increase was observed at 3 weeks after UCAO/JPD, in all examined WM areas (including CC, EC, and IC, both in occluded and in nonoccluded sides (Figure 1A). The HIF-1α colocalized with GFAP immunoreactivity, demonstrating the expression of HIF-1α in the CC astrocytes (Figure 1B).
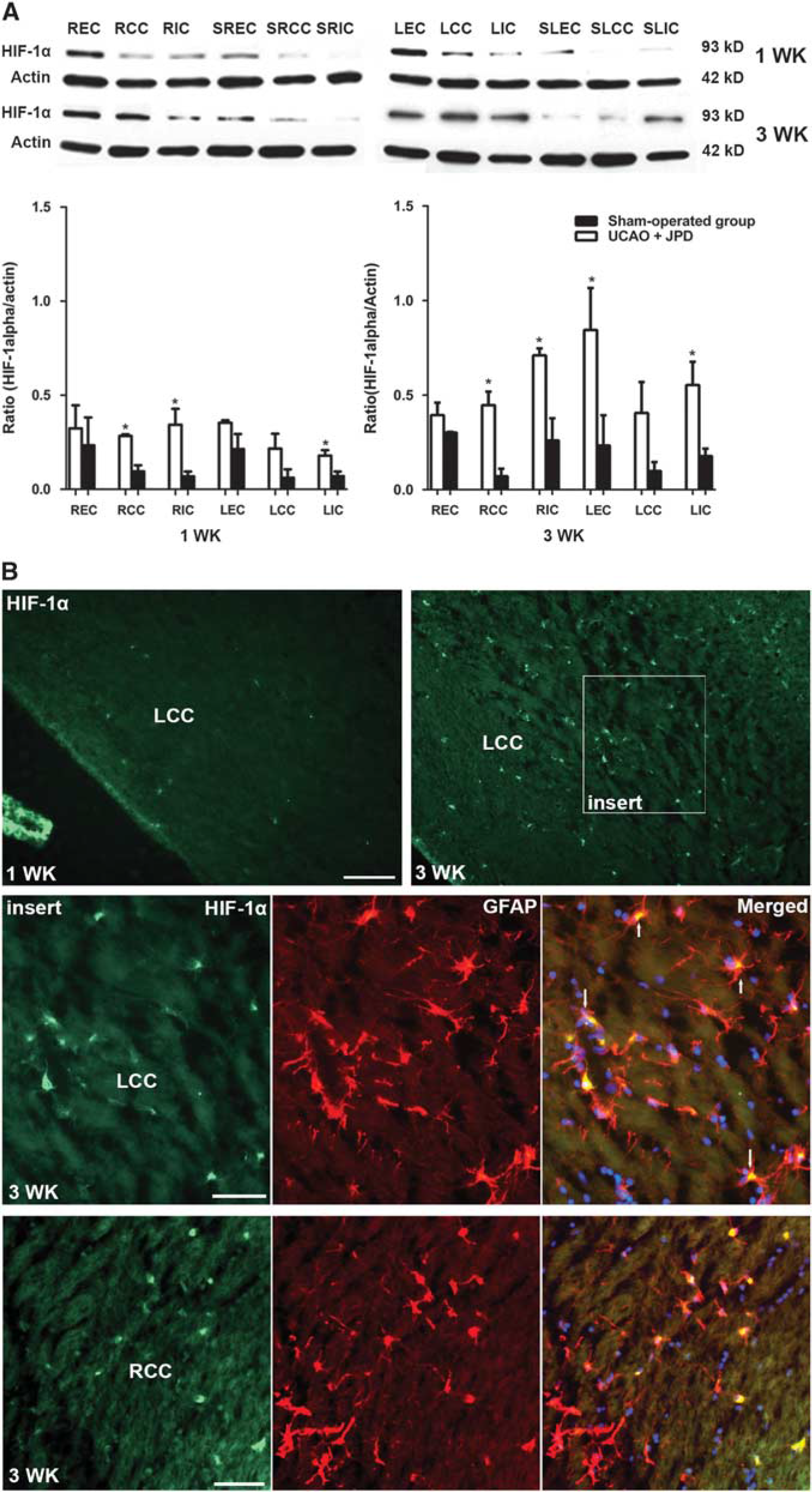
Increased hypoxia inducible factor-1α (HIF-1α) in unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD). (
Western blot showed that PHD2 was decreased in several regions on the occluded side and EC of the nonoccluded side 1 week after UCAO/JPD. While levels of PHD2 increased in the sham-operated animals at 3 weeks, they remained low in the SHR/SP/UCAO/JPD (Figure 2).
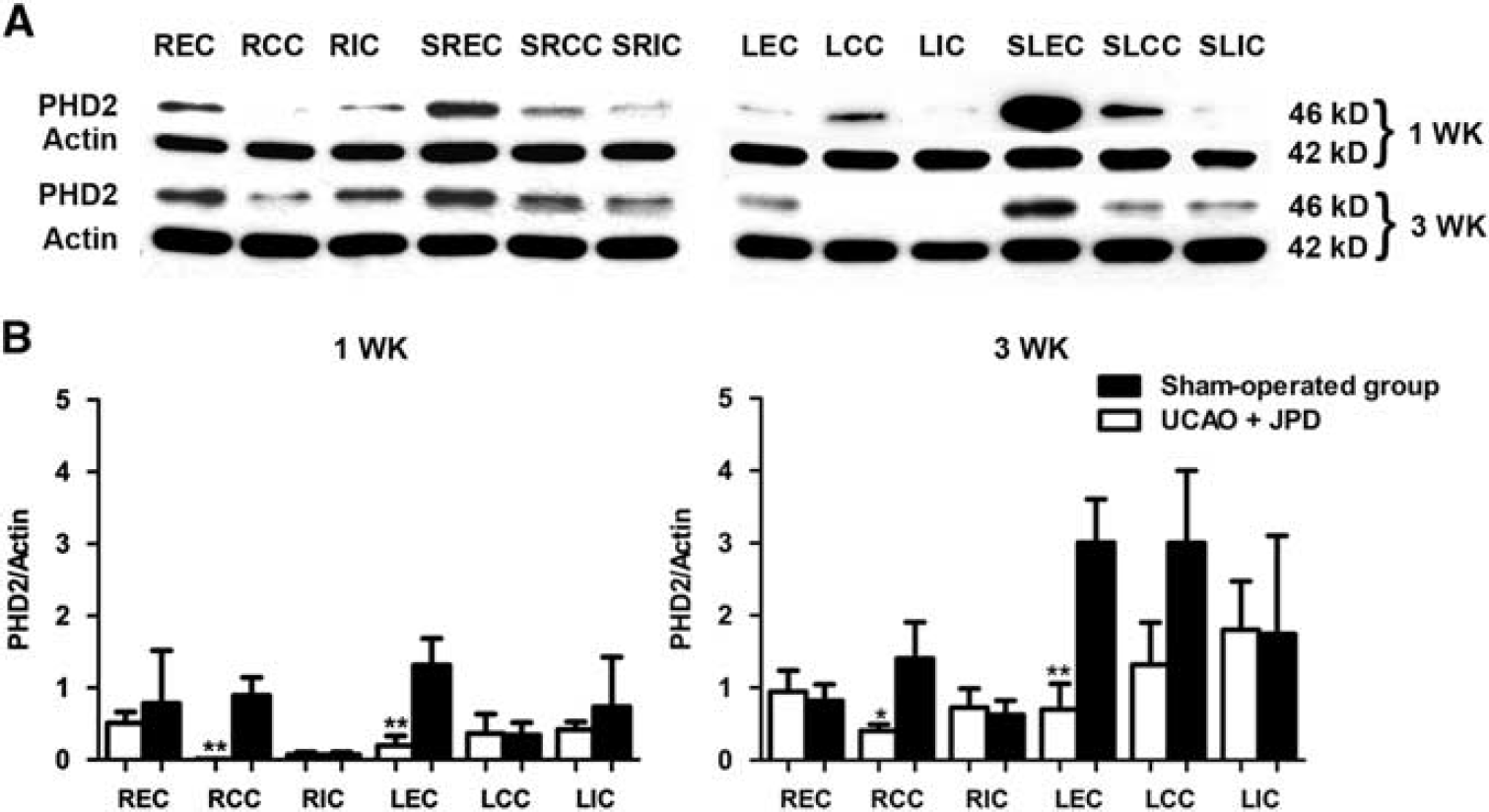
Decreased prolyl hydroxylase-2 (PHD2) in unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD). (
ProMMP-9 was significantly greater in EC and CC in the occluded side and the area of CC in the nonoccluded side of the UCAO/JPD group 1 week after the interventions. At week 3 of UCAO/JPD, proMMP-9 was also increased in the EC and CC areas of the occluded and the area of CC in the nonoccluded brain hemisphere when compared with the sham-operated group (Figure 3A).
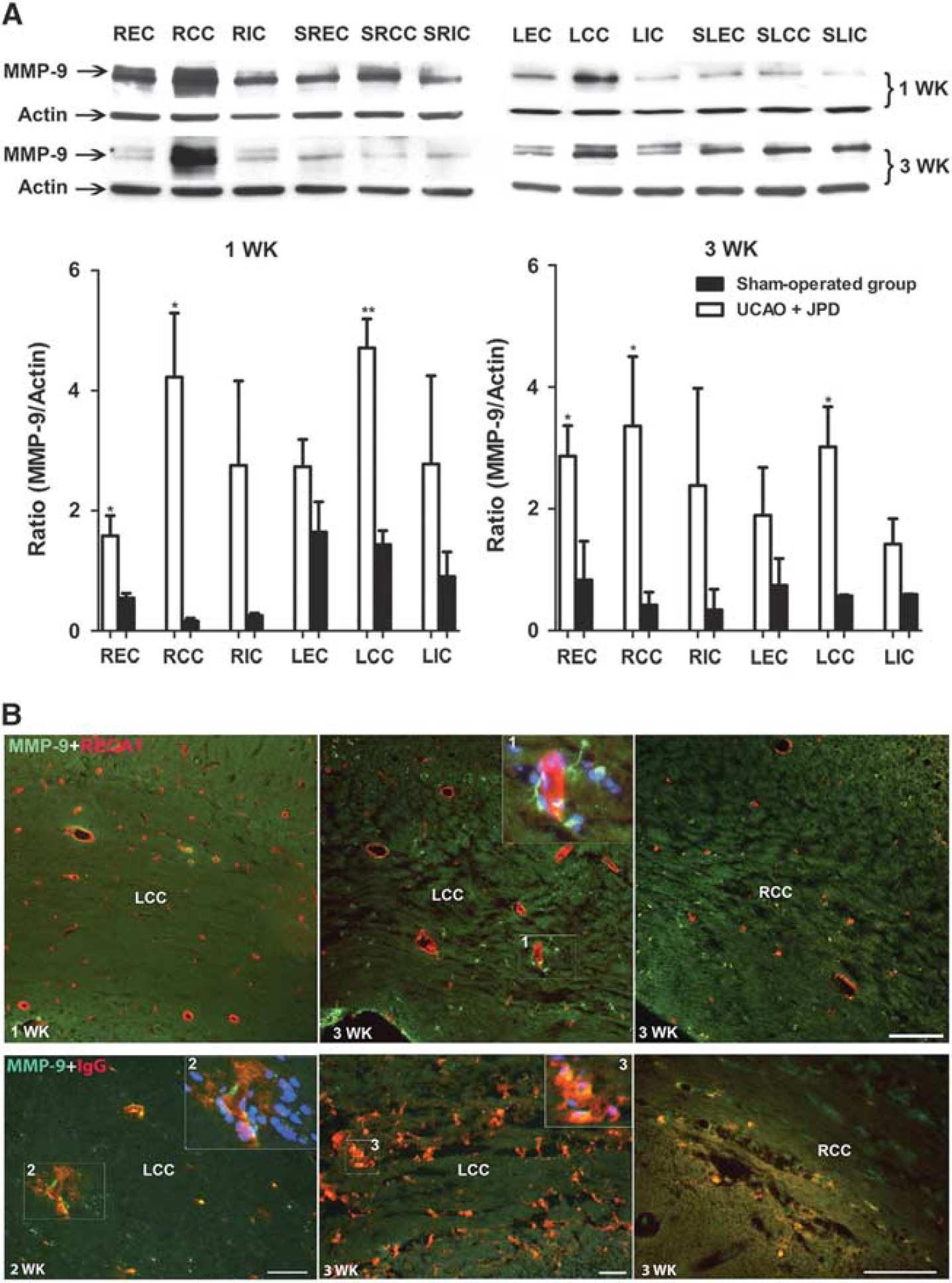
Blood-brain barrier damage associated with matrix metalloproteinase-9 (MMP-9) expression after unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD). (
The BBB integrity was assessed using IgG immunohistochemistry. One week after UCAO/JPD, IgG was not detected in the area of WM (data not shown). IgG staining was detected at week 2 after UCAO/JPD. Strong immunostaining was seen closely associated with blood vessels. Leakage of the IgG was clearly detected in the WM areas 3 weeks after UCAO/JPD (Figure 3B). Colabeling of MMP-9 with IgG was seen in the area of WM, suggesting that MMP-9 may be associated with BBB disruption (Figure 3B).
Immunohistologic staining for MPO showed expression in the WM starting at 1 week and continuing into week 3 after UCAO/JPD when compared with sham-operated group. Double immunofluorescent staining showed that MPO-positive cells were colocalized with MMP-9 (Figure 4). Immunostaining of lymphocytes confirmed the T-cell presence within and surrounding the blood vessels 1 week after UCAO/JPD. By week 3, T cells infiltrated WM (Figure 5).
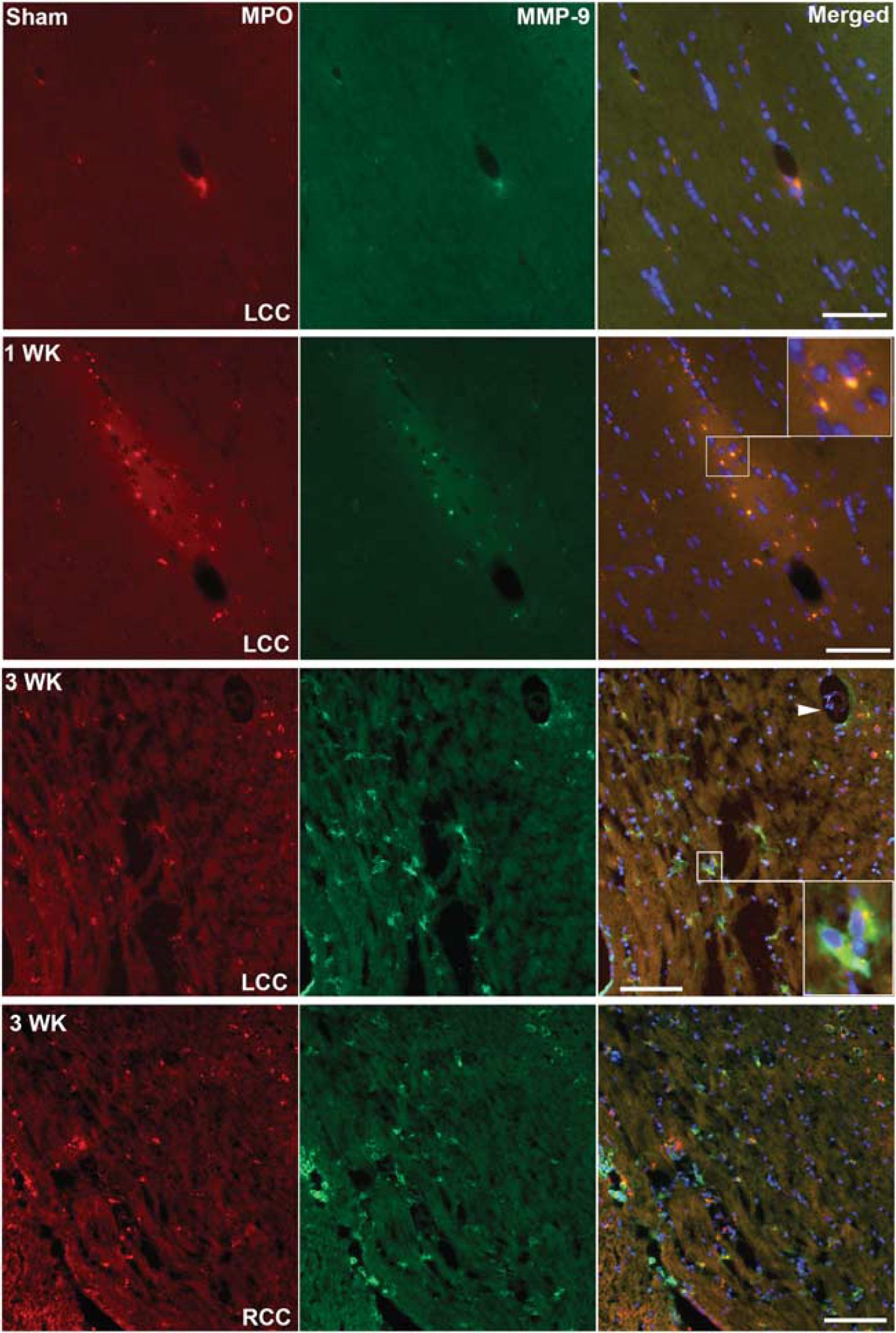
Increased myeloperoxidase (MPO) and matrix metalloproteinase-9 (MMP-9) expression 1 and 3 weeks after unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD). Double staining with antibodies against MMP-9 (green) and against MPO (red) shows neutrophils expressing MMP-9 after UCAO/JPD in the area of white matter (WM). Inserts show the colocalization of MMP-9 and MPO. Arrowhead indicates vessel surrounded with expression of MMP-9 and EPO. Scale bar = 100 μm.
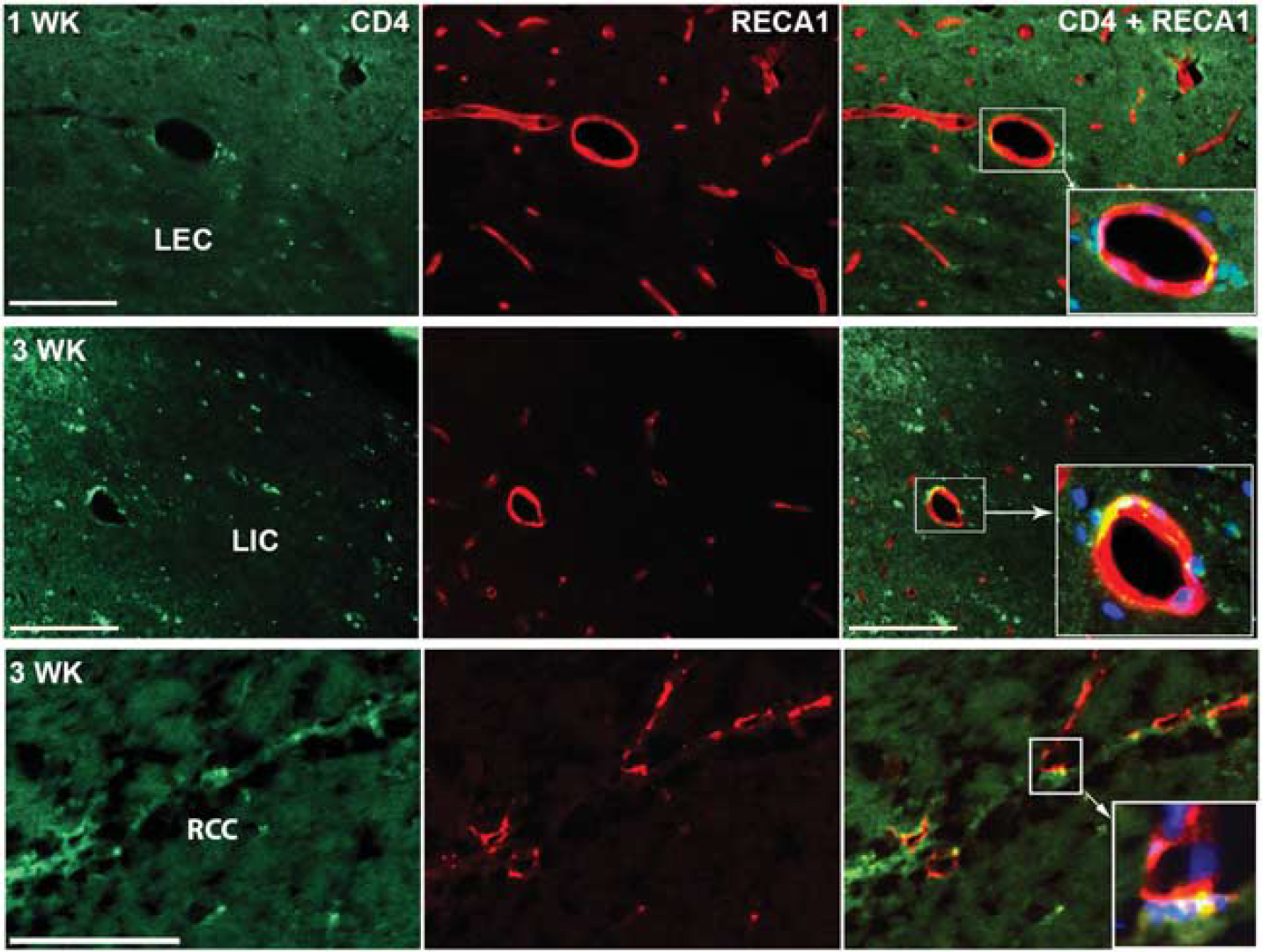
Tissue-infiltrating T cells in white-matter (WM) area after unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD). CD4 is found in the vessel wall in WM area that closely surrounds vessels. RECA1 (red), CD4 (green), 4′-6-diamidino-2-phenyllinidole (DAPI) (blue). Scale bar = 100 μm.
Anatomic T2 MRI illustrated the lesions present in the WM in untreated animals. Treatment with minocycline significantly reduced lesion size in WM compared with vehicle-treated group. Minocycline administration significantly lowered ADC values compared with vehicle-treated rats, suggesting a restoration of WM integrity. Fractional anisotropy, another measure of diffusivity that is increased in the axial direction in intact WM, increased significantly in the minocycline-treated group compared with the decreased values seen in vehicle-treated group. Increased FA suggests WM fiber tract integrity, which is consistent with reduced WM damage with minocycline. Cerebral blood flow as measured with ASL was significantly increased in the minocycline-treated animals compared with controls (Figure 6).
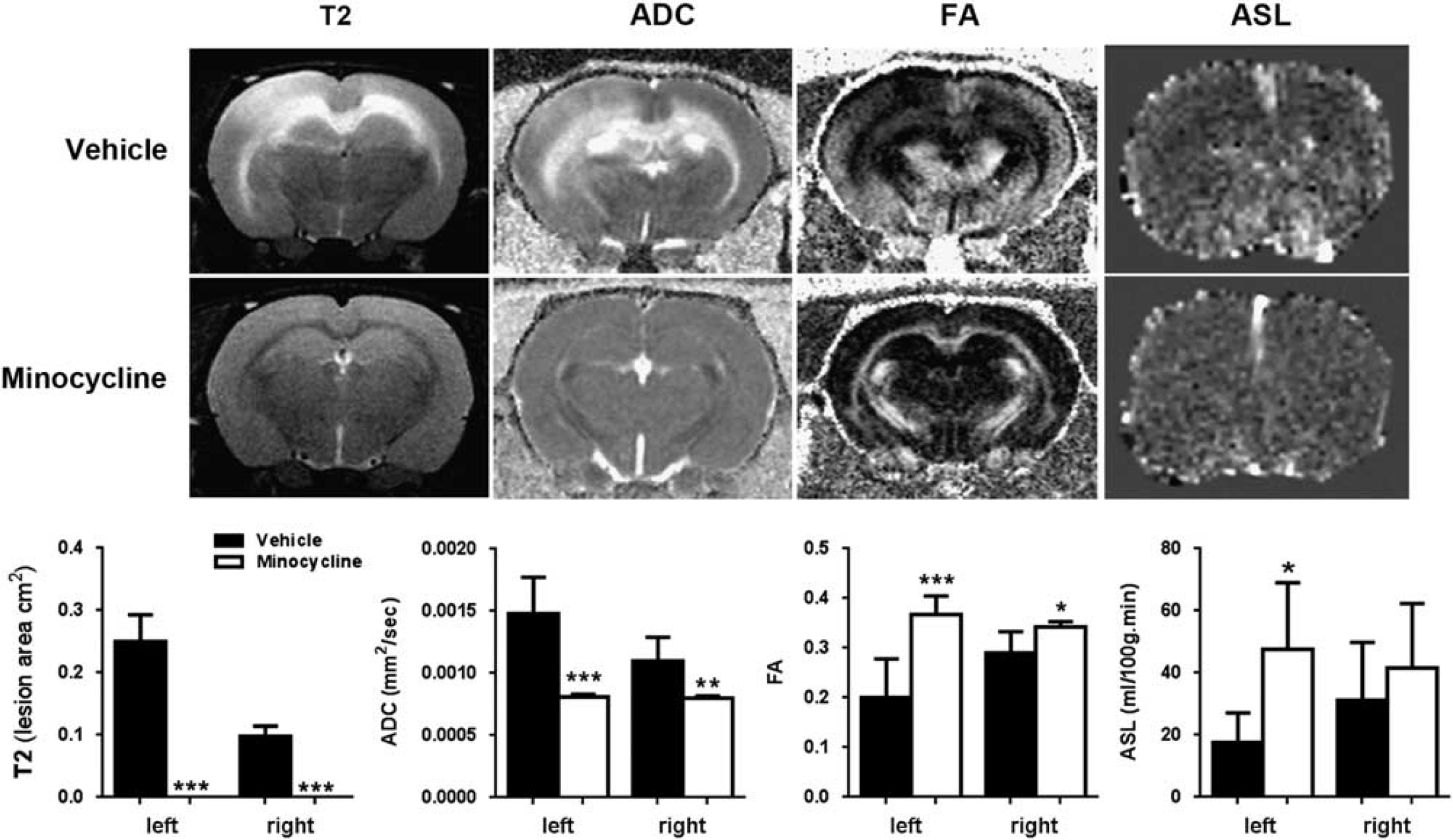
Magnetic resonance imaging (MRI) studies of white-matter (WM) injury, diffusion, and cerebral blood flow. Representative T2-weighted images, apparent diffusion coefficient (ADC), and fractional anisotropy (FA), and arterial spin labeling (ASL) in left and right hemispheres from the minocycline treated and unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD) groups at 9 weeks after UCAO/JPD. Graphs show that the changes in all four parameters are significantly improved with minocycline treatment. *P < 0.05, **P < 0.01, ***P < 0.001, n = 5 in each group.
Cognitive testing in the MWM showed that the minocycline-treated group had significantly higher escape latencies during the acquisition trials than the sham-operated group on day 4 (Figure 7A). Memory testing in the probe trial, in which the platform was removed, requiring rats to recall the location of the platform in the southwest quadrant with distal cues, showed that the minocycline-treated group had intact memory, evidenced by greater time spent in the southwest quadrant, while the vehicle-treated group spent significantly less time, revealing memory impairments (Figure 7B). Furthermore, swimming speed during 4 days of the acquisition trial was similar (Figure 7C), indicating that deficits in performance by the vehicle-treated group were not due to poor motor function. Additionally, there were no group differences in the latencies to find a platform with a visual cue, indicating that visual function was normal (Figure 7D).
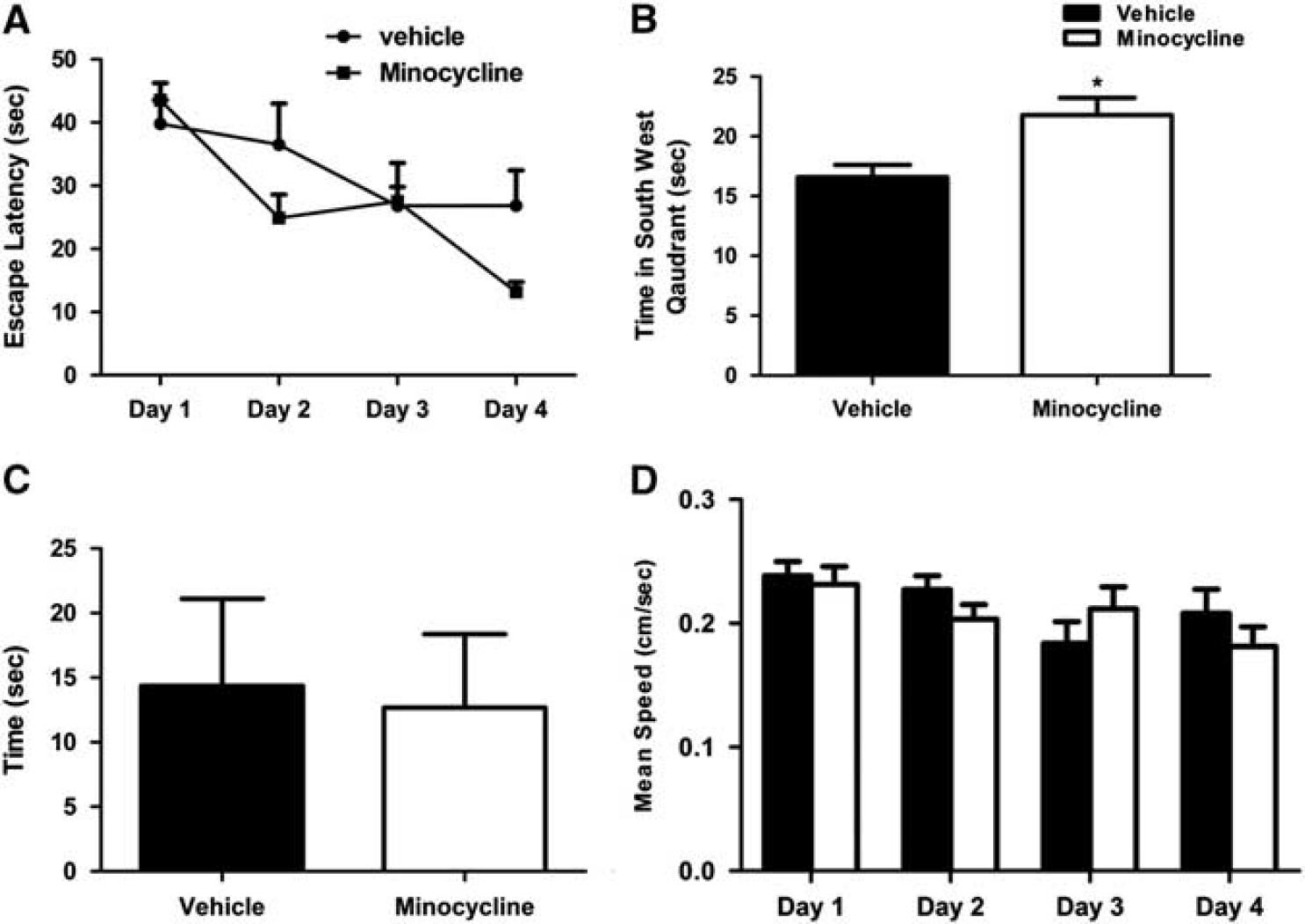
Effect of minocycline on unilateral carotid artery occlusion (UCAO)/Japanese permissive diet (JPD)-induced cognitive dysfunction in Morris water maze test. (
DISCUSSION
We showed that by the first week after initiation of UCAO/JPD in 12-week-old SHR/SP that the WM has increased HIF-1α and reduced PHD2, which is consistent with tissue hypoxia. By the third week, there was MMP-9-associated IgG leakage around blood vessels that accompanied an immunologic response characterized by influx of neutrophils and T lymphocytes. These changes indicate that in chronically hypertensive rats, there was impaired tissue oxygenation, inflammation, and infiltration of T lymphocytes and neutrophils, which was associated with extensive WM damage as shown by injury patterns on multimodal MRI and behavioral impairments in the MWM. Minocycline, which has anti-inflammatory actions, including MMP-9 inhibition, effectively restored WM integrity as shown by MRI measurements of lesion size, ADC, FA, and ASL, and more importantly, improved behavior.
Hypoxia inducible factor-1α induces a cassette of genes involved in erythropoiesis, angiogenesis, and apoptosis. 9 Oxygen levels control expression of HIF-1α through the enzymatic activity of the PHDs. 29 Under low oxygen levels, hydroxylation of HIF-1α ceases, leading to accumulation of HIF-1α, which drives expression of cassettes of genes involved in tissue damage and repair. Our data show that SHR/SP with UCAO/JPD have increased expression of HIF-1α in all WM regions, both ipsilateral and contralateral to carotid occlusion, suggesting that hypoxia has widely compromised hydroxylation by PHDs and degradation of HIF-1α. These results complement our recent electron paramagenetic resonance study, which showed hypoxia in WM in regions that corresponded to those showing HIF-1α expression. Electron paramagnetic resonance showed an increase in tissue oxygen levels, beginning at 9 weeks of age, reaching hyperoxic levels at 12 weeks before precipitously falling into the hypoxic range from 13 weeks to death at 16 weeks. 22 Thus, the direct measurements of oxygen in tissue corroborated the increase in HIF-1α in this study, which has been shown to also occur in acute stroke. 30
Prolyl hydroxylase has a complex pattern of expression. During hypoxia, levels of PHD may remain the same or fall. We observed an apparent reduction in the levels of PHD2, which could explain the observed rise in HIF-1α. We speculate that chronic hypoxia in the WM could result in decreased PHD2 expression and activity, which in addition to reduced oxygen content, could result in the accumulation of HIF-1α. In some samples, decreased levels of PHD2 did not correlate with the increased levels of HIF-1α. These results are in agreement with the general concept that during hypoxia, HIF-1α stabilization is mostly associated with low oxygen levels, rather than with the downregulation/dysfunction of PHD2. Different levels of PHD2 in the examined samples could be explained by region-specific pattern of expression (for example, consistently high levels in EC of all animals), as well as by different effects of hypoxia on PHD2 in different WM regions. An important caveat is that further studies of gene transcription products will be needed to confirm that the HIF-1α is in an active form.
We observed HIF-1α expression in astrocytes, which agrees with previous investigations demonstrating HIF-1α in astrocytes after hypoxia, where it may protect hypoxic astrocytes from glutamate toxicity. 31 HIF-1α has been shown to have a key role in brain edema formation and BBB disruption via a molecular pathway involving MMP-9 in brain trauma, but this is the first report to link the two in chronic vascular disease. 16 Previous investigations have showed that hypoxia-induced MMP-9 expression leads to vascular leakage, which could be reduced by MMP inhibition; in mice exposed to hypoxia, the resulting edema formation was mediated by MMP-9-dependent tight junction rearrangement, which was blocked with an MMP-9 inhibitor. 17 The finding of neutrophils and T cells associated with MMP-mediated BBB disruption provides strong evidence of the major role played by inflammation in the SHR/SP UCAO/JPD animal model of VCI. The increase in the expression of CD4+ cells observed in our study indicates that T cells are involved in VCI pathology.26–28,32 We observed that minocycline had several beneficial effects compared with vehicle-treated controls: treated animals survived 4 weeks longer, showed improvement in multimodal MRI, and behavior. The vehicle, DMSO, also prolonged survival, which is consistent with its positive effect on ischemia from its antioxidant effects, but the improvement in survival was not mirrored by MRI.33,34 Improvement in MRI parameters corresponded with improved behavior in minocycline-treated animals. We observed an increase in ADC in the untreated animals that was corrected with minocycline; increased ADC may be seen in vasogenic edema, which was seen as an increase in WM lesions in T2-weighted signal.7,35 Fractional anisotropy is composed of radial and axial diffusivity. Untreated animals showed reduced FA, which could be explained by an increase in radial diffusivity secondary to loss of WM tissue integrity. Treatment with minocycline increased FA compared with vehicle treatment, which suggests that minocycline treatment resulted in an increase in axial diffusivity and a decrease in radial diffusivity, which would be consistent with WM protection by minocycline. Finally, ASL measurements of cerebral blood flow, which were reduced in untreated rats, were restored with minocycline treatment.
Several animal models are used to study chronic cerebrovascular disease. Bilateral carotid occlusion in the normotensive rat results in hypoxic hypoperfusion with injury to the WM. 36 Placing a small coil around the carotid arteries of the mouse restricts blood flow, leading to WM damage. 8 However, hypertensive small vessel disease seen in SHR/SP most closely resembles the pathologic changes in VCI patients with WM disease. Unmanipulated SHR/SP can survive for a year or longer, depending on the genetic strain. Without dietary manipulation they fail to show MRI changes for up to 9 months and have normal histology. However, when they are feed a low protein, high-salt diet beginning at the sixth week of life, they develop WM damage and die by 16 to 20 weeks. 35 A recent study showed that SHR/SP with JPD and added salt developed increased permeability before development of intracerebral bleeding. 37 We combined the JPD with occlusion of one carotid to accelerate the pathologic changes in the WM; we began the abnormal diet at the twelve rather than the sixth week, resulting WM damage that was generally seen before strokes. 21 While none of the rodent models replicate fully the human condition, the SHP/SP with the abnormal diet shows similar pathology in an accelerated time frame.
An important feature of the SHR/SP/UCAO/JPD animal model for VCI is that the damage to the WM occurs over time, which is similar to VCI patients, but with a shorter time span in the rat. Hypertension leads to damage to the small vessels, which limits cerebral perfusion and reduces the ability of the cerebral blood vessels to dilate during hypoxia to increase tissue oxygenation. This results in hypoxic hypoperfusion, which is occurring with long-standing, poorly treated hypertension in humans. 38 We propose that the development of WM damage in SHR/SP/UCAO/JPD is a multiphase process with several sequential events required to produce the final tissue injury. This process can be visualized as a series of ‘hits': (1) initially, the chronic hypertension causes vessel lumen narrowing and loss of normal autoregulation of the blood vessels, which leads to hypoperfusion and hypoxia in the vulnerable deep WM; (2) hypoxia leads to production of HIF-1α, inducing an inflammatory response; and (3) finally, there is production of MMPs, disrupting the tight junctions and extracellular matrix with opening of the BBB. Vasogenic edema accelerates tissue hypoxia, damaging myelin and leading to oligodendrocyte death. Hypertensive vascular disease is only one of the causes of WM injury in humans, and other animal models will be needed to fully explain the WM damage. The benefit of this chronic inflammatory model is that noninvasive MRI and MWM can be used to develop potential treatments for VCI.
AUTHOR CONTRIBUTIONS
FJ designed and carried out the studies and participated in writing the report. YY designed the studies, carried out the experiments and participated in the writing of the report. JT provided technical assistance and helped with the manuscript preparation. TR helped with the HIF studies and the writing. GR obtained the funding, designed the experiments, and participated in the writing of the manuscript.
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.
Footnotes
ACKNOWLEDGMENTS
The study was supported by grants from the NIH (RO1NS045847) to GR.