
Abstract
Gamma oscillations (~30 to 100 Hz) provide a fundamental mechanism of information processing during sensory perception, motor behavior, and memory formation by coordination of neuronal activity in networks of the hippocampus and neocortex. We review the cellular mechanisms of gamma oscillations about the underlying neuroenergetics, i.e., high oxygen consumption rate and exquisite sensitivity to metabolic stress during hypoxia or poisoning of mitochondrial oxidative phosphorylation. Gamma oscillations emerge from the precise synaptic interactions of excitatory pyramidal cells and inhibitory GABAergic interneurons. In particular, specialized interneurons such as parvalbumin-positive basket cells generate action potentials at high frequency (‘fast-spiking’) and synchronize the activity of numerous pyramidal cells by rhythmic inhibition (‘clockwork’). As prerequisites, fast-spiking interneurons have unique electrophysiological properties and particularly high energy utilization, which is reflected in the ultrastructure by enrichment with mitochondria and cytochrome c oxidase, most likely needed for extensive membrane ion transport and γ-aminobutyric acid metabolism. This supports the hypothesis that highly energized fast-spiking interneurons are a central element for cortical information processing and may be critical for cognitive decline when energy supply becomes limited (‘interneuron energy hypothesis’). As a clinical perspective, we discuss the functional consequences of metabolic and oxidative stress in fast-spiking interneurons in aging, ischemia, Alzheimer's disease, and schizophrenia.
Keywords
INTRODUCTION
Research on brain energy metabolism in the neocortex and the hippocampus has made major progress with regard to oxidative metabolism, energy utilization, and survival of neurons.1–6 Notably, most of this research has focused on excitatory neurons such as glutamatergic pyramidal cells that release neurotransmitter, glutamate. These principal projection neurons are generally thought to compute, transfer, store, and retrieve information, which underlies higher brain functions such as sensory perception, motor behavior, and memory formation.7–9
Neuronal information processing, however, requires precise spatial and temporal coordination of pyramidal cell activities in cortical networks.9–11 Different classes of network oscillations provide such a fundamental mechanism by enabling coordinated synchronous activity during normal brain function.9,12,13 There are multiple different classes of network oscillations, which span a wide range of frequencies from ~0.05 to almost 1,000 Hz and which define different cognitive and behavioral states. 12 Dominant patterns in cortical networks comprise theta- (4 to 12 Hz), beta- (13 to 30 Hz) and gamma- (~30 to 100 Hz) frequency oscillations, which synchronize the action potential generation (‘spiking’) of principal cells with great precision, sometimes over large distances of corticocortical connections.10,13–15
Importantly, inhibitory interneurons have a key role in organizing these highly coordinated patterns of activity. In cortical networks, inhibition is mainly mediated by neurotransmitter, γ-aminobutyric acid (GABA), and research during the past decade has revealed an astonishing heterogeneity of GABAergic interneurons at the molecular, morphologic and functional level.16,17 This raises the fundamental question of electrophysiological and neuroenergetical properties of GABAergic interneurons, including the impact on cortical information processing under physiologic conditions as well as pathologic conditions such as ischemia. However, GABAergic interneurons have attracted little attention about brain energy metabolism so far. 18
Here, we summarize and discuss studies on the cellular mechanisms and neuroenergetics of hippocampal gamma oscillations (~30 to 100 Hz). Gamma oscillations have a strong relationship to higher brain functions.7,11,13 Despite occurring in many cortical regions, gamma oscillations have been particularly well studied in the hippocampus that has become an important model system in cognitive neuroscience for several reasons.10,16,19–21 First, hippocampal gamma oscillations are evoked during specific behavioral states such as exploration. Second, as the hippocampus has a central role for spatial navigation and episodic memory, the relevance of network oscillations for encoding, storage, and retrieval of information can be studied. Third, extracellularly recorded hippocampal gamma oscillations show larger amplitudes because of the more simple laminated architecture of the hippocampus compared with the neocortex. We will focus on hippocampal gamma oscillations in the present review, combining mechanistic results from in vitro and in vivo studies in rodents. Where available, we will include data from the neocortex and from humans.
Collectively, the available evidence indicates that information processing in the cortex critically depends on the activity of a specific type of inhibitory interneurons, so-called ‘fast-spiking’ interneurons that generate action potentials at high frequencies. At the same time, these interneurons feature particularly high energy utilization, which puts them into a critical position for the maintenance or failure of cognitive functions when metabolic stress occurs (‘interneuron energy hypothesis’).
GAMMA OSCILLATIONS AND CORTICAL INFORMATION PROCESSING
Gamma oscillations have been found in many mammalian brain regions, including the hippocampus and the neocortex.9,13,20 They are characterized by rhythmic and synchronous fluctuations of the membrane potential of most or all neurons in a neuronal network, with a characteristic frequency domain of ~30 to 100 Hz.11,22 These subthreshold fluctuations are generated by a complex interplay between excitatory pyramidal cells and inhibitory GABAergic interneurons.16,19,23,24 Functionally, gamma oscillations in neuronal networks bind neurons into a common temporal regime during higher brain functions like motor behavior, sensory perception, or memory formation.7,25–28 The synchronizing effect of gamma oscillations permits the coordinated activation of defined sets of neurons, which constitute functional ensembles—the putative information-carrying multicellular subsets of neuronal networks.9,11,29 Moreover, the precise timing of action potentials is central for use-dependent synaptic plasticity and, thus, supports learning and memory formation.13,20,30,31
In neuronal networks in vivo, gamma oscillations occur transiently on the hundred millisecond time scale upon sensory input or during specific cognitive tasks. In the human brain and dependent on the task, however, they can last for prolonged times in the range of minutes.28,32 This aspect is important for the validation of in vitro models, most of which feature persistent gamma oscillations.8,9 Another characteristic of gamma oscillations is the synchronization of remote cortical networks. This peculiar phenomenon has been proposed to underlie binding of the distributed neuronal coding, and may thus form a crucial prerequisite for the unity of perception, attention and, perhaps, consciousness.10,13,33 Of course, the fact that research on neuronal network phenomena has focused on gamma oscillations does not withstand the importance of slower network oscillations for higher brain functions.10,12 Interestingly, high-frequency network oscillations are consistently present during cognitive tasks that require attention or consciousness, and they are often correlated with adaptations in local cerebral blood flow.13,34
CELLULAR MECHANISMS OF GAMMA OSCILLATIONS: INSIGHT FROM IN VITRO MODELS
In vivo studies have yielded valuable insights into the relationships between cortical gamma oscillations and behavior, including the identification of the involved neuronal subpopulations.13,22,24,35,36 However, detailed experimental analyses of the cellular and subcellular mechanisms underlying gamma oscillations have been widely performed in vitro, particularly in neuronal networks of the hippocampus.16,17,19,37 Rodent hippocampal slices express various patterns of naturally occurring network oscillations and permit high-resolution recordings with electrophysiological methods, live-cell imaging techniques and oxygen sensor microelectrodes under well-defined recording conditions.38–41 This in vitro preparation also provides a reasonable model for examining energy metabolism during different forms of neuronal network activation.42–45
In the hippocampal slices, persistent gamma oscillations can be reliably induced by bath application of cholinergic or glutamatergic receptor agonists at nanomolar to low micromolar concentrations (Figures 1A and 1B). The most widespread models use acetylcholine or carbachol for activation of metabotropic cholinergic receptors, or kainate (kainic acid) for activation of ionotropic glutamatergic receptors.40,41,46,47 Pharmacologically induced hippocampal gamma oscillations in vitro are most prominent in the neuronal network of subfield CA3, weaker in subfield CA1, and less prominent or absent in the dentate gyrus.16,41 They can occur in the presence or in the absence of theta oscillations,40,41 similar to the pattern of gamma oscillations in vivo.8,30,48 Alternative models are based on the induction of transient phases of gamma oscillations: local tetanic electrical stimulation, topic application of potassium-rich solution, or application of metabotropic glutamate receptor agonists.16,38,49 However, electrical stimulation of neuronal tissue can only partially mimic the features of naturally occurring network oscillations. 9 Nonetheless, electrical stimulation of neuronal tissue at different frequencies and intensities is a very useful tool for well-defined, transient neuronal activation in experimental studies on ion homeostasis and energy metabolism.42,43,45 All in vitro models of hippocampal gamma oscillations share a critical role for rhythmic, transient activation of GABAA receptors because the oscillations are all blocked by GABAergic receptor antagonists such as bicuculline. They do, however, differ in the relative contribution of glutamatergic synaptic transmission: while gamma oscillations that are induced by metabotropic glutamate receptor agonists or kainate persist in the presence of AMPA receptor antagonists, carbachol-induced gamma oscillations depend on the function of AMPA receptor-mediated excitatory postsynaptic potentials.8,16,19 Studies using transgenic mice with cell-specific deletion of glutamate receptor subunits show that synaptic excitation of fast-spiking interneurons is required for generation of normal gamma oscillations in vivo.50,51
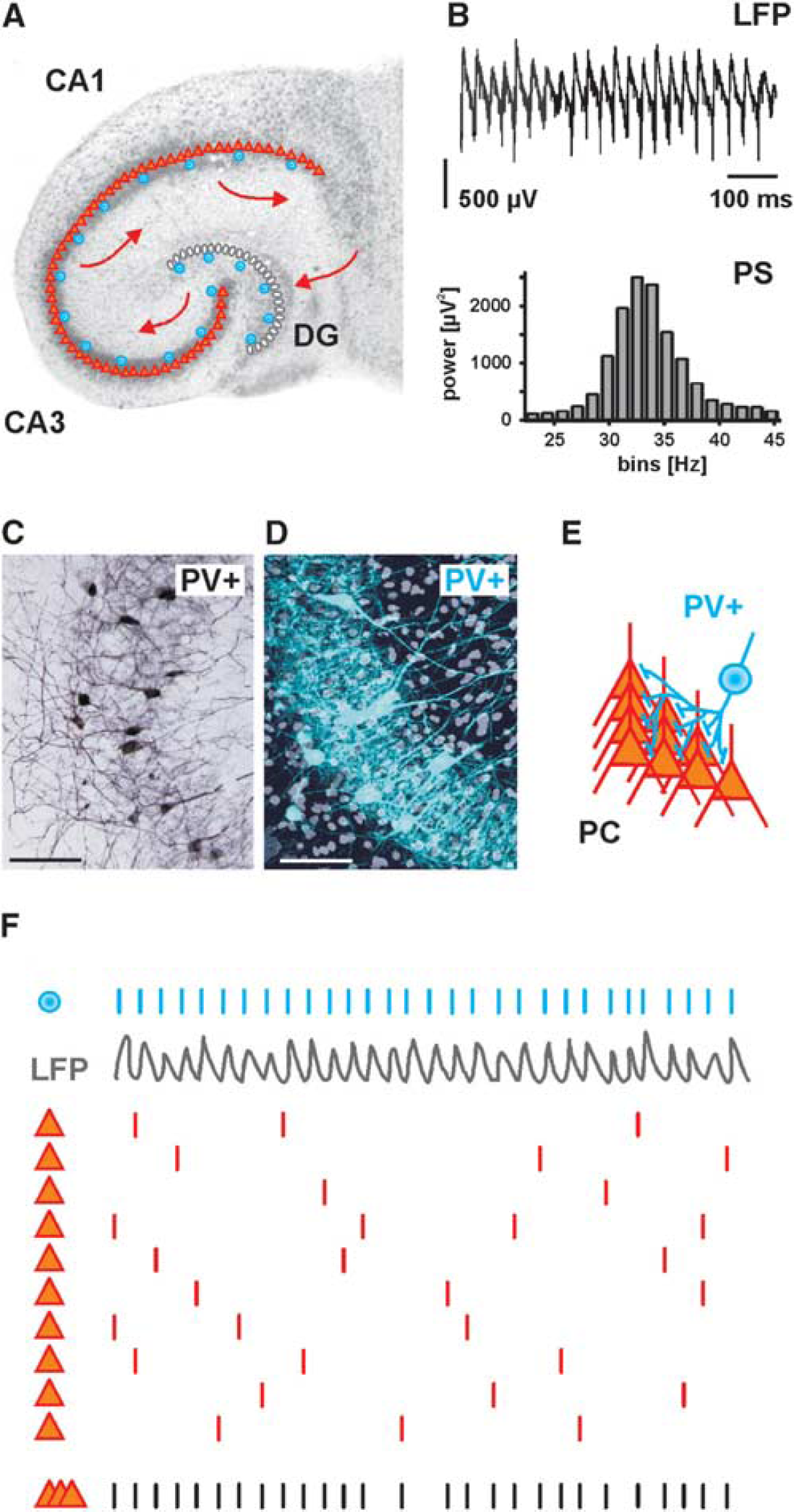
Gamma oscillations are exemplified from the CA3 network of the hippocampus. (
The indispensable role of phasic GABAergic inhibition for generation of gamma oscillations is underlined by the fact that local field potentials in the gamma-frequency band are dominated by inhibitory postsynaptic potentials (IPSPs), rather than action potentials or excitatory postsynaptic potentials.17,52,53 It is also in line with the models of gamma oscillations in neuronal networks that emphasize the importance of rhythmic and synchronous inhibition of both pyramidal cells and interneurons, which finally generates alternating phases of enhanced or decreased probability of spiking.9,54 Initial models argued that the cycle of gamma oscillations (~25 ms for 40 Hz) is dominated by the duration of slow IPSPs. Subsequent direct electrophysiological measurements revealed faster kinetics of IPSPs than compatible with the gamma cycle. 16 This has led to the proposals of modified models that include conduction delays in axons of interneurons but have not changed the basic concept of rhythmic inhibition as a key factor for gamma oscillations. 16 Indeed, synchronized inhibition of multiple interneurons and pyramidal cells, followed by feedback excitation of interneurons is the common core concept of the existing network models on gamma oscillations.9,55
HIGH ENERGY UTILIZATION DURING HIPPOCAMPAL GAMMA OSCILLATIONS
Recent studies have started to directly address the energy utilization of different activity states in neuronal networks using slice preparations from mouse and rat. Recordings of local field potential and interstitial partial oxygen pressure (pO2) with high spatiotemporal resolution revealed a positive correlation between the power of gamma oscillations and oxygen consumption.
41
The power spectrum reveals the strength of potential variations (energy) as a function of frequency and, thus, the contribution of different frequencies to a complex oscillation40,41,56 (Figure 1B). Surprisingly, oxygen consumption during gamma oscillations reached similar levels as observed during pathologic hyperactive states, i.e., seizure-like events. High-resolution depth profiles of local pO2 and mathematical modeling of convective transport, diffusion- and activity-dependent consumption of oxygen suggest a 2.2-fold increase in the oxygen consumption rate during gamma oscillations compared with asynchronous neuronal network activity (‘baseline’ activity) in vitro, indicating that gamma oscillations are associated with high energy utilization.
57
This value is higher than observed in vivo where local cerebral metabolic rate of oxygen consumption rarely increases more than 30% during functional activation of cortical regions.58–61 This discrepancy might primarily reflect much lower levels of ‘baseline’ activity and thus ‘resting’ metabolic rates in slice preparations compared with ‘baseline’ activity in vivo. In addition, methodological aspects need to be considered (see below). Support for the notion of high energy utilization during gamma oscillations comes from studies in humans based on positron emission tomography with 2-deoxy-2-[18F] fluoro-
Several methodological aspects need to be considered when comparing the above-cited results from experiments in vitro and in vivo. First, the 2.2-fold increase in oxygen consumption rate in vitro was determined during persistent gamma oscillations in the pyramidal cell layer (perisomatic region) of the CA3 network in the hippocampal slices of the rat. Thus, ‘local’ in the pO2 depth profile refers to a distance of ~0.2 mm 57 while the resolution of functional magnetic resonance imaging technologies is in the order of 1 mm3 and more. Second, the CA3 network is known to be an intrahippocampal generator of high-power gamma oscillations,9,10 and the electrophysiological and neuroenergetical features of this network may not entirely apply to other cortical regions that express gamma oscillations. Third, ‘baseline’ activity without detectable episodes of gamma oscillations might be higher in vivo than in partly deafferentiated slice preparations. For a full understanding of the state-, task-, region-, and lamina-specific energy metabolism, we will need further experimental studies in vitro and in vivo, including electrode-based and optical methods with improved spatiotemporal resolution.18,60
The high energy utilization of neurons during gamma oscillations is most likely caused by increased rates of action potentials and synaptic interactions. In particular, the significant increase in the rates of excitatory postsynaptic potentials and IPSPs during gamma oscillations elicits strong ion fluxes across the neuronal membrane of pyramidal cells and inhibitory interneurons.8,19,70 These ion fluxes tend to dissipate the gradients of sodium, calcium, potassium, and chloride ions, thus utilizing potential energy. In order to maintain physiologic levels of excitability, these ionic gradients have to be continuously restored by ion pumps such as Na+/K+-ATPase and Ca2+-ATPase as well as transporters such as Na+/Ca2+-exchanger, Na+/H+-exchanger, K+/Cl−-cotransporter, and Na+/K+/Cl−-cotransporter. These transport processes are directed against equilibrium and finally fueled by adenosine-5′-triphosphate (ATP) that is generated mainly by oxidative phosphorylation in the neuronal mitochondria.2,3,70 This was recently supported by fluorescence imaging of nicotinamide adenine dinucleotide (phosphate) [NAD(P)H] and flavin adenine dinucleotide, i.e., by monitoring the cellular and mitochondrial redox state. Hippocampal gamma oscillations in vitro were associated with near-limit utilization of mitochondrial oxidative capacity, in line with oxygen consumption data.41,57 Interestingly, subunits of mitochondrial complex I are strongly expressed in CA3, the hippocampal network with highest gamma oscillation power and oxygen consumption.41,71 Complex I (NADH:ubiquinone oxidoreductase) is a large protein composed of up to 45 individual subunits that is part of the mitochondrial respiratory chain. 72 It seems to have a key role for oxidative phosphorylation under normal conditions as well as in neurodegenerative diseases. 73 The peculiar pattern of complex I subunit expression in the hippocampus likely reflects special enzymatic properties of neuronal mitochondria in the CA3 network, in agreement with the high cytochrome c oxidase activities in this region.70,74 These data also support the notion that mitochondrial DNA genotype and nuclear-mitochondrial interactions are important determinants of higher cognitive tasks. 75
It is currently hard to quantify the relative contribution of excitatory and inhibitory neurons to energy utilization during gamma oscillations. Owing to the lack of cell-specific data and in the interest of simplicity, global energy budgets for gray matter have been based on glutamatergic neurons that provide 85% to 90% of the neuronal population in the cortical circuits.3,18 Based on the assumption of an average spiking rate of 4 Hz and a ‘sodium entry ratio’ of 1.24 during the action potential, the updated calculations predict energy costs in the cerebral cortex of ~25% for housekeeping, 15% for resting potentials, 16% for action potentials, and 44% for synaptic processes.3,76 In primates, the synaptic costs might be even larger because of the higher numbers of synapses per neuron. 76 Other studies also suggested that the major neuronal energy utilization is for excitatory synaptic signaling (neuronal input) rather than action potentials of the axon (neuronal output).60,77–79 This might be reflected by the strong linear correlation between synaptic activities, oxygen metabolism, and cerebral blood flow in vivo, at least for some experimental stimulation paradigms.61,80 In contrast to the excitatory projection cells, estimates about energy costs of inhibitory interneurons are widely lacking because of insufficient experimental data and cell-specific neuroenergetic knowledge.18,76 Few studies from hippocampus and neocortex provide first evidence that (i) glucose metabolism is increased during long-term recurrent inhibition of hippocampal pyramidal cells, 81 (ii) the contribution of GABA to the glutamate/GABA-glutamine cycle might account for 10% to 15% of the total oxidative metabolism 82 and (iii) glucose metabolism might be significantly stronger in GABAergic neurons than in glutamatergic neurons as revealed by combining high-resolution 2-deoxyglucose and immunohistochemistry. 83 For the latter technically advanced study with single-cell resolution, the methodological obstacles such as adequate label retention during immunohistochemical processing that may limit interpretation have been discussed.83,84 For the remainder of this review, we will summarize current knowledge on the role of interneurons in cortical high-frequency oscillations, in particular, gamma oscillations, and on the possible neuroenergetical and pathophysiological implications of high energy utilization in a specific subset of these GABAergic cells, i.e., fast-spiking, parvalbumin-positive (PV+) interneurons.
HIPPOCAMPAL GAMMA OSCILLATIONS AND FAST-SPIKING INHIBITORY INTERNEURONS
Inhibitory interneurons represent highly diverse neuronal subpopulations in both the hippocampus and neocortex. In the rodent hippocampus, GABAergic interneurons comprise about 10% to 15% of the entire neuronal population, and at least 16 distinct types have been identified in the hippocampal CA1 region.23,85 However, from the functional point of view, inhibitory interneurons can be grossly classified into (i) dendritic-targeting interneurons that control synaptic input to principal cells, (ii) perisomatic interneurons that control principal cell spiking, and (iii) interneurons that innervate and control other interneurons. 8
Some dendritic-targeting interneurons such as bistratified and trilaminar GABAergic interneurons were reported to show fast-spiking behavior during hippocampal gamma oscillations.23,86 However, the class of perisomatic inhibitory interneurons has gained much more attention.8,16,19,87 This class comprises two types of basket cells as well as axo-axonic (chandelier) cells. 87 Basket cells that contain cholecystokinin show regular spiking behavior and have been proposed to modulate synchronous neuronal activities as a function of subcortical inputs carrying information about motivation and emotions;52,87 these will not be further discussed in this review. By contrast, basket cells and axo-axonic cells that contain Ca2+-binding protein, parvalbumin (PV+) show fast-spiking behavior8,16,19,88 (Figures 1C–1F). PV+ basket and axo-axonic cells are abundant; they account for about 20% of the GABAergic interneuron population in the hippocampus. Within or adjacent to the principal cell layers, this value even rises to ~50%.85,89 Notably, many of the studies discussed below focused on fast-spiking behavior and parvalbumin content (PV+) for identification. Thus, unless detailed morphologic post hoc analysis was included, the data may originate from basket cells and/or axo-axonic cells. 23
A single PV+ axo-axonic cell provides GABAergic synapses exclusively to the axon initial segments of up to 1,200 pyramidal cells. 23 The characteristics of axo-axonic cells have been less explored but recent evidence suggests that these cells exert highly specialized functions during information processing in cortical networks.52,87,90 For the hippocampus, it was demonstrated in vitro that axo-axonic cells generate action potentials at much higher rate (~100 Hz) compared with the frequency of gamma oscillations (~35 Hz) in the CA3 network, and that they can control the antidromic propagation of ectopically generated spikes from axons into the soma of pyramidal cells during gamma oscillations. 88 Moreover, axo-axonic cells in the CA3 network show significant phase coupling to hippocampal gamma oscillations in vivo, comparable in strength and phase to those of pyramidal cells and PV+ basket cells. 91 In the rat somatosensory cortex, axo-axonic cells have been reported to depolarize pyramidal cells and thus initiate stereotyped series of synaptic events in neuronal networks because of a depolarized reversal potential for axonal relative to perisomatic GABAergic inputs. 92 Further studies are required to clarify the crucial role of axo-axonic cells as suggested by their morphology and strategical positioning.
For the generation of hippocampal gamma oscillations, PV+ basket cells are of special importance (Table 1). These interneurons show extensive mutual connections with strong inhibitory synapses.23,87 Moreover, they are coupled via electrical synapses, thus forming functional interneuron networks.49,93–98 This special feature is well suited to synchronize basket cell spiking with high temporal fidelity in the hippocampus as well as in the neocortex.8,16,24,99 In brief, an action potential exerts a short-lasting depolarization of electrically coupled basket cells, which is immediately curtailed by the subsequent strong IPSP.16,100,101 This mechanism defines a short time window for mutual excitation of PV+ basket cells, thereby generating near-synchronous inhibition of multiple excitatory and inhibitory neurons in the network. These models are also in line with the observed short phase delay (<90°) of interneuron discharges behind pyramidal cells during hippocampal gamma oscillations in vitro.8,9 It has been suggested that a single PV+ basket cell receives excitatory synaptic input from about 2,000 pyramidal cells. 85 However, the highly divergent axonal plexus of basket cells has a mediolateral extent of >700 μm within the hippocampus where it contacts somata and proximal dendrites of pyramidal cells18,37,86,102 This connectivity feature allows synchronous inhibition of a multitude of pyramidal cells; the estimated numbers range from several hundred up to 2,500 cells.85,103,104 Together with the dendritic-targeting interneurons, the pattern of convergence and divergence in hippocampal network connectivity thus reflects the intense feed-forward and feedback control of principal cells by GABAergic inhibition.17,37,101,105,106
Features of fast-spiking inhibitory interneurons and gamma oscillations
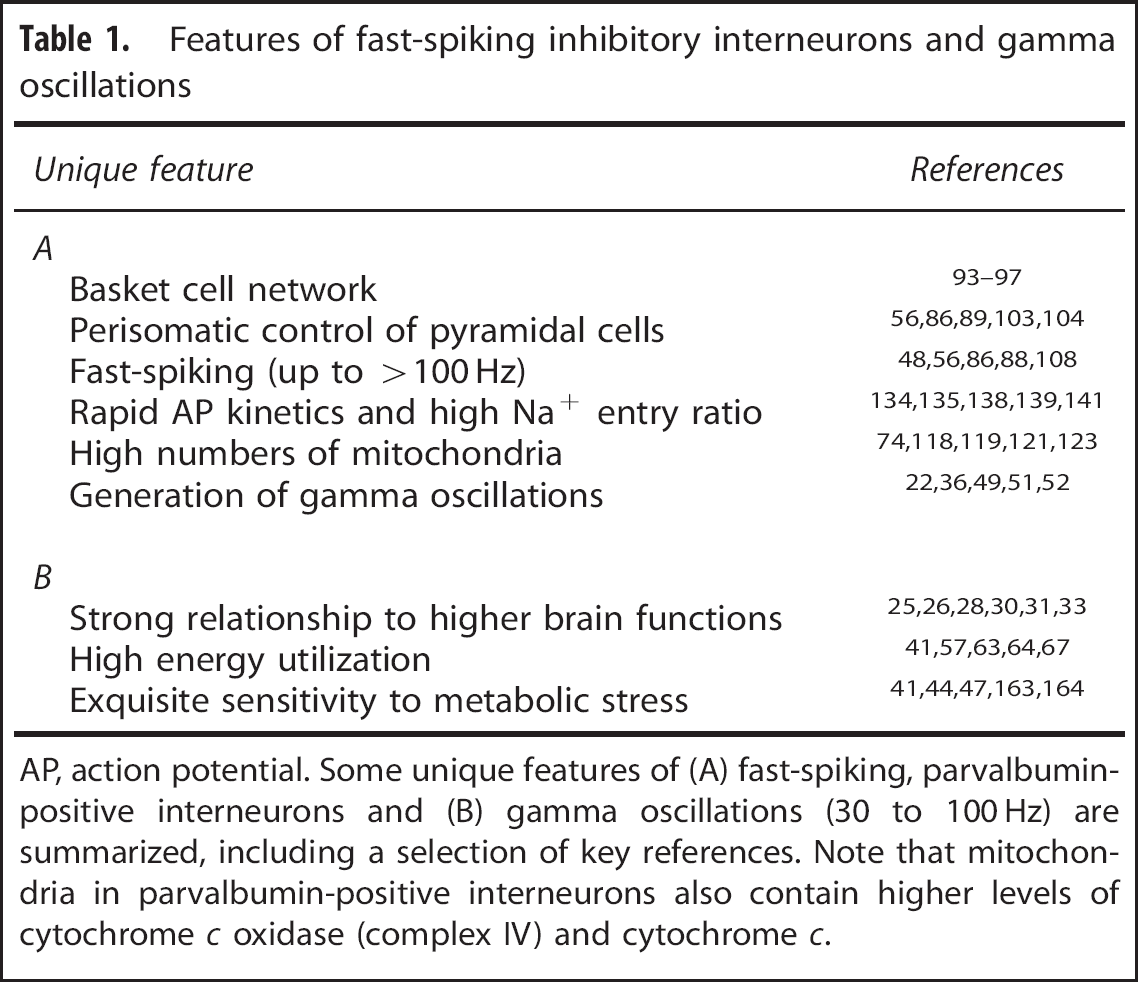
AP, action potential. Some unique features of (A) fast-spiking, parvalbumin-positive interneurons and (B) gamma oscillations (30 to 100 Hz) are summarized, including a selection of key references. Note that mitochondria in parvalbumin-positive interneurons also contain higher levels of cytochrome c oxidase (complex IV) and cytochrome c.
Importantly, PV+ basket cells and axo-axonic cells strongly increase their spiking rate during specific oscillating network states of the hippocampus, especially during theta-, gamma-, and ripple oscillations.22,35,47,91,107 These interneurons are able to generate fast series of action potentials up to several hundred Hertz (hence: ‘fast-spiking’), and they are active in almost every cycle of gamma oscillations (~30 to 100 Hz) in vitro and in vivo.35,48,52,56,86,88,108 The resulting fast periodic release of GABA induces strong rhythmic perisomatic inhibition of almost all local principal cells,9,10,23 which is reflected by the typical gamma oscillations in local field potential recordings.8,48,52 Thus, fast-spiking interneurons are ideally suited to control both pattern and timing of the action potential generation in principal cells (‘clockwork’), which has been demonstrated by experimental data and mathematical modeling.29,37,38,102,107 Based on these properties, fast-spiking, PV+ interneurons establish a temporal matrix for information processing in the cortex (Figures 1F and 2A). This notion has been underlined by elegant in vivo studies in mice showing that selective genetic modification of glutamate receptor subtypes in fast-spiking, PV+ interneurons strongly affects power and synchrony of gamma oscillations, together with impaired hippocampus-dependent cognitive functions.50,51 Similar to the hippocampus, fast-spiking interneurons control pyramidal cell activity during gamma oscillations in the neocortex and thus promote neocortical information processing in vitro and in vivo.36,99,109–111 Pyramidal cells, in contrast, show strong action potential frequency accommodation and have typical spiking rates around 1 to 3 Hz during hippocampal gamma oscillations in vitro and in vivo.56,86,98,112 This finding underlines the general principle of sparse coding by pyramidal cells, which has been suggested to provide an optimal trade-off between information processing and energy efficacy.113,114 However, pyramidal cells comprise ~85% of all neurons in the hippocampus proper, and even with sparse coding they are able to encode multiple representations of spatial and episodic memories.18,20,23,85 By propagating specific activity patterns into downstream cortical and subcortical regions, pyramidal cells also impact on network activity and information processing in multiple other brain regions.9,10,21 From a neuronal network perspective, it can be said that gamma oscillations are characterized by high action potential frequencies, strongly divergent local connectivity and high synaptic efficacy of fast-spiking, PV+ interneurons. This contrasts with the far higher number but lower spiking frequency and efficacy of principal cells, such that excitation and inhibition are in balance.16,18 This balance is dynamically regulated and maintained by the feedback connectivity between pyramidal cells and PV+ basket cells.17,115,116
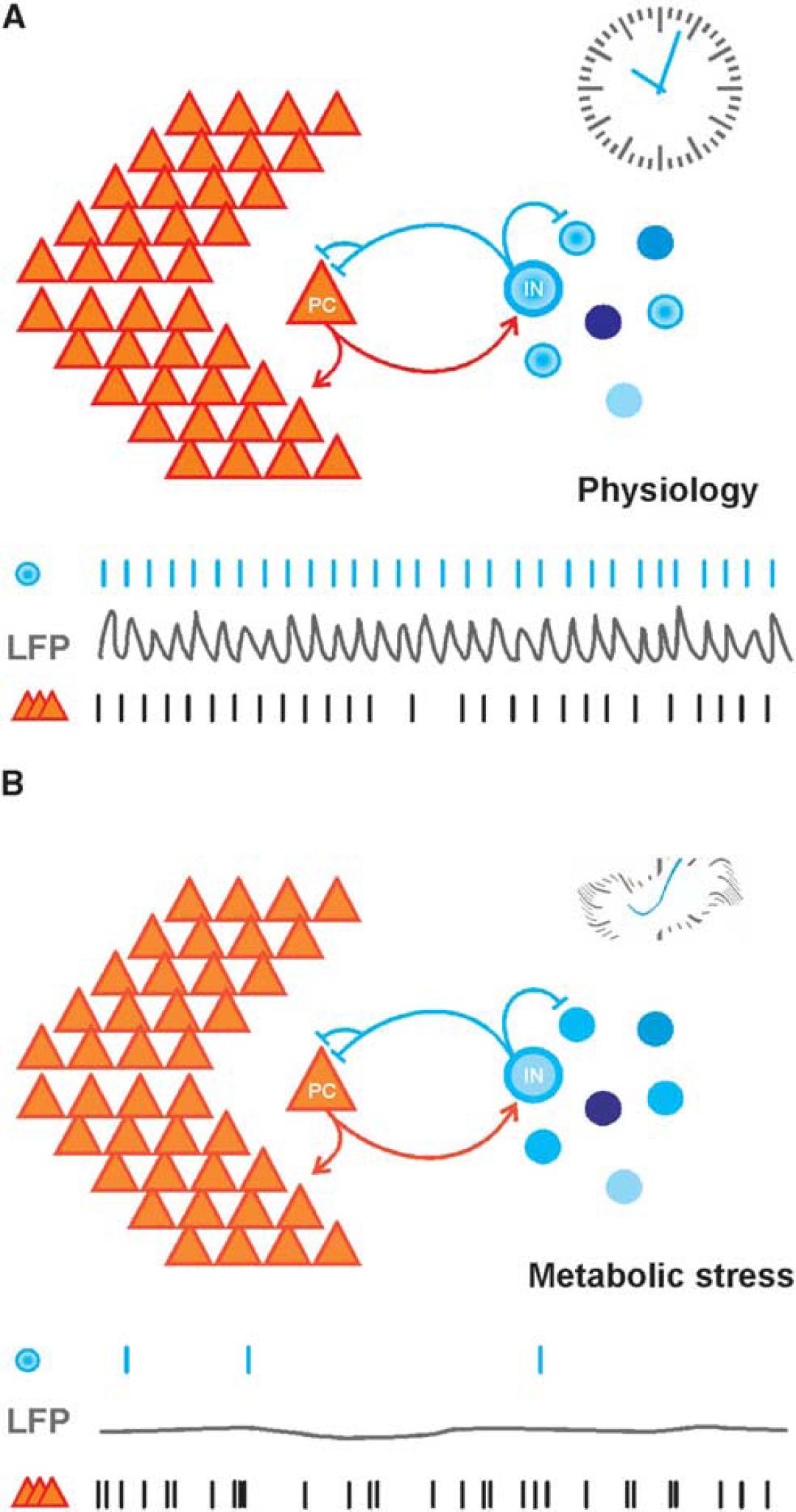
Highly energized inhibitory interneurons are a central element for information processing in cortical networks (interneuron energy hypothesis). (
In summary, convergent evidence from electrophysiological recordings, genetic interventions, quantitative neuroanatomical analysis, and mathematical modeling in both the hippocampus and neocortex reveals that fast-spiking, PV+ inhibitory interneurons are a central element for the generation of gamma oscillations and, therefore, information processing in the cortex.
MITOCHONDRIA IN FAST-SPIKING INHIBITORY INTERNEURONS
While this key role of fast-spiking, PV+ interneurons for fast rhythmic network activity has been consistently recognized, the underlying neuroenergetics have gained much less attention. It is likely, however, that patterns of neuronal network activity represent a compromise between maximal information processing and minimal energy utilization.113,114 It is therefore important to understand the activity-dependent neuroenergetic demands and constraints of this highly active class of neurons. Indeed, several findings indicate that fast-spiking interneurons use much more energy than other cells in the central nervous system, which might render them particularly vulnerable to conditions of energy deficiency. Energy utilization is counterbalanced by generation of cellular energy carrier, ATP via cytosolic glycolysis and oxidative phosphorylation in mitochondria, the latter of which requiring oxygen as a final electron acceptor.2,72 Indeed, most basket cells and axo-axonic cells contain considerably higher numbers of mitochondria in dendrites, somata and presynaptic terminals as compared with other interneuron subpopulations and pyramidal cells.74,117–123 This property is highly conserved in various species of mammals.74,117,124 Moreover, mitochondria of fast-spiking, PV+ interneurons are enriched with proteins such as cytochrome c oxidase (complex IV) and cytochrome c that are crucial for electron transport in the mitochondrial respiratory chain.74,121 Similar observations have been made in interneuron subpopulations of the visual cortex.124,125 We note that these findings indicate relative metabolic capacity rather than actual activity of fast-spiking interneurons in vivo. However, as there is evidence for a positive correlation between the capacity for oxidative metabolism and the rates of metabolic activity,74,126–128 the findings suggest that fast-spiking interneurons do indeed use high amounts of energy, which they gain from oxidative phosphorylation in mitochondria. Interestingly, several studies indicate that GABAergic interneurons, including PV+ interneurons participate in regulating hemodynamic responses.129–131 Moreover, the oxygen overshoot during the activity-dependent hemodynamic response might serve the upkeep of sufficient oxygen levels in mitochondria of fast-spiking interneurons, particularly when located in some distance from the vasculature.132,133
Why would fast-spiking interneurons need higher amounts of energy than more scarcely spiking principal cells or other types of interneurons? Two distinguishing features of fast-spiking interneurons have been suggested as activity-dependent mechanisms of high energy utilization: (i) high frequency and fast kinetics of the action potential and (ii) synaptic inhibition per se.18,134,135 While there is no final consensus on the reasons for the specific neuroenergetic profile of fast-spiking interneurons, several arguments indicate that both distinguishing features contribute to the high energy utilization and thus oxygen consumption.
ENERGY UTILIZATION OF FAST-SPIKING BEHAVIOR AND SYNAPTIC INHIBITION
As indicated by their name, fast-spiking interneurons can generate action potentials at high frequencies (> 100 Hz) for prolonged periods of time, with weak or no accommodation.37,85 Several properties of these neurons support this high level of activity: (i) the ‘resting’ membrane potential is ~10 to 15 mV closer to spike threshold than in pyramidal cells; (ii) intrinsic membrane properties include resonance at frequencies within the gamma-frequency band; (iii) action potentials follow a particularly rapid time course and propagate with high reliability along the largely unmyelinated axon.37,48,102,134,136–138 Notably, the conduction velocity of fast-spiking interneurons is higher than that of unmyelinated axons of glutamatergic principal cells. 138 These features of the action potential, however, come at a price: fast-spiking interneurons show (i) a ‘supercritical’ density of voltage-gated Na+ -channels all over the unmyelinated axon that results in high Na+-conductance density and (ii) a larger overlap of depolarizing Na+ -entry and repolarizing K+-efflux (‘sodium entry ratio’ of ~1.98), which differs markedly from cortical principal cells.135,138,139 The ‘supercritical’ density of Na+-channels is thought to compensate for the morphologic axonal properties of fast-spiking interneurons, i.e., small segmental diameter, extensive branching and high bouton density and to permit fast propagation of the action potential. 138 Thus, high-frequency repetitive firing in fast-spiking interneurons likely causes a much larger dissipation of ion gradients than in neurons with energetically optimized action potential waveforms.138,139 This would enhance the need for ATP to fuel Na+/K+-ATPase and Ca2+ -ATPase to restore the original, non-equilibrium ion distributions. However, energy-saving mechanisms that may partially compensate for the high ATP demand have been discussed for fast-spiking interneurons, i.e., fewer excitatory input sites, partially higher input resistance as well as expression of voltage-gated K+-channels of the Kv3 subtype and rapidly inactivating Na+-channels.18,134,138,140–143 Examples from the basal ganglia also suggest that high spiking rates of GABAergic neurons do not necessarily correlate with their energetic needs. 18 As long as we are lacking direct empirical evidence for the activity-dependent energy utilization of these interneurons, it will be difficult to decide how much energy is required to permit their fast-spiking behavior. Systematic recordings from dendrites and axons under physiologic and pathophysiological conditions are required to precisely assess the energy costs of fast-spiking interneurons.
As an alternative of complementary explanation for the high energy utilization the synaptic functions of fast-spiking GABAergic interneurons need to be considered. During fast network oscillations, almost all local neurons are phasically inhibited on every oscillation cycle, and most of this inhibition is perisomatic, i.e., at the soma, the proximal dendrites and the axon initial segment.35,52,87,107 In neuronal networks of the hippocampus proper, fast-spiking PV+ interneurons form numerous GABAergic synaptic contacts to pyramidal cells in the perisomatic region23,87 (Figures 1C–E). As outlined above, these synapses are strongly and rhythmically activated during fast network oscillations, and multiple lines of evidence indicate that this phasic inhibition is a key factor in coordinating neuronal activity during network states such as gamma oscillations or sharp wave-ripple complexes.9,10 It would, therefore, be desirable to precisely know the energy utilization as well as the frequency of unitary GABAergic events during resting states and gamma oscillations. This would allow for a solid quantification of the inhibition-related energy utilization of fast-spiking interneurons, which includes the presynaptic GABA release as well as the postsynaptic GABA effects in both principal cells and interneurons. However, this turns out to be a difficult task. 18 Fast phasic GABAergic inhibition acts via activation of postsynaptic anion channels, i.e., GABAA receptors.16,37 These channels are selectively permeable to chloride and, to a lower extent, bicarbonate (HCO3−), thereby shifting the postsynaptic membrane potential more closely the reversal potential of these ions.144,145 These processes contain several energy-utilizing steps for release and metabolism of GABA (see below) and, most importantly, the restoration of membrane ion gradients. Transmitter release is a very fast and highly specialized process that involves the presynaptic vesicular cycle, Ca2+-dependent membrane fusion and subsequent buffering or extrusion of calcium from the presynaptic terminal.146,147 In principle, the energy costs of these processes should not differ between excitatory and inhibitory synapses, but the frequency of release is likely much higher in terminals of fast-spiking interneurons as compared with other cell types. Fast-spiking interneurons have by far the highest frequency of action potentials during gamma oscillations.23,52,85,88 At the same time, the release of GABAergic synaptic vesicles from fast-spiking interneurons shows large quantal size, high release probability and, presumably, less conduction failures as compared with typical excitatory neurons.4,37,100,148,149 Moreover, an activity-dependent gating mechanism that reduces release probability appears to preserve the releasable pool of synaptic vesicles during high-frequency repetitive firing. 100 While a quantification of energy costs is difficult in the absence of direct experimental data, a recent study in the hippocampal slices does indeed suggest that presynaptic Ca2+-removal, vesicle turnover, and neurotransmitter transport use more energy than generally assumed. 150 Thus, it is well feasible that the sustained and fast cycling of vesicles as well as the very frequent Ca2+-entry in presynaptic terminals of fast-spiking interneurons require significant amounts of ATP, in line with the high numbers of presynaptic mitochondria.74,117,121,123 A second energy-utilizing process in synaptic transmission is the restoration of ion gradients across neuronal membranes. This process primarily affects the postsynaptic target neurons of inhibitory synapses. Indeed, the perisomatic region of pyramidal cells in CA3, which is the hippocampal subfield with most prominent gamma oscillations and oxygen consumption, shows alternating current sinks and sources during gamma oscillations in vitro and in vivo.22,56 Moreover, this perisomatic region contains numerous mitochondria, which is not a general feature of cortical principal cells.70,74,151 In case of inhibitory synapses with dominant Cl−-conductance, restoration of ion gradients is served by secondarily active transporters, especially the K+/C;−-cotransporter, KCC2.16–18,144 When chloride enters the cell during synaptic inhibition, subsequent Cl−-transport will be directed outwardly, against the equilibrium potential of chloride. The required driving force for the outward transport is provided by the electrochemical potential of potassium being established by the Na+/K+-ATPase. However, it is currently not clear how much Cl−-flux underlies synaptic inhibition during different activity states in cortical networks. 18 Consider the case where resting membrane potential and equilibrium potential of chloride are identical: in this situation, there is no net flux of chloride, despite an increased membrane conductance because of the open GABAA receptors. Such ‘shunting inhibition’ can still be highly efficient to reduce the effect of glutamatergic inputs,16,17,145 without changing the distribution of chloride. In addition to hyperpolarizing inhibition, shunting inhibition seems to be indeed a typical case rather than an exception in cortical principal cells.16,152,153 As a general rule, resting membrane potential of pyramidal cells and equilibrium potential of chloride are not very far from each other, which perhaps constitutes an energy-saving mechanism in networks with intense synaptic inhibition. 18 Fast-spiking interneurons, in contrast, have ~10 to 15 mV more depolarized membrane potentials than pyramidal cells37,102 and may thus have considerably higher energy costs for Cl−-extrusion associated with mutual GABAergic inhibition. There might be yet another challenge to ion homeostasis even when the equilibrium potential of chloride is not far from the resting membrane potential: excitatory and inhibitory synaptic inputs do usually form a dynamic equilibrium, i.e., an increase of inhibition goes along with an increase of excitation and vice versa. 54 This increase in overall synaptic activity keeps excitability within a narrow functional range; but in situations of high membrane conductance, it causes increased ion fluxes for excitatory transmission, even when inhibition is purely shunting. 18 Thus, ion homeostasis during network activity states with strong synaptic inhibition may create a considerable energy demand, in both inhibitory interneurons and excitatory pyramidal cells.
A related important aspect is neurotransmitter metabolism, i.e., biosynthesis, vesicular storage, and release of GABA in inhibitory interneurons. GABA is derived from glutamate by glutamic acid decarboxylase, which occurs in two isoforms, GAD65 and GAD67. While GAD67 is expressed throughout the cytosol, GAD65 is the dominant presynaptic isoform, which is directly associated to presynaptic vesicles and therefore seems to be important for their filling.106,155,156 This filling process, however, requires activity of a vacuolar H+-ATPase that provides the electrical (Δψ) and chemical (ΔpH) components of the driving force for the vesicular inhibitory amino-acid transporter.147,157 After release to the synaptic cleft, re-uptake of GABA occurs via Na+-/Cl−-dependent GABA transporter 1 in the presynaptic terminal or via GAT-3 in processes of astrocytes that enwrap the synapse. Again, these processes are secondarily active, thus utilizing potential energy that has been previously provided by the Na+/K+-ATPase and other transport systems. 147 However, for mouse astrocytes in primary culture, it was described that GABA uptake resulted in increases of cytosolic Na+-concentration but not of glucose utilization. 158 Systematic studies in complex brain preparations during naturally occurring network states are required to further elucidate the energy costs of astrocytic GABA uptake. In astrocytes, GABA is subsequently converted to glutamine by several enzymatic steps. Glutamine is finally shuttled from astrocytes to neurons by secondarily active transporters (glutamate/GABA-glutamine cycle), before being converted to glutamate via phosphate-activated glutaminase.106,159 It is important to note that all involved membrane transport processes depend directly or indirectly on ATP. They may, therefore, significantly contribute to the energetic needs of enhanced synaptic inhibition. Mitochondria of interneurons are also involved in the conversion of α-ketoglutarate or glutamine to glutamate while conversion of GABA to glutamate takes place in astrocytic mitochondria.106,159 Notably, several studies suggest that the mitochondrial tricarboxylic acid cycle is central for the biosynthesis of GABA from glutamine or lactate, an alternative source of the transmitter.155,159–162
In summary, electrophysiological recordings, quantitative neuroanatomical and biochemical analyses in different cortical preparations suggest that fast-spiking interneurons have unique electrophysiological and neuroenergetical properties that ultimately require sufficient substrate and oxygen supply as well as proper mitochondrial performance for generation of ATP and allocation of GABA.
PERSPECTIVE: METABOLIC AND OXIDATIVE STRESS IN FAST-SPIKING INTERNEURONS
The rapid abolishment in the power of gamma oscillations has been described under various experimental conditions in slice preparations: (i) when the pO2 of the ambient atmosphere was lowered to the normoxic range under semi-interface recording condition, 44 (ii) when the flow rate of oxygenated recording solution was below a critical limit under submerged recording condition, 47 and (iii) when hypoxic events were induced. 46 In line with these findings, unilateral hippocampal ischemia in vivo caused a transient of pathologic high-frequency discharges followed by a dramatic and long-lasting selective decrease in the power of the gamma oscillations. 163 Such decreases in gamma oscillation power were also observed in vitro during pharmacological interference with mitochondrial function (‘poisoning’), namely inhibition of the mitochondrial respiratory chain by rotenone (acting on complex I) or potassium cyanide (acting on cytochrome c oxidase in complex IV), or after mitochondrial uncoupling by protonophores.41,164,165 Intriguingly, other forms of activity such as electrical stimulus-evoked neuronal activation, low-frequency oscillations, and seizure-like events were much more resistant to low pO2 or pharmacological inhibition of the mitochondrial respiratory chain in vitro and in vivo,41,44,163 suggesting that fast-spiking interneurons that are crucial for the gamma oscillations were primarily affected.
The studies summarized above show that fast-spiking interneurons are capable to maintain cellular energy homeostasis during gamma oscillations under physiologic conditions. By contrast, fast-spiking interneurons may dramatically lower the threshold for neuronal network dysfunction during metabolic stress that occurs during hypoxia, ischemia, or dysfunction of mitochondria. This notion is indirectly supported by unique morphologic, electrophysiological, and neuroenergetical properties of fast-spiking interneurons as well as by their central role in the generation of network oscillations (see above). A recent study provided the first direct experimental insight into the cellular mechanisms underlying the rapid decrease of gamma oscillations during metabolic stress. 164 The authors found no significant changes in resting membrane potential of pyramidal cells during application of potassium cyanide (acting on cytochrome c oxidase in complex IV), indicating maintenance of cellular excitability. By contrast, fast-spiking basket cells showed a sustained membrane depolarization, resulting in almost complete cessation of spiking and thus strongly reduced rates of IPSPs impinging on pyramidal cells. This rapid dysfunction does not necessarily imply a lower threshold of PV+ interneurons to ischemia-induced neuronal death. 166 Similar findings were obtained for fast-spiking interneurons during gamma oscillations when oxygen supply was lowered. 47 This first experimental evidence suggests that even moderate hypoxia or poisoning of mitochondrial oxidative phosphorylation selectively results in rapid decrease in ATP levels and GABA release in fast-spiking interneurons.41,167 This might be of particular importance for those fast-spiking interneurons that are located in the periphery of cortical vascular supply regions, with limited oxygen availability. 133 Moderate metabolic stress may thus disrupt fast phasic inhibition of fast-spiking interneurons more effectively than other types of inhibition or pyramidal cell excitation in neuronal networks87,116 (Figure 2B). The disruption of rhythmic inhibition would cause rapid loss of gamma oscillations, with devastating consequences on information processing in local and, perhaps, remote cortical networks34,41,44,164 (Figure 3). This pathophysiological mechanism might help to explain the exceptional vulnerability of higher brain functions during acute metabolic stress, which has long been known from clinical medicine and experimental neurology. 53 In particular, symptoms such as visual field narrowing and unconsciousness being accompanied with slow-wave activity in the electroencephalogram occur very rapidly during ischemia in both humans and animals, whereas evoked responses and ion distributions are more resistant.168–171
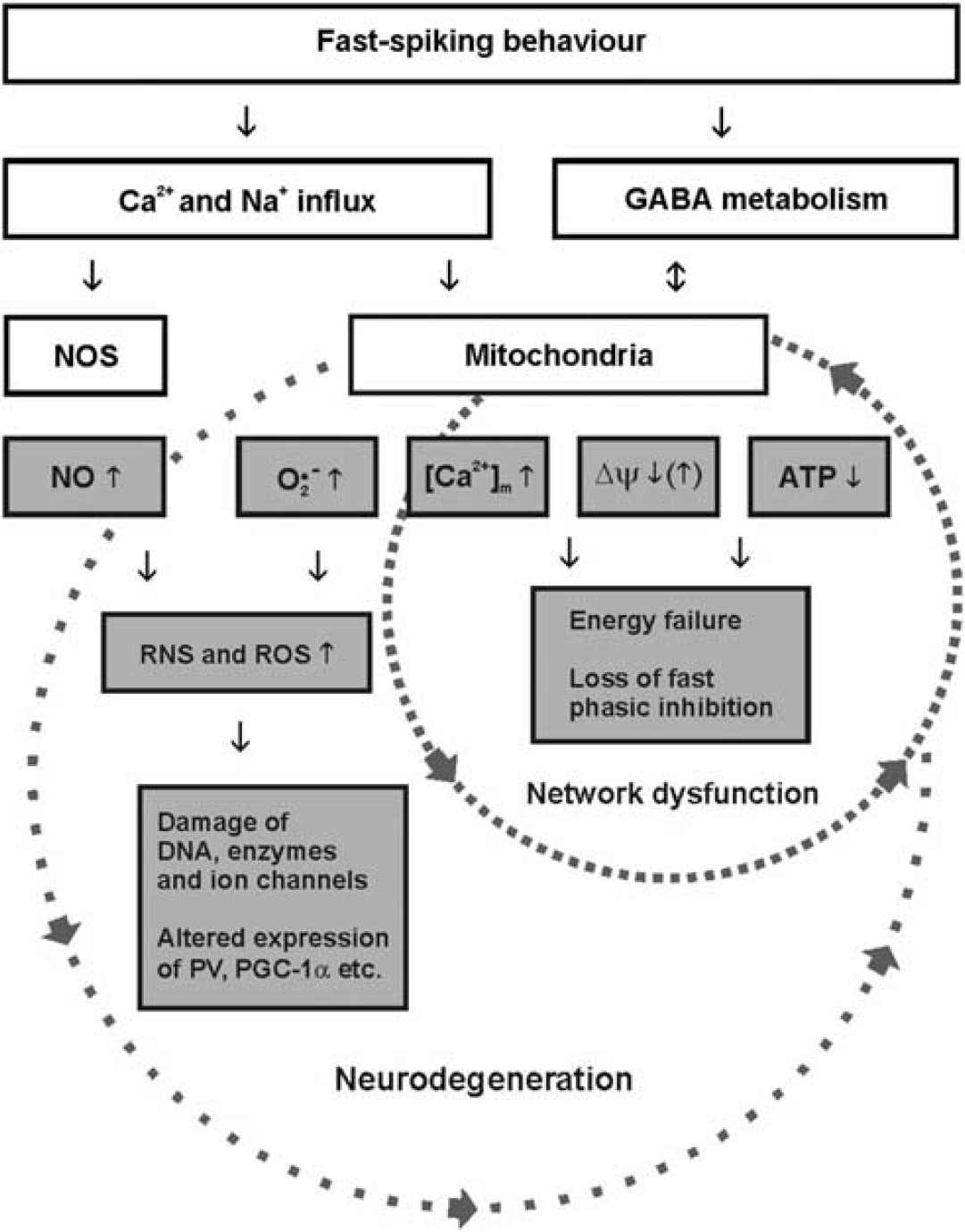
The simplified scheme illustrates the perspective on metabolic and oxidative stress in fast-spiking, parvalbumin-positive interneurons. The small circle reflects acute effects of hypoxia/ischemia or mitochondrial poisoning that rapidly result in neuronal network dysfunction. The large circle reflects chronic effects of mitochondrial dysfunction and/or cerebral hypoperfusion that finally result in neurodegeneration. Note that the pathophysiological mechanisms illustrated in both circles may partially interact, depending on the course of the disease and pathogenic factors such as alterations in expression of ion channels, parvalbumin or PGC-1α that may result in imbalances of oxidative and anti-oxidative processes. [Ca2+]m, mitochondrial Ca2+-concentration; GABA, γ-aminobutyric acid; NO, nitric oxide; NOS, nitric oxide synthase; O2•−, superoxide anion; Δψ, mitochondrial membrane potential; RNS, reactive nitrogen species; ROS, reactive oxygen species; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-α PV, parvalbumin.
Fast-spiking interneurons might also experience higher levels of oxidative stress under both physiologic and pathophysiological conditions. At the mitochondrial respiratory chain, the small amount of electron leakage (0.1% to 4%) induces one-electron reduction of oxygen, which results in relatively stable superoxide anion—a free radical with biologic toxicity.
172
Superoxide anion is generated at several mitochondrial sites such as complexes I and III, and this occurs readily during mitochondrial respiration at physiologic levels of neuronal activity. Specific genome mutations, for example, alter the electron transfer properties of respiratory chain complexes and result in abnormal high levels of superoxide anion.
173
Such abnormal levels can favor the accumulation of hydrogen peroxide and the appearance of other molecular oxygen-derived free radicals and precursors, collectively known as reactive oxygen species (ROS).172,173 Another gaseous molecule is nitric oxide (NO) that is generated through enzymatic conversion of
These putative pathophysiological mechanisms in fast-spiking interneurons, i.e., higher susceptibility to metabolic and oxidative stress, might apply to several neurologic and psychiatric disorders that go along with chronic mitochondrial dysfunction and/or chronic cerebral hypoperfusion (Figure 3). Clinical examples are mitochondrial disorders (mitochondriopathies), arteriosclerosis, ageing, Alzheimer's disease, Parkinson's disease, and schizophrenia.5,13,72,167,172,185 Indeed, experimental evidence suggests that fast-spiking interneurons have a key role in the (patho)physiologic processes underlying ageing, Alzheimer's disease, and schizophrenia.165,183,186–188 While recent studies yielded considerable insight into mitochondria and neuroenergetics during gamma oscillations, important features of interneurons, in particular, of fast-spiking interneurons, have been less defined. This includes detailed information about electrophysiological properties, bioenergetics, energy efficacy, GABA metabolism, free radicals, microdomains with densely packed respiring mitochondria, as well as supply of oxygen and energy substrates distant from the vasculature. These aspects need to be studied under physiologic and pathophysiological conditions in adult and aged animals. Therefore, further understanding of neuronal network dysfunction as related to metabolic and oxidative stress requires combined efforts from molecular genetics, biochemistry, electrophysiology, and live-cell imaging techniques with high spatial and temporal resolution, including mathematical modeling.
Footnotes
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
The authors thank Dr Nikolaus Berndt for critical reading of the manuscript and Andrea Lewen for text editing assistance.