
Abstract
Activation of the basal forebrain (BF), the primary source of acetylcholine (ACh) in the cortex, broadly increases cortical cerebral blood flow (CBF), a response downstream to ACh release. Although endothelial nitric oxide and cholinoceptive GABA (γ-aminobutyric acid) interneurons have been implicated, little is known about the role of pyramidal cells in this response and their possible interaction with astrocytes. Using c-Fos immunohistochemistry as a marker of neuronal activation and laser-Doppler flowmetry, we measured changes in CBF evoked by BF stimulation following pharmacological blockade of c-Fos-identified excitatory pathways, astroglial metabolism, or vasoactive mediators. Pyramidal cells including those that express cyclooxygenase-2 (COX-2) displayed c-Fos upregulation. Glutamate acting via NMDA, AMPA, and mGlu receptors was involved in the evoked CBF response, NMDA receptors having the highest contribution (∼33%). In contrast, nonselective and selective COX-2 inhibition did not affect the evoked CBF response (+0.4% to 6.9%, ns). The metabolic gliotoxins fluorocitrate and fluoroacetate, the cytochrome P450 epoxygenase inhibitor MS-PPOH and the selective epoxyeicosatrienoic acids (EETs) antagonist 14,15-epoxyeicosa-5(Z)-enoic acid (14,15-EEZE) all blocked the evoked CBF response by ∼50%. Together, the data demonstrate that the hyperemic response to BF stimulation is largely mediated by glutamate released from activated pyramidal cells and by vasoactive EETs, likely originating from activated astrocytes.
Introduction
Stimulus-induced changes in cerebral blood flow (CBF) underlie the basis of functional brain imaging techniques such as positron emission tomography and functional magnetic resonance imaging that use hemodynamic signals to map changes in neuronal activity under physiological and pathological conditions. Although the cellular and molecular bases of this tight coupling are not yet fully elucidated, evidence suggests that the hemodynamic changes are driven by the incoming afferents and their local processing within the activated area (Lauritzen and Gold, 2003). Additional
Stimulation of the basal forebrain (BF), the primary cholinergic input to the neocortex arising from the substantia innominata-nucleus basalis complex, produces large increases in ipsilateral cortical CBF in the rat concomitant with the local release of acetylcholine (ACh) (for review, see Hamel, 2004). Cholinergic basalocortical afferents have a widerange of targets including excitatory pyramidal cells (Henny and Jones, 2008; Houser et al, 1985), inhibitory interneurons (Cauli et al, 2004), microvessels, and astrocytes (Vaucher and Hamel, 1995). In accordance with these anatomical findings, part of the CBF response elicited by BF stimulation has been attributed to a direct vasodilatory effect of ACh through muscarinic receptor (mAChR)-mediated endothelial, but not neuronal, NO (nitric oxide) release (Zhang et al, 1995). We accordingly found that the hemodynamic response to BF stimulation was virtually eliminated by selective cortical cholinergic deafferentation despite preserved NO synthase containing BF neurons and cortical projections. Further, we showed that the full expression of the CBF response requires GABA-A (γ-aminobutyric acid) receptor activation by specific subsets of cholinoceptive GABA interneurons that contain SOM (somatostatin) and NPY (neuropeptide Y) (Kocharyan et al, 2008). These findings correlated very well with the synchronized bursting induced in these GABA interneurons by mAChR activation (Beierlein et al, 2000), and with the cortical hemodynamic signals induced by γ oscillations—as elicited by activation of BF cholinergic afferents (Cape et al, 2000)—being initiated by the firing of inhibitory interneurons (Niessing et al, 2005).
Pyramidal cells are key players in the neurovascular coupling response to sensory stimulation that is driven by glutamatergic thalamocortical afferents. Pyramidal cells act mainly through the release of cyclooxygenase-2 (COX-2)-derived vasoactive prostanoids (Lecrux et al, 2011; Niwa et al, 2000) and that of glutamate. The latter is thought to induce calcium transients in astrocytes (Zonta et al, 2003), which results in the synthesis and release of potent vasoactive derivatives such as the epoxyeicosatrienoic acids (EETs) (Koehler et al, 2009). Pyramidal cells are also activated following stimulation of cholinergic basalocortical afferents (Kocharyan et al, 2008), but nothing is known regarding their role or that of glutamate or COX-2 products, or about the interplay between pyramidal cells, interneurons, and astrocyte-derived vasoactive messengers in this hyperemic response. In this study, we sought to identify the contribution of these cell types in the CBF response evoked by BF stimulation. Our results demonstrate that glutamate-releasing pyramidal cells and AA (arachidonic acid)-derived cytochrome P450 epoxygenase EETs, but not COX-1 or COX-2 products, mediate the bulk of this neurovascular coupling response.
Materials and methods
Procedures for BF stimulation, CBF measurements, and immunohistochemistry were similar to those detailed previously (Kocharyan et al, 2008; Lecrux et al, 2011). All experiments were approved by the McGill University animal ethics committee and conformed to the Canadian Council on Animal Care.
Animals, Surgical Procedures, and Basal Forebrain Stimulation
The experimental procedure was in two steps: first, chronic implantation of either an electrode or a guide cannula in the BF, and second, 1 week later, electrical or chemical BF stimulation for pharmacological or immunohistochemical studies. Male Sprague-Dawley rats (280 to 300 g; Charles River, Montréal, QC, Canada) were anesthetized with ketamine-xylazine (100/5 mg/kg, intraperitoneally) or isoflurane (2% in a 40%/60% oxygen/air mixture via a face mask) and placed in a stereotaxic frame (model 962; D Kopf Instruments, Tujunga, CA, USA) for chronic implantation of monopolar tungsten electrodes (0.35 mm outer diameter; FHC, Bowdoin, ME, USA) or guide cannulas (Plastics One, Roanoke, VA, USA) in the left substantia innominata (AP: −1.2 mm, lateral: +2.4 mm, ventral: −6.9 mm (electrodes) or −5.3 mm (guide cannulas) from Bregma). Body temperature was maintained at 37°C using a heating pad. One week later, BF electrical stimulation was performed under urethane anesthesia (1.1 g/kg, intraperitoneally) using parameters previously determined as optimal for immunohistochemistry (100 Hz, 80 μA, 0.5 milliseconds, 1 second on/1 second off for 5 minutes) or pharmacological experiments (100 Hz, 50 μA, 0.5 milliseconds, 1 second on/1 second off for 20 seconds) (isolated pulse stimulator; A-M Systems, Sequim, WA, USA) (Kocharyan et al, 2008). During pharmacological studies, a catheter was inserted in the femoral artery for blood pressure measurement (AD Instruments, Colorado Springs, CO, USA) and blood gas analysis (pH, pO2, and pCO2) (Bayer Rapid Lab 348, Siemens Healthcare Diagnostics Inc., Deerfield, IL, USA). All remained within normal range throughout the experiments (see Supplementary Table S1), which lasted at most 2.5 hours. For immunohistochemical studies, we additionally performed chemical BF stimulation by injecting 0.1 μL of sodium glutamate (100 nmol,
Pharmacological Experiments: Cerebral Blood Flow Measurements by Laser-Doppler Flowmetry
Cerebral blood flow increases induced by BF stimulation were measured by laser-Doppler flowmetry (Transonic Systems, Ithaca, NY, USA) with two needle-shaped probes positioned on the thinned bone overlying the ipsilateral and contralateral frontal cortices (∼4 mm2 anterior from the Bregma). Intracisternal injection (ic) of drugs or vehicles was performed under microscope monitoring using a 30-gauge needle connected by a polyethylene-10 catheter to a Hamilton microsyringe (3 μL over 1 minute; Harvard Apparatus infusion pump) (Figure 1A). After 15 to 20 minutes of CBF stabilization, CBF changes induced by a 20-second BF stimulation were measured at baseline and following vehicle or drug injection. Time courses of drug effects were determined, and the evoked CBF changes were taken at the time of maximal effect. Unless indicated otherwise, these times corresponded to 20 minutes after receptor antagonism or 40 to 50 minutes after enzyme inhibition. Each rat received only one compound or combination of compounds, and the corresponding vehicle. Diffusion of the ic-injected drugs over the cortex was validated on the evoked CBF response to whisker stimulation by the identical reducing effects of picrotoxin after ic injection or cortical superfusion through a closed cranial window (Lecrux et al, 2011). It was further confirmed in the present study in urethane-anesthetized rats injected with saline or 3% Evans blue (
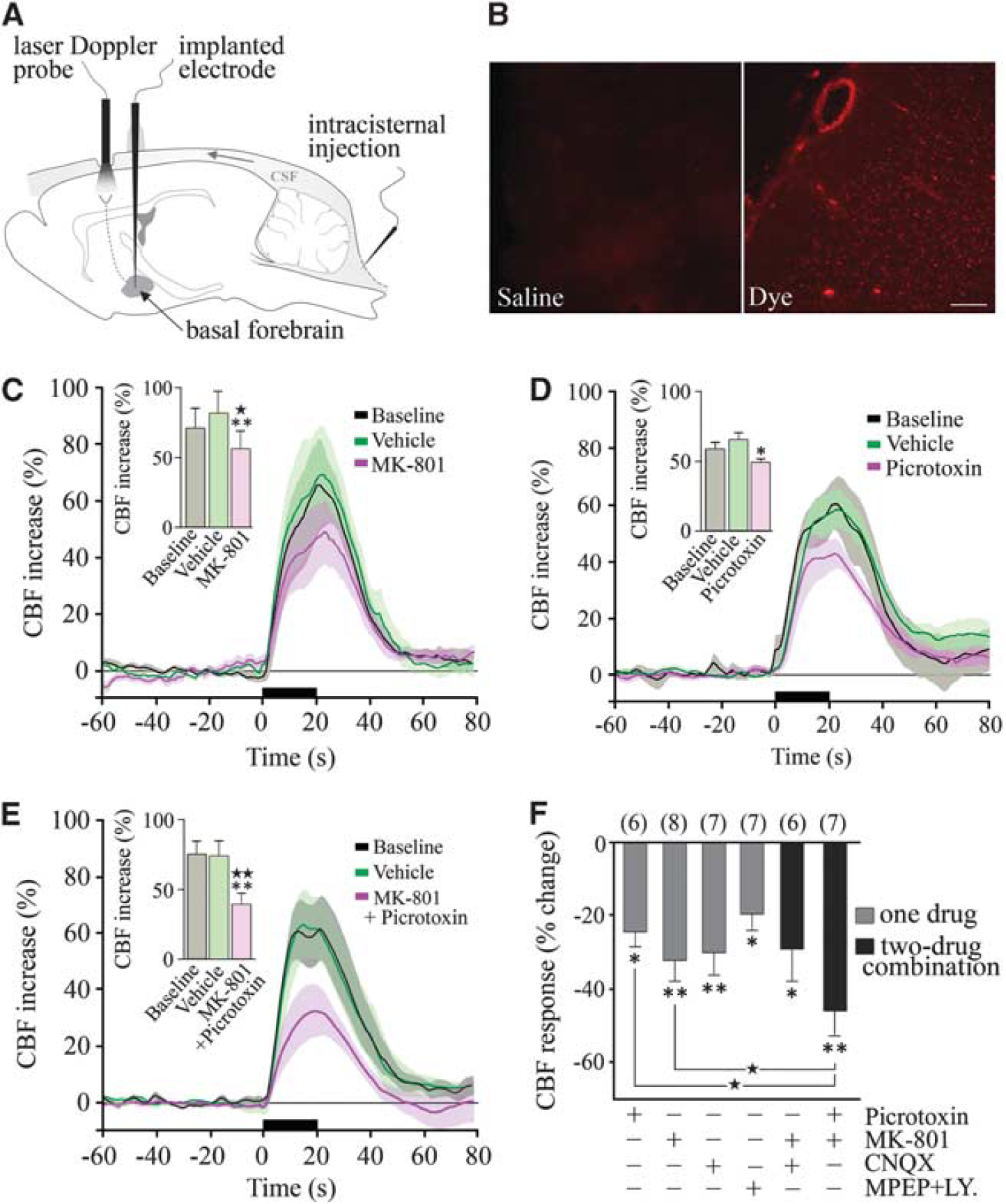
Glutamate and GABA (γ-aminobutyric acid) in the cerebral blood flow (CBF) response to basal forebrain (BF) stimulation. (
It is well documented that it is impossible to completely block CBF responses induced by increased neuronal activity even with high doses of receptor antagonists or enzyme inhibitors, or through combined blockade of several enzyme activities or receptors (Koehler et al, 2009; Leithner et al, 2010), likely because of overlap and interactions between several effector pathways. Therefore, we used the
Drug Preparation
Unless otherwise stated, 3 μL of a 10−4 mol/L buffered solution (pH 7.4) of each compound or combination of compounds, or their corresponding vehicle were injected. 6-Cyano-7-nitroquinoxaline-2,3(1H4H)-dione disodium salt (CNQX; vehicle: 0.5 mol/L phosphate-buffered saline (PBS)), MK-801 (vehicle: 0.5 mol/L PBS), and picrotoxin (vehicle: 0.5% ethanol in 0.5 mol/L PBS) were purchased from Tocris Biosciences (Minneapolis, MN, USA). Fluoroacetate sodium (vehicle: 0.5 mol/L PBS), fluorocitrate (vehicle: 0.5 mol/L PBS, 3 × 10−4 mol/L), indomethacin (vehicle: 0.2% ethanol in 0.5 mol/L PBS), MPEP (6-methyl-2-(phenylethynyl)pyridine; vehicle: 0.5 mol/L PBS), LY-367385 (vehicle: 10−3 mol/L NaOH in 0.5 mol/L PBS equilibrated to pH 7.4 with 1 N HCl), and scopolamine (vehicle: 0.5 mol/L PBS, 1 mg/kg, intravenous) were purchased from Sigma-Aldrich Canada Ltd (Oakville, ON, Canada). 14, 15-Epoxyeicosa-5(Z)-enoic acid (14,15-EEZE; vehicle: 0.25% ethanol in 0.5 mol/L PBS),
Immunochemical Staining for Pyramidal Cells Recruited by Basal Forebrain Stimulation
One hour after stimulation of the left BF, rat brains were fixed by intraaortic perfusion (500 mL of 4% paraformaldehyde in 0.1 mol/L phosphate buffer), postfixed by immersion (2 hours, 4°C), cryoprotected, frozen (–45°C in isopentane) and sectioned as free-floating coronal sections (25 μm thick) on a freezing microtome. Activated pyramidal cells were detected by double-immunocytochemistry for c-Fos (rabbit anti-c-Fos, 1:15,000; Oncogene, San Diego, CA, USA) and a ubiquitous pyramidal cell marker (mouse anti-rat brain pyramidal cells (RBPC), 1:2,000; SWANT, Bellinzona, Switzerland) or COX-2 (mouse anti-COX-2, 1:3,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), which is constitutively expressed by a subset of cortical pyramidal cells (Kaufmann et al, 1996). Rat brain pyramidal cell and COX-2 were detected in the first position with an anti-mouse secondary antibody (1:200; Vector Labs, Burlingame, CA, USA) followed by the ABC complex (Vectastain ABC kit; Vector Labs) and 3,3-diaminobenzidine (Vector Labs; brown precipitate), and c-Fos was detected in the second position with a biotinylated secondary antibody and the SG reagent (Vector Labs; blue—gray precipitate) (Kocharyan et al, 2008). Sections were observed under light microscopy and digital pictures taken, calibrated, and edited with MetaMorph 6.1r3 (Universal Imaging, Downington, PA, USA) and Adobe Photoshop 7 (Adobe Systems, San Jose, CA, USA).
Quantitative Analysis
Laser-Doppler flowmetry values were measured as arbitrary units, and CBF changes induced by BF stimulation expressed as percent changes of the peak CBF value compared with 1 minute average baseline. For figure representation, CBF was averaged every 1 second starting 1 minute before until 1 minute after the stimulation, and expressed as percent change compared with the 1-minute average prestimulus baseline. Physiological parameters and changes in peak CBF were compared by repeated-measures ANOVA (analysis of variance) or by one-way ANOVA for three group comparisons followed by
Results
Cortical Cerebral Blood Flow Effects of Basal Forebrain Electrical Stimulation
We first confirmed that electrical BF stimulation induced a substantial increase in ipsilateral cortical CBF compared with the contralateral side (65.5% ± 2.9% versus 21.8% ± 1.4%,
Immunohistochemical Identification of Activated Pyramidal Cells
Pyramidal cells being primarily accountable for the increased cortical activity and hemodynamic response to sensory stimuli (Koehler et al, 2009; Lecrux et al, 2011; Norup Nielsen and Lauritzen, 2001; Zonta et al, 2003), we investigated their contribution in the hyperemic response to incoming basalocortical afferents. We used c-Fos as a marker for increased neuronal activity as it is well validated in the neocortex (Staiger et al, 2000) where it was previously found to selectively accumulate in SOM- and NPY-containing GABA interneurons in BF-stimulated rats (Kocharyan et al, 2008). In double-immunostained sections following either chemical or electrical BF stimulation, c-Fos protein levels were significantly upregulated in a large number of pyramidal cells immunopositive for the ubiquitous marker RBPC (Figure 2A, not quantified) and for COX-2, which is coexpressed in a subset of these neurons (Kaufmann et al, 1996). Except for layer V, RBPC immunostaining distributed throughout the different layers of the ipsilateral cortex, whereas COX-2 immunoreactive pyramidal cells were primarily found in layers II/IV (Figures 2B and 2C). A smaller proportion of COX-2 pyramidal cells were activated by electrical compared with chemical stimulation with NMDA or glutamate (Figure 2C). Quantitative analysis showed that NMDA, glutamate, or electrical BF stimulation induced c-Fos in 30.5% ± 7.9%, 38.9% ± 10.2%, and 27.1% ± 2.3% of COX-2 pyramidal cells in the ipsilateral cortex, compared with 8.0% ± 4.2%, 15.4% ± 3.1%, and 17.3% ± 0.5%, respectively, on the contralateral side (Figure 2C). These data indicate activation of various glutamate-releasing pyramidal neurons spanning several layers of the cortex targeted by BF afferents. Together with our previous findings of activated GABA interneurons (Kocharyan et al, 2008), these results demonstrate that both excitatory pyramidal cells and inhibitory interneurons are recruited by BF afferents, consistent with both cell types being targeted by basalocortical afferents (Cauli et al, 2004; Henny and Jones, 2008; Houser et al, 1985).
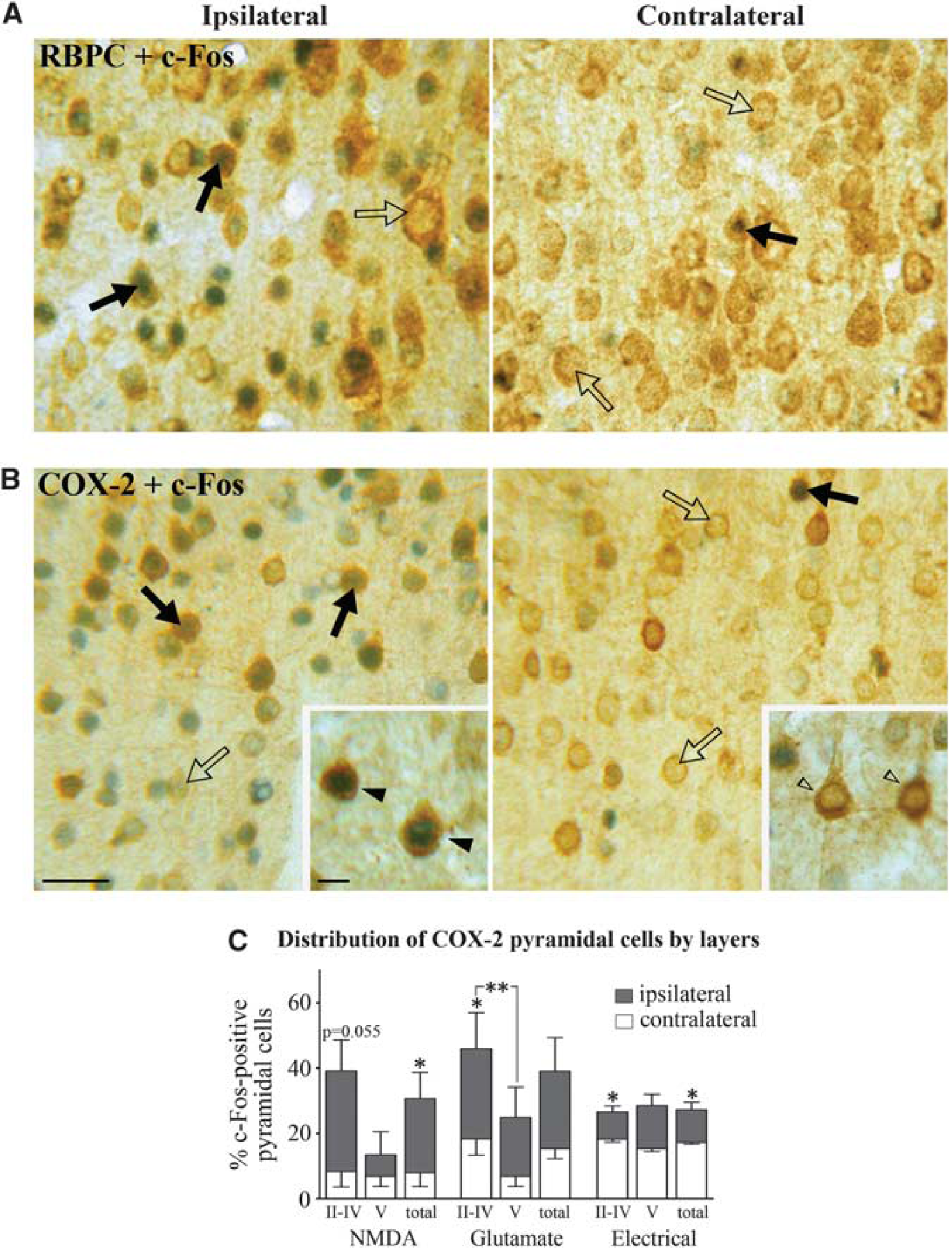
Basal forebrain (BF) stimulation activates cortical pyramidal cells. BF stimulation activated pyramidal cells predominantly in the ipsilateral cortex, as shown by upregulation of c-Fos protein (blue—gray) in the nuclei of RBPC (rat brain pyramidal cell) (
Glutamate in the Neurovascular Coupling Response to Basal Forebrain Stimulation
We then tested the possible implication of excitatory glutamate-releasing pyramidal cells in the CBF response to BF stimulation. We found significant decreases after selective antagonism of NMDA receptors with MK-801 (–32.2% ± 5.3%,
Activation of Glutamate- and γ-Aminobutyric Acid-Releasing Cells Is Downstream of mAChR Activation
Acetylcholine is the prime instigator of the hyperemic response to BF stimulation (Zhang et al, 1995) and the latter is almost completely abolished (–85%) by cortical cholinergic denervation with the selective cholinotoxin 192-IgG saporin (Kocharyan et al, 2008) and by combined blockade of muscarinic and nicotinic ACh receptors (Biesold et al, 1989). Here, we observed an important role for mAChRs with large decreases in the evoked CBF after scopolamine (–55.5% ± 4.2% at 40 minutes,
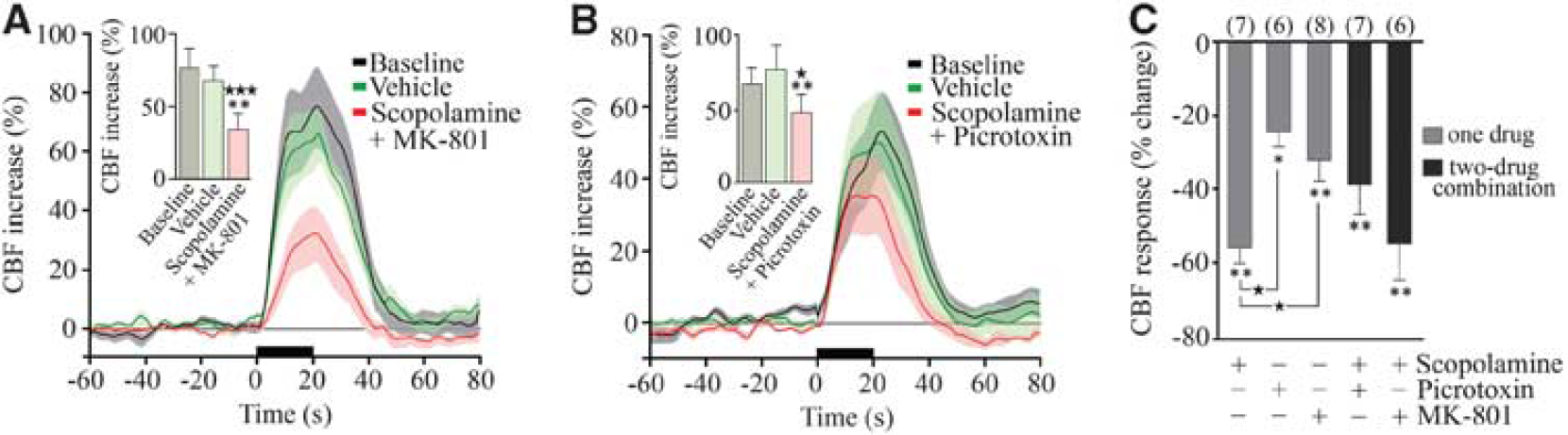
The reducing effects of NMDA (
Arachidonic Acid Products in the Basal Forebrain-Evoked Cerebral Blood Flow Response: A Role for Astrocytes?
It is believed that glutamate requires intermediary cells and mediators to alter CBF because there is no convincing evidence for dilatory glutamate receptors on brain vessels. In this respect, several AA metabolites display vasodilator properties (Figure 4E), and have been implicated as possible mediators of glutamate-induced CBF changes. The fact that BF stimulation activated COX-2 pyramidal neurons, which have a central role in functional hyperemia to sensory stimulation (Lecrux et al, 2011; Niwa et al, 2000), raised the possibility that COX-2 products may participate in the BF-evoked hemodynamic response. Surprisingly, selective inhibition of COX-2 with NS-398 had no impact on the CBF response to BF stimulation (+ 6.9% ± 9.5% at 45 minutes, ns,
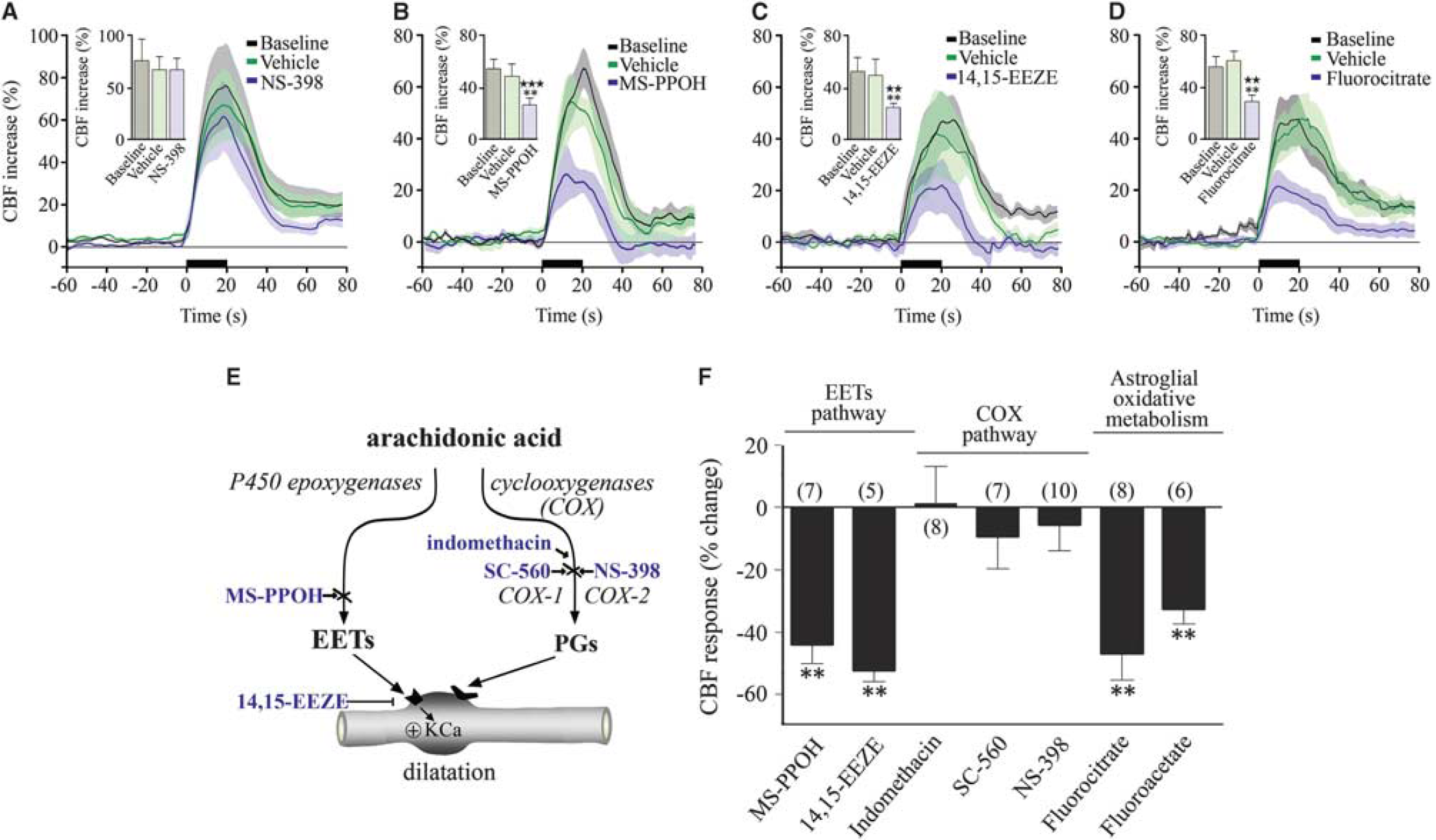
Astroglial pathways and epoxyeicosatrienoic acids (EETs) contribute to the neurovascular coupling response to basal forebrain (BF) stimulation. (
Consequently, we investigated whether EETs, AA products synthesized through the cytochrome P450 epoxygenase pathway (Figure 4E) that mediate an important part of the functional hyperemic response to sensory stimulation (Lecrux et al, 2011; Leithner et al, 2010; Peng et al, 2004) could be involved. Using MS-PPOH, a specific substrate inhibitor of P450 epoxygenase and 14,15-EEZE, an EETs receptor antagonist (Koehler et al, 2009), we found respective decreases of 44.2% ± 6.2% (
Epoxyeicosatrienoic Acids as Intermediaries for Glutamate and γ-Aminobutyric Acid Neurons
The role of glutamate in neurovascular coupling has been linked, primarily through mGluR5, to the synthesis and release of vasodilatory EETs (Shi et al, 2008), although these receptors would not be required for the initial hemodynamic response (Calcinaghi et al, 2011). Here, we assessed whether the glutamate receptors predominantly involved in the BF-induced increases in CBF also engage the EETs signaling cascade. We coadministered the selective NMDA or AMPA/kainate receptor antagonist with the P450 epoxygenase inhibitor MS-PPOH. Such treatments exerted no additive effect compared with each compound alone (–34.5% ± 3.5%,
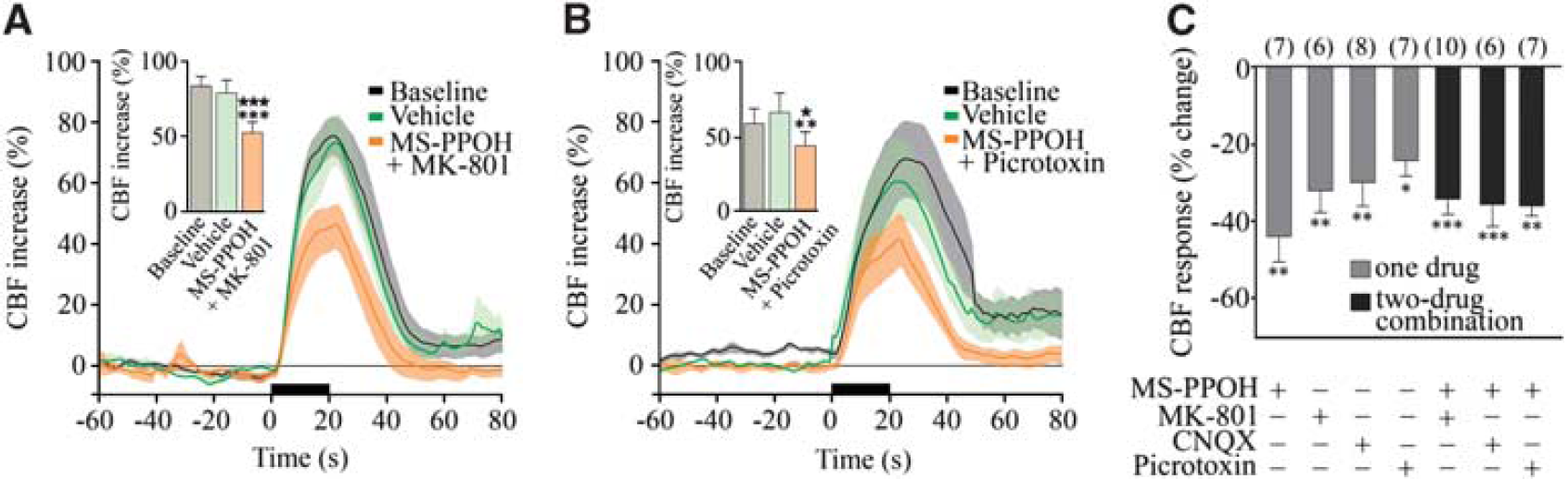
Glutamate and GABA (γ-aminobutyric acid) networks modulate the evoked cerebral blood flow (CBF) through epoxyeicosatrienoic acids (EETs). Average CBF responses to basal forebrain (BF) stimulation were reduced by MS-PPOH injection combined to MK-801 (
Discussion
The key new findings from our study are (1) activation of glutamate-releasing pyramidal cells, acting though various receptor subtypes, is required for the full expression of the neurovascular coupling response to BF stimulation; (2) COX-2 vasoactive products do not contribute to the CBF response, suggesting that COX-2 pyramidal cells recruited (c-Fos positive) by BF stimulation act through glutamate; (3) NMDA- and GABA-A-receptor-mediated pathways contribute in parallel to the CBF response; (4) EETs are essential in this neurovascular response, possibly acting as intermediaries for both pyramidal cells and GABA interneurons; and (5) metabolically active astrocytes are required, likely to synthesize and release vasoactive EETs.
Glutamate and γ-Aminobutyric Acid in the Perfusion Response to Basal Forebrain Stimulation
The cellular interactions that lead to increased cortical activity following BF stimulation are complex and likely not limited to ACh (Henny and Jones, 2008). However, there is substantial evidence that the neurovascular coupling response to the BF is predominantly driven by ACh: it occurs concurrently with the local release of ACh (Zhang et al, 1995), it is virtually abolished (–85%) by selective cortical cholinergic deafferentation (Kocharyan et al, 2008) and following blockade of mAChR and nicotinic receptors (Biesold et al, 1989). Here, we found that combined blockade of mAChRs and either NMDA or GABA-A receptors had no additive attenuating effect over mAChR blockade alone (∼60%), indicating that activation of these pathways was downstream to mAChR activation and reflected primarily activation of cortical glutamate and GABA neurons targeted by incoming BF afferents.
Accordingly, BF stimulation was accompanied by widespread increased activity (detected by c-Fos) in cortical pyramidal cells, consistent with their innervation by ACh BF terminals (Houser et al, 1985). Although ACh exerts mixed effects on pyramidal cells depending on cortical layers, subdivisions, and receptor subtypes (Gulledge et al, 2009; McCormick and Prince, 1986), strong mAChR-mediated depolarization leading to tonic firing (Carr and Surmeier, 2007) and mAChR-induced persistent spiking (Rahman and Berger, 2011) of pyramidal cells are prominent. Increased firing in pyramidal cells induced by BF stimulation further results from their disinhibition following mAChR activation of electrically coupled SOM interneurons and subsequent silencing of other GABA interneurons, that normally drive pyramidal cell inhibition (for detail, see Kocharyan et al, 2008). Hence, together the combined effects of ACh on pyramidal cells and cortical networks of inhibitory neurons contribute to the overall increased activity in glutamate-releasing pyramidal cells following BF stimulation (Figure 6). This dual mechanism of cortical activation concurs with our findings that combined blockade of NMDA and GABA-A receptors attenuated the BF-evoked CBF response significantly more than blocking either receptor alone.
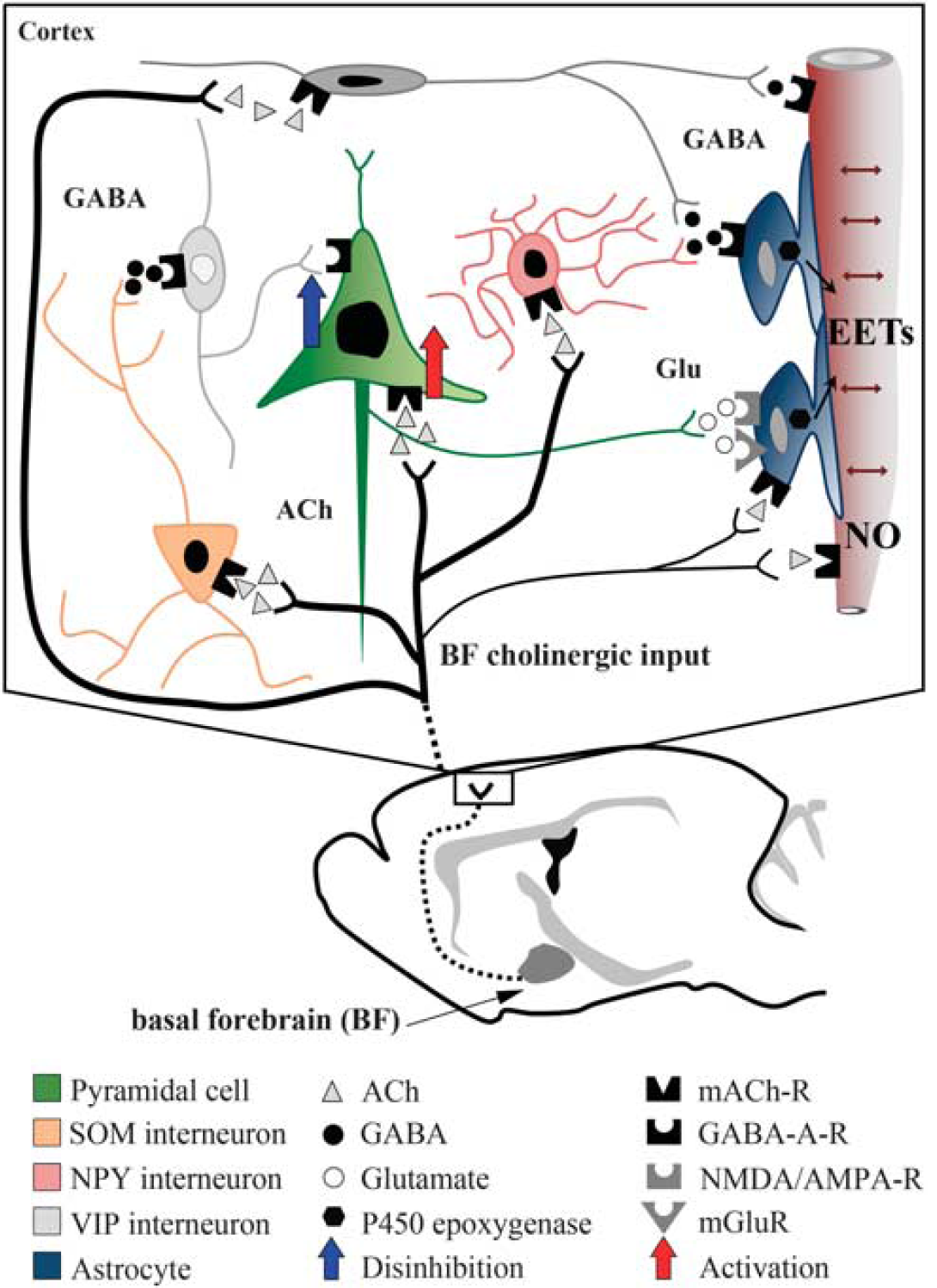
Schematic representation of how glutamate, GABA (γ-aminobutyric acid), and astrocytes may interact in the evoked cerebral blood flow (CBF) response to basal forebrain (BF) stimulation. Our c-Fos studies (black nuclei = activation, white nuclei = no activation or inhibition) indicate that BF cholinergic fibers activate both GABA interneurons—in particular, the somatostatin (SOM) subfamily (Kocharyan et al, 2008)—and glutamate-releasing pyramidal cells. Our pharmacological studies show that glutamate (Glu) and GABA effects are downstream of mAChR (ACh through muscarinic receptor) activation, and that pyramidal cells act through the release of glutamate, and not through cyclooxygenase-2 (COX-2) metabolites. Increased activity in glutamatergic pyramidal cells, either activated directly by acetylcholine (ACh) (red arrow) or indirectly by disinhibition (blue arrow) following activation of GABAergic interneurons, will primarily exert its CBF increasing effect through the epoxyeicosatrienoic acids (EETs), likely released from astrocytes. Direct vascular effects of ACh through mAChR-induced endothelial nitric oxide (NO)-mediated dilation also account for a significant part of the evoked CBF response. Similar, albeit small, dilatory effects of GABA through vascular GABA-A receptors cannot be discarded (Lecrux et al, 2011), as well as possible effects of ACh on astrocytes.
We provided evidence that following its release from activated pyramidal cells, glutamate can act on AMPA, NMDA and, to a lesser extent, group1 mGlu receptors that are expressed by several cortical neurons (Baude et al, 1993) and by astrocytes (Lalo et al, 2006). As mGluRs are normally recruited for selective circuit activation during glutamate spillover (Iserhot et al, 2004), this may explain their relatively modest contribution in the ACh driven-CBF response as compared with that induced by sensory stimulation that is driven by glutamate released by both thalamocortical afferents (Zonta et al, 2003) and local pyramidal cells (Lecrux et al, 2011). The previous (Kocharyan et al, 2008) and current c-Fos data correlated very well with our pharmacological findings, further emphasizing the contribution of excitatory and inhibitory neurons in the neurovascular coupling response to BF afferents, as also reported for thalamocortical and cortico-cortical afferent pathways (Enager et al, 2009; Lecrux et al, 2011).
Cyclooxygenase Metabolites do not Couple Neural Activity to Cerebral Blood Flow Following Basal Forebrain Stimulation
Nuclear c-Fos was upregulated in COX-2 pyramidal cells, whose products have a substantial role in the functional hyperemic response to incoming glutamatergic afferents following sensory stimulation (Lecrux et al, 2011; Niwa et al, 2000). However, neither COX-2 nor COX-1 products were found to significantly contribute to the BF-induced CBF response. We conclude that COX-2 activation and contribution to functional hyperemia requires high levels of glutamate that cannot be reached following activation of a cholinergic pathway. Evidence supporting this hypothesis includes the failure of mAChR activation to stimulate COX-2 in rat cerebral cortex (Orman et al, 2006), whereas NMDA receptors induce immediate release of neuronal COX-2-derived prostaglandin E2 (Pepicelli et al, 2005). Additionally, light-evoked vasodilatation in the retina, a pathway where neurovascular coupling does not involve glutamate but purinergic transmission, likewise did not operate via COX-2 products (Metea and Newman, 2006).
Astroglial Cells and Epoxyeicosatrienoic Acids in the Cerebral Blood Flow Response to Basal Forebrain Stimulation
Fluorocitrate and fluoroacetate are preferential inhibitors of astroglial oxidative metabolism
Conclusions
Our anatomical and
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.