
Abstract
We tested the hypothesis that Niaspan (a prolonged release formulation of niacin) increases tumor necrosis factor-α-converting enzyme (TACE) expression and Notch signaling activity and promotes arteriogenesis after stroke. Rats were subjected to middle cerebral artery occlusion and were treated with or without Niaspan. Niaspan significantly elevated local cerebral blood flow, and increased arteriogenesis as indicated by increased arterial diameter and vascular smooth muscle cell (VSMC) proliferation in the ischemic brain after stroke. The increased arteriogenesis significantly correlated with the functional outcome after stroke. Niaspan treatment of stroke upregulated TACE, Notch1, and Notch intracellular domain expression in the ischemic brain. To further investigate the mechanisms of Niaspan-induced arteriogenesis, a primary brain arterial culture was used. Niacin treatment significantly increased arterial sprouting and VSMC migration compared with control nontreated arterial cells. Inhibition of TACE by the TACE inhibitor or knockdown of TACE gene expression in brain arterial culture significantly attenuated Niacin-induced arterial sprouting and VSMC migration. In addition, TACE treatment of arterial culture significantly increased arterial VSMC migration and arterial sprouting. Knockdown of Notch1 marginally decreased arterial sprouting and VSMC migration compared with scrambled control. Niaspan promotes arteriogenesis, which is mediated, in part, by TACE.
Introduction
Arteriogenesis is an important process for adapting the preexisting circuit of vessels into functional collateral conduits for the delivery of oxygen -enriched blood to tissue distal to occlusion of a large, peripheral conduit artery. In brain circulation, collateral circulation defines the extent of the ischemic penumbra, providing blood flow to tissues at risk of infarction downstream from an occluded artery (Liebeskind, 2004). Collateral perfusion demarcates the ischemic penumbra and alters clinical outcome (Liebeskind, 2005). Early clinical improvement after stroke is linked to the presence of arteriolar collaterals. The presence of arteriogenesis is predictive of improved long-term clinical outcome in patients treated with and without thrombolysis for stroke (Christoforidis et al, 2005).
Arteriogenesis is modulated by many factors, such as chemotactic factors, nitric oxide synthase, growth factors, and proteases (Deindl and Schaper, 2005; Heil and Schaper, 2004, 2005). The initial step in arteriogenesis is the development of elevated shear stress against the wall of the arteriole that stimulates endothelial cells to react by activating cytokines, such as tumor necrosis factor-α (TNF-α), monocyte chemoattractant protein-1, and cell adhesion molecules (Hoefer et al, 2002). Tumor necrosis factor-α serves as a pivotal modulator of arteriogenesis, which is attenuated by treatment with TNF-α inhibitors (Grundmann et al, 2005). The TNF-α-converting enzyme (TACE) is the principal protease involved in the activation of pro-TNF-α (Edwards et al, 2008) and regulates the function of several transmembrane proteins and cell adhesion molecular shedding (Garton et al, 2003). Tumor necrosis factor-α-converting enzyme also plays a prominent role in the activation of the Notch signaling (Ehebauer et al, 2006). The Delta/Notch signaling pathway mediates arteriogenesis and angiogenesis (Liu et al, 2003). In zebrafish, Notch signaling is required for arterial identity by suppressing the venous fate in developing artery cells (Lawson et al, 2002). Notch signaling in vascular smooth muscle cells (VSMCs) is required to pattern the cerebral vasculature (Proweller et al, 2007). Whether TACE induces arteriogenesis has not been investigated.
Niacin (nicotinic acid) is an effective medication in the clinical use for increasing high-density lipoprotein cholesterol. Elevated high-density lipoprotein cholesterol is associated with higher collateral grade in coronary artery disease (Pohl et al, 2001). Our previous studies have found that Niaspan (a prolonged release formulation of niacin) increases high-density lipoprotein cholesterol level and upregulates angiogenesis after stroke (Chen et al, 2007). Angiogenesis differs from arteriogenesis. Angiogenesis is the formation of a capillary network, through the activation and proliferation of endothelial cells. Arteriogenesis consists of the formation of new arterioles, which presumably occurs when preexisting capillaries acquire smooth muscle coating, and these newly formed and/or preexisting arterioles transform into channels with larger diameters (van Royen et al, 2001). Whether Niaspan treatment of stroke alters arteriogenesis and the mechanisms underlying Niaspan-induced arteriogenesis are unknown. This study is the first to examine the effects and the mechanisms of Niaspan treatment of stroke on arteriogenesis in a rat model of middle cerebral artery occlusion (MCAO).
Materials and methods
Animal Middle Cerebral Artery Occlusion Model and Experimental Groups
Adult male Wistar rats weighing 270 to 300 g were used in all our experiments. Transient right MCAO was induced for 2 h by advancing a 4-0 surgical nylon suture from the external carotid artery into the lumen of the internal carotid artery to block the origin of the MCA (Chen et al, 2001). Sham-operated rats underwent the same surgical procedure without suture insertion. Rats were randomized and gavaged starting 24 h after MCAO with (
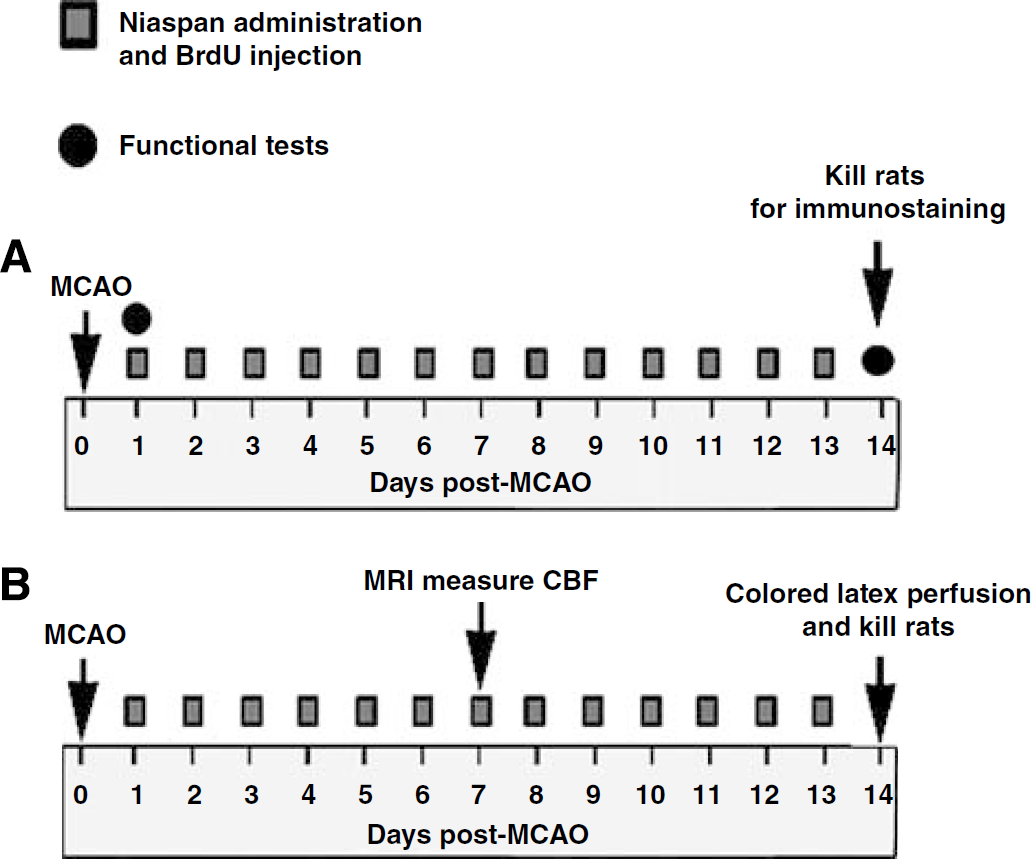
Experimental timeline profile.
Neurologic Functional Tests
Foot-fault and a modified neurologic severity score evaluation were carried out before MCAO, and at 14 days after MCAO by an investigator who was blinded to the experimental groups. Functional and angiogenesis data from these animals have been reported previously (Chen et al, 2007).
Cerebral Blood Flow Measurement by Magnetic Resonance Imaging
To test whether Niaspan treatment regulates cerebral blood flow (CBF), another set of rats were subjected to 2h of MCAO and treated with or without Niaspan (40 mg/kg) daily. Cerebral blood flow was measured at 7 days after MCAO (
Colored Latex Perfusion
To test cerebral arterioarchitecture and the diameter of the circle of Willis, rats were killed at 14 days after MCAO and perfused with carbon black-stained latex (
Histologic and Immunohistochemical Assessment
Rats were killed 14 days after stroke. The brains were fixed by transcardial perfusion with saline, followed by perfusion and immersion in 4% paraformaldehyde before being embedded in paraffin. A standard paraffin block was obtained from the center of the lesion (bregma −1 mm to + 1 mm). A series of 6-μm thick sections were cut from the block. Every 10th coronal section for a total five sections was used for immunohistochemical staining. Antibody against BrdU, a proliferating cell marker (1:100, Boehringer Mannheim, Indianapolis, IN, USA), smooth muscle actin (α-SMA, mouse monoclonal IgG 1:800, Dako); Notchl (1:100 dilution, Santa Cruz Biotechnology, Santa Cruz, CA, USA), Notch intracellular domain (NICD, rabbit polyclonal IgG 1:1000 dilution, Abcam), and TACE (1:200 dilution, Santa Cruz, CA, USA) immunostaining were carried out. Control experiments consisted of staining brain coronal tissue sections as outlined above, but nonimmune serum was substituted for the primary antibody. The immunostaining analysis was carried out by an investigator blinded to the experimental groups.
Double Immunohistochemical Staining
To specifically identify BrdU, Notchl, NICD-reactive cells co-localized with SMCs, double immunofluorescence staining of BrdU/α-SMA, Notchl/α-SMA and NICD/α-SMA were carried out. FITC (Calbiochem) and cyanine-5.18 (CY5, Jackson Immunoresearch) were used for double-label immunoreactivity. Each coronal section was first treated with the primary anti-α-SMA antibody with Cy5, and then followed by BrdU, Notchl, or NICD with FITC. Control experiments consisted of staining brain coronal tissue sections as outlined above, and nonimmune serum was used in place of the primary antibody.
Arterial Diameter Measurement
α-SMA immunoreactivity was used as a marker to identify arteries (Ho et al, 2006). Consequently, five sections from the standard reference coronal section were acquired, and the α-SMA-stained thick wall vessels and diameter ≥ 10 μm arterial diameters were measured in the ischemic boundary (IBZ) and in the surface of cortex of the ipsilateral using the MCID (Imaging Research, St Catharines, Canada) computer imaging analysis system.
BrdU, Notch, and Notch Intracellular Domain Expression Quantification
Double immunostaining BrdU, Notchl, or NICD with α-SMA were carried out. BrdU, Notchl, or NICD positive VSMC number were counted in a total of 20 enlarged arteries located in the IBZ and surface of cortex of the ipsilateral in each section. The number of α-SMA positive cells colocalized with DAPI were counted as the total SMCs. BrdU, Notchl, or NICD positive cells colocalized with α-SMA were counted as BrdU, Notchl, or NICD positive SMCs, respectively. Data were analyzed in a blinded manner and presented as percentage of the number of the BrdU, Notchl, or NICD immunoreactive SMCs/total SMCs.
Tumor Necrosis Factor-α-Converting Enzyme Expression Quantification
The TACE immunostained sections were digitized using a x 40 objective (Olympus BX40) via the MCID system. The TACE positive cell number was counted in 20 enlarged arteries localized in the IBZ in each section. Five sections and eight views (Chen et al, 2007) in each section were counted per rat with the number of TACE reactive cells averaged. The total number of hematoxylin positive nuclear cells localized in the arteries were counted. The data are presented as the percentage of the TACE immunoreactive cells in artery/total cell number in artery.
Rat Brain Microvascular Endothelial Cell and Vascular Smooth Muscle Cell Cultures
Rat brain was obtained, placed in 2 ml of wash buffer (RMPI 1640 + 2%FBS + 1%Pen/Strep), and briefly homogenized. The resulting suspension was mixed with a 30% Dextran solution and centrifuged. The pellet was resuspended in digestion buffer (RPMI 1640 + 2%FBS), to which collagenase/dispace (Roche) was added. The resulting suspension was centrifuged and mixed with Percoll (Sigma). The final cell pellet was resuspended in endothelial cell growth media, and allowed to culture for 24 h. Rat brain microvascular endothelial cells and VSMCs (ATCC, CRL-2018) were cultured and treated with (
Western Blot
Rats were killed at 8 days after MCAO and brain tissues were extracted from the ischemic core, IBZ, and homologous contralateral tissue (Chen et al, 2008). In addition, the MCA, PCA, and internal carotid artery were isolated from ipsilateral cerebral arteries. Equal amounts of cell lysate were subjected to western blot analysis (Chen et al, 2003). Specific proteins were visualized using a Super-Signal West Pico chemiluminescence kit (Pierce). The following primary antibodies were used: anti-β-actin (1:2000; Sigma, St Louis, MO, USA), anti-Notch1 (1:500 dilution), anti-NICD (1:1000 dilution), and anti-TACE (1:500 dilution).
Primary Arterial Culture
To elucidate the mechanisms underlying Niaspan mediation of arteriogenesis, a primary cerebral artery culture model was used (Saward and Zahradka, 1997). Rats were subjected to 2h of MCAO and were killed at 8 days after stroke. The PCA were removed from rat brain. The segments of these arteries were placed into matrigel. The cerebral artery was treated with (1) nontreatment for control; (2) +TACE (100 nmol/L, R&D Systems Human Recombinant Human TACE/ADAM17); (3) +TAPI-2 (10 μmol/L, Peptides International); (4) + Niacin (1 mmol/L); and (5) Niacin + TAPI-2 for 7 days. TAPI2 is a TACE inhibitor (Katakowski et al, 2007). After incubation, dishes were observed at 7 days with a phase contrast microscope and photographed at x 4 magnification. The lengths of SMC migration and arterial sprouting were measured.
Notch1 and Tumor Necrosis Factor-α-Converting Enzyme Small Interfering RNA in Primary Arterial Culture
The PCA was removed from rat brain and segments of arteries were placed into matrigel. Notchl small interfering RNA (siRNA) and TACE siRNA (Santa Cruz Biotech) were transfected using Lipofectamine, 2000 (Invitrogen), following standard protocol. Briefly, arterial sections were put in Matrigel in 24-well plates and were transfected with 1 μg of TACE siRNA, Notchl siRNA, or scrambled RNA per well, respectively. Media were added to each well and transfection cocktail was added. The knockdown TACE was treated with or without Niacin (1 mmol/L) for 7 days. The length of VSMC migration and arterial sprouting was measured.
Real-Time Polymerase Chain Reaction
The cells were harvested and total RNA was isolated from treated cells with TRIzol (Invitrogen), following a standard protocol. Quantitative PCR was carried out using the SYBR Green real-time PCR method on an ABI 7000 PCR instrument (Applied Biosystems, Foster City, CA, USA) using three-stage program parameters provided by the manufacturer, as follows; 2 min at 50°C, 10 min at 95°C, and then 40 cycles of 15 s at 95°C and 1 min at 60°C. The following primers for real-time PCR were designed using Primer Express software (ABI). Notchl Fwd: CTGTACAG AGGATGTGGACGAA; Rev: TGACACACACACAGTTGTA GCC. and TACE Fwd: ACTCTGAGGACAGTTAACCAAA CC; Rev: AGTAAAAGGAGCCAATACCACAAG; Hes1: Fwd: ACACCGGACAAACCAAAGAC; Rev: ATGCCGGGA GCTATCTTTCT. GAPDH Fwd: AGAACATCATCCCTGCA TCC; Rev: CACATTGGGGGTAGGAACAC. Each sample was tested in triplicate, and the samples were obtained from three independent experiments that were used for analysis of relative gene expression data using the 2−ΔΔCT method.
Statistical Analysis
Comparisons within treatment groups of arteriogenesis data measured on day 14 were made using two-way analysis of variance. Variables included BrdU, arterial diameter, Notchl, NICD and TACE expression in the ischemic brain, and artery SMC migration. If an overall treatment group effect was detected at
Results
Niaspan Treatment of Stroke Increases Arteriogenesis
To test whether Niaspan increases arteriogenesis, α-SMA immunostaining was carried out on brain coronal sections. Figures 2A and 2B show that treatment with Niaspan significantly increased arterial diameter (A) and BrdU positive VSMCs (B) in the IBZ compared with the control MCAO rats. Correlation coefficient analysis shows strong negative correlations between Foot-fault (14 days after MCAO) and artery diameter (C,
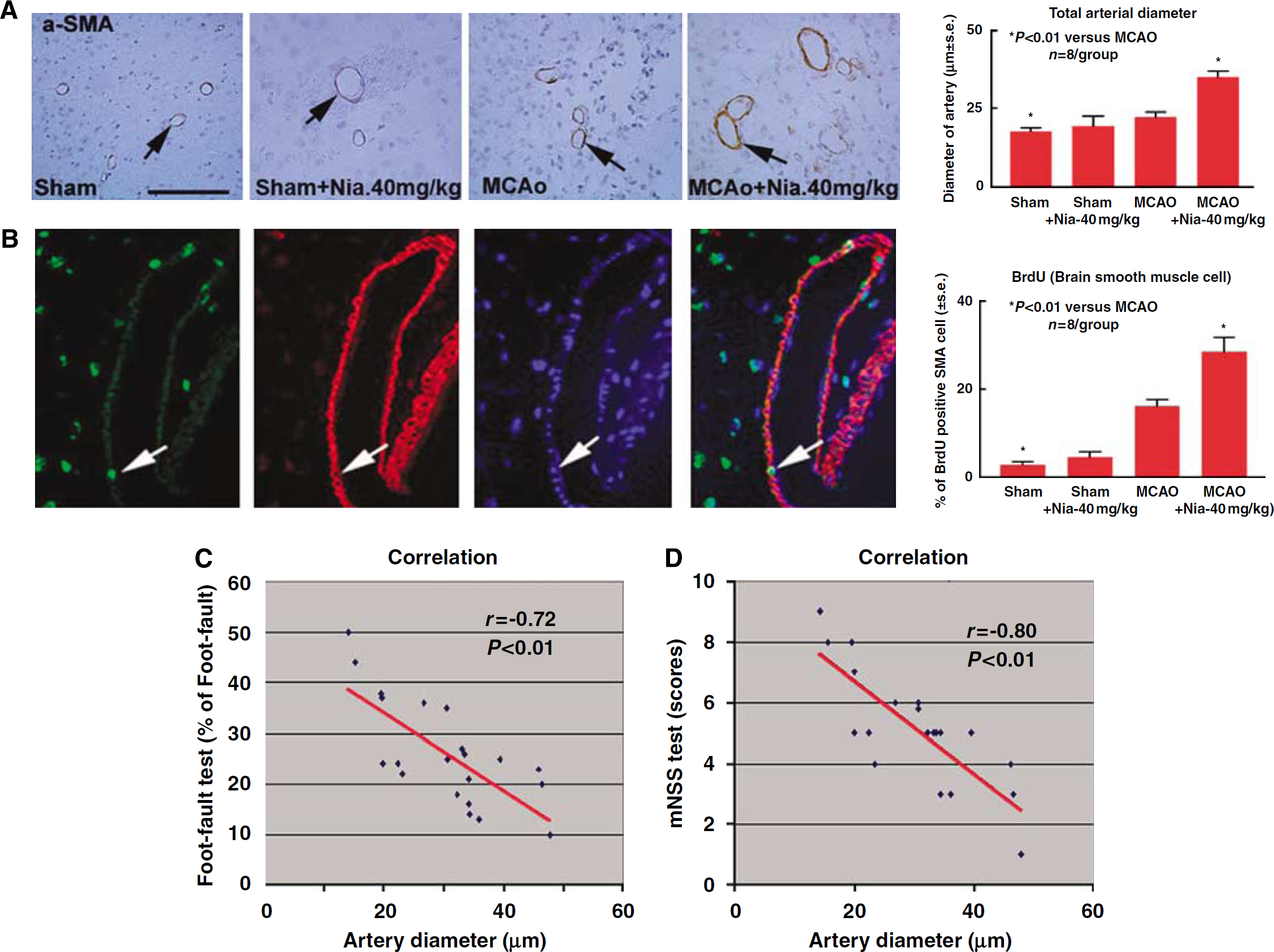
Niaspan treatment of stroke increases arteriogenesis in the ischemic brain. (
Niaspan Treatment of Stroke Increases Focal Cerebral Blood Flow in the Ischemic Brain and Increases the Diameter of the Circle of the Willis Arteries
Arteriogenesis serves as the most efficient mechanism to restore flow after arterial occlusion (Hershey et al, 2001). As our previous study found that Niaspan treatment improved functional outcome start from 7 days after stroke (Chen et al, 2007), we tested whether Niaspan treatment increases CBF after stroke using magnetic resonance imaging measurements at 7 days after MCAO. Figures 3A-E show that the CBF was significantly increased in Niaspan treated group (D) compared with control MCAO group (C) in the ipsilateral hemisphere. Rats were killed at 14 days after MCAO and perfused with colored latex. Figures 3F-N show that Niaspan treatment of stroke rats significantly increased the MCA and internal carotid artery diameter compared with control MCAO rats (
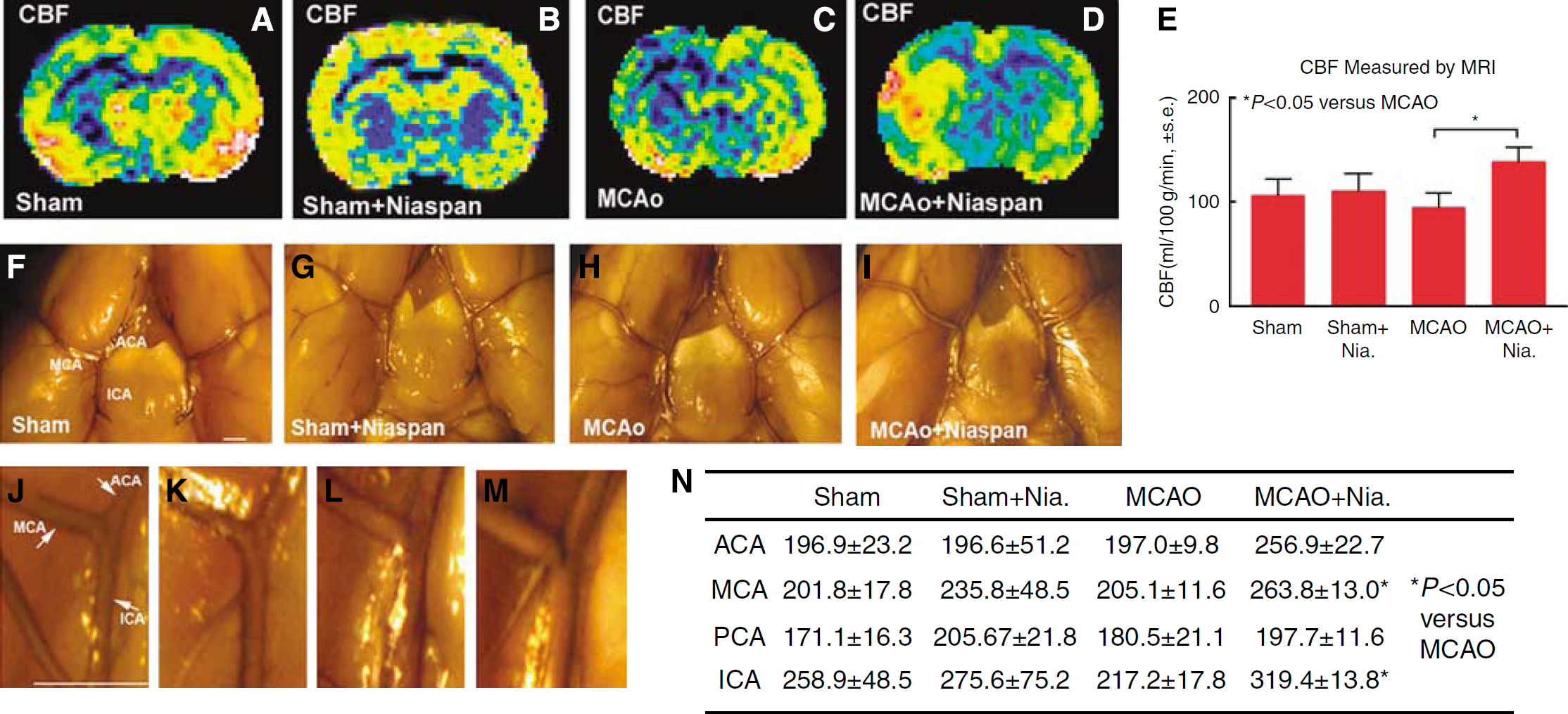
Niaspan treatment of stroke increases CBF and arteries diameter in the ischemic brain. (
Niaspan Treatment of Stroke Promotes Notchl, Tumor Necrosis Factor-α-Converting Enzyme, and Notch Intracellular Domain Expression after Stroke
Vascular expression of Notch pathway receptors and ligands is restricted to arterial vessels (Villa et al, 2001). The TACE is a proteolytic cleavage enzyme involved in Notch signaling activation (Mumm et al, 2000). To test the mechanism of Niaspan-induced arteriogenesis, TACE, Notchl, and NICD expression in the ischemic brain were measured. Figures 4A-C show that Niaspan treatment significantly increased arterial SMC TACE, Notchl, and NICD (activated form of Notch) expression in the ischemic brain arteries compared with control MCAO rat.
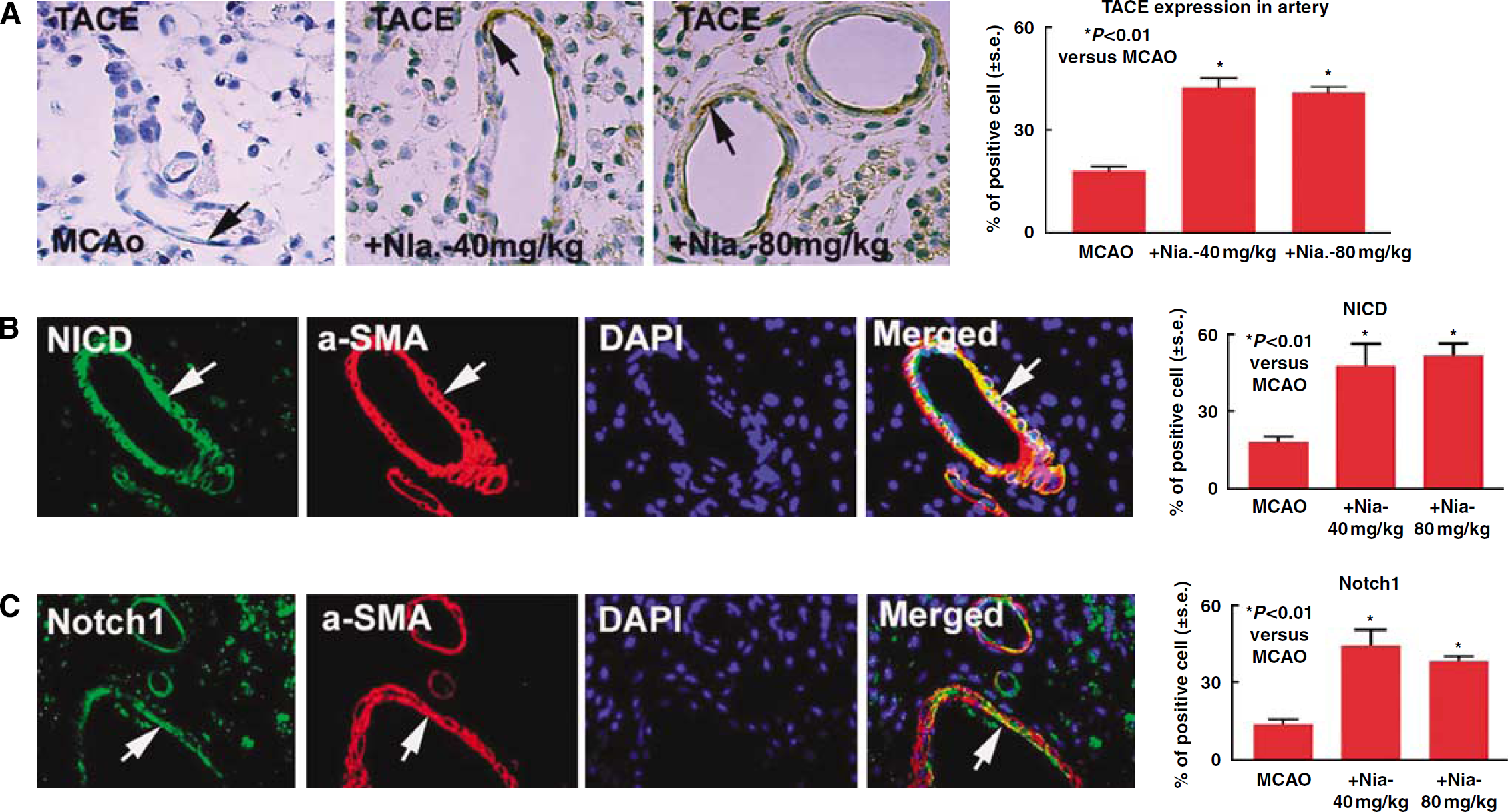
Niaspan treatment of stroke increases TACE, Notchl, and NICD expression in the ischemic brain. (
Tumor necrosis factor-α-converting enzyme, Notchl, and NICD expression were also measured by western blot. Brain tissues were extracted from the ischemic core, IBZ, and homologous contralateral tissues (Chen et al, 2008). Cerebral arteries were isolated from the ipsilateral hemisphere. Figure 5A shows that Niaspan treatment of stroke rats increased NICD protein expression in the ischemic core, IBZ and contralateral tissue, and increased TACE expression in the IBZ compared with control MCAO rats, respectively. Using real-time PCR and western blot assay, Figures 5B and 5C show that Niaspan treatment increased cerebral artery Notchl, TACE gene (B) and protein (C) expression compared with MCAO control artery.
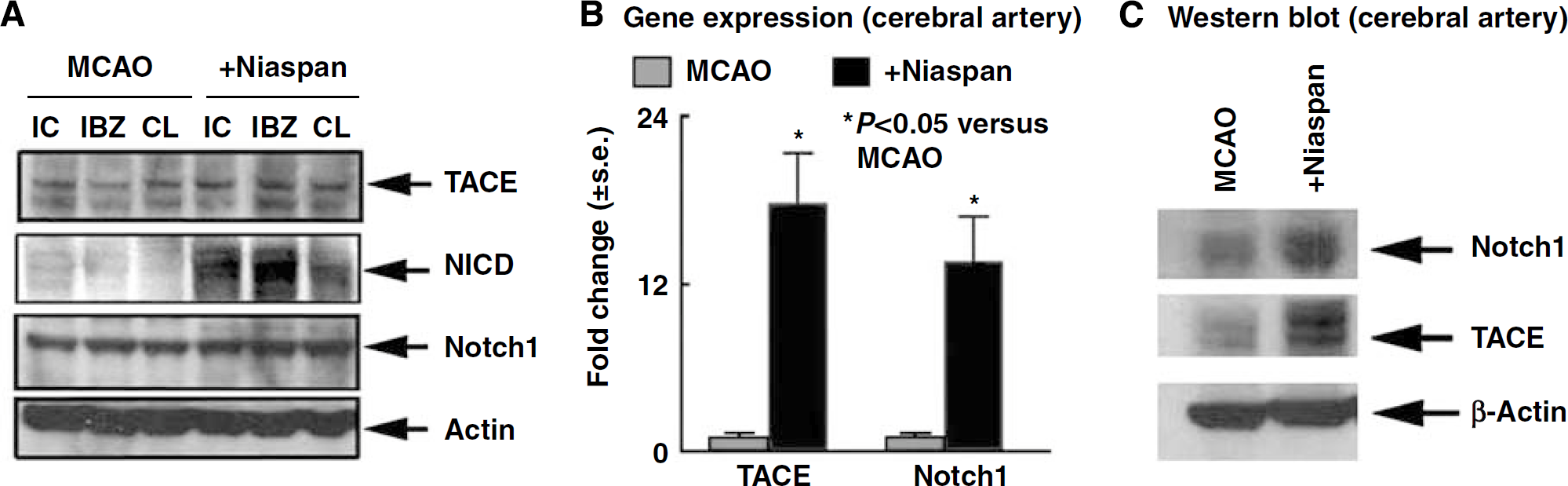
Niaspan increases Notchl, NICD, and TACE expression in the ischemic brain. (
Niacin Treatment Regulates Rat Brain Microvascular Endothelial Cell and Vascular Smooth Muscle Cell Tumor Necrosis Factor-α-Converting Enzyme and Notchl Gene and Protein Expression
To test whether Niacin regulates brain endothelial cell and VSMC TACE and Notchl gene and protein expression, rat brain microvascular endothelial cell (RBMEC) and VSMC cultures were used. Figure 6 shows that Niacin treatment of RBMECs significantly increased TACE gene (A) and TACE and Notchl protein (C) expression compared with control. Niacin treatment of VSMC increased TACE and Notchl gene (B) and protein (D) expression compared with nontreatment control.
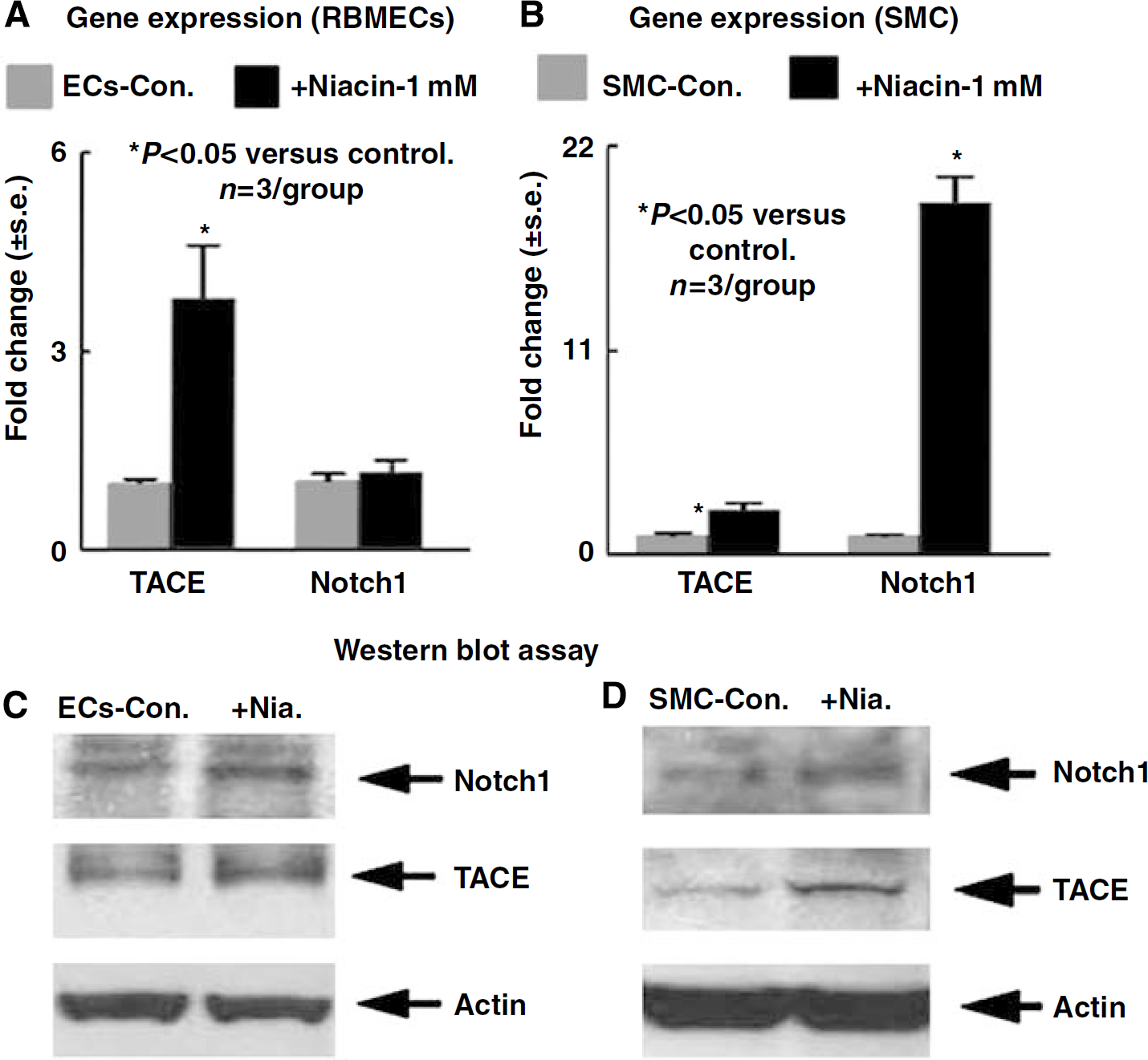
Niacin regulates Notchl, TACE gene, and protein expression in cultured RBMECs and VSMCs. (
Niacin and Tumor Necrosis Factor-α-Converting Enzyme Increase Cerebral Artery Smooth Muscle Cell Migration In Vitro
The stages of arteriogenesis consist of arteriolar thinning, followed by transformation of VSMCs from the contractile into the proliferative and synthetic phenotype (Scholz et al, 2000). Endothelial cells and VSMCs proliferate, and VSMCs migrate to form a neointima (Scholz et al, 2000). To test whether Niacin regulates arteriogenesis, arterial sprouting and VSMC migration were measured in primary artery culture. Figure 7 shows that PCA from stroke rat treated with Niacin (B) significantly increased arterial sprouting and cell migration compared with nontreatment control (A). Inhibition of TACE with TAPI2 (E) or knockdown of TACE (G) in arterial culture significantly attenuated Niacin-induced arterial sprouting and cell migration. These data suggest TACE signaling may regulate Niacin-induced arteriogenesis.
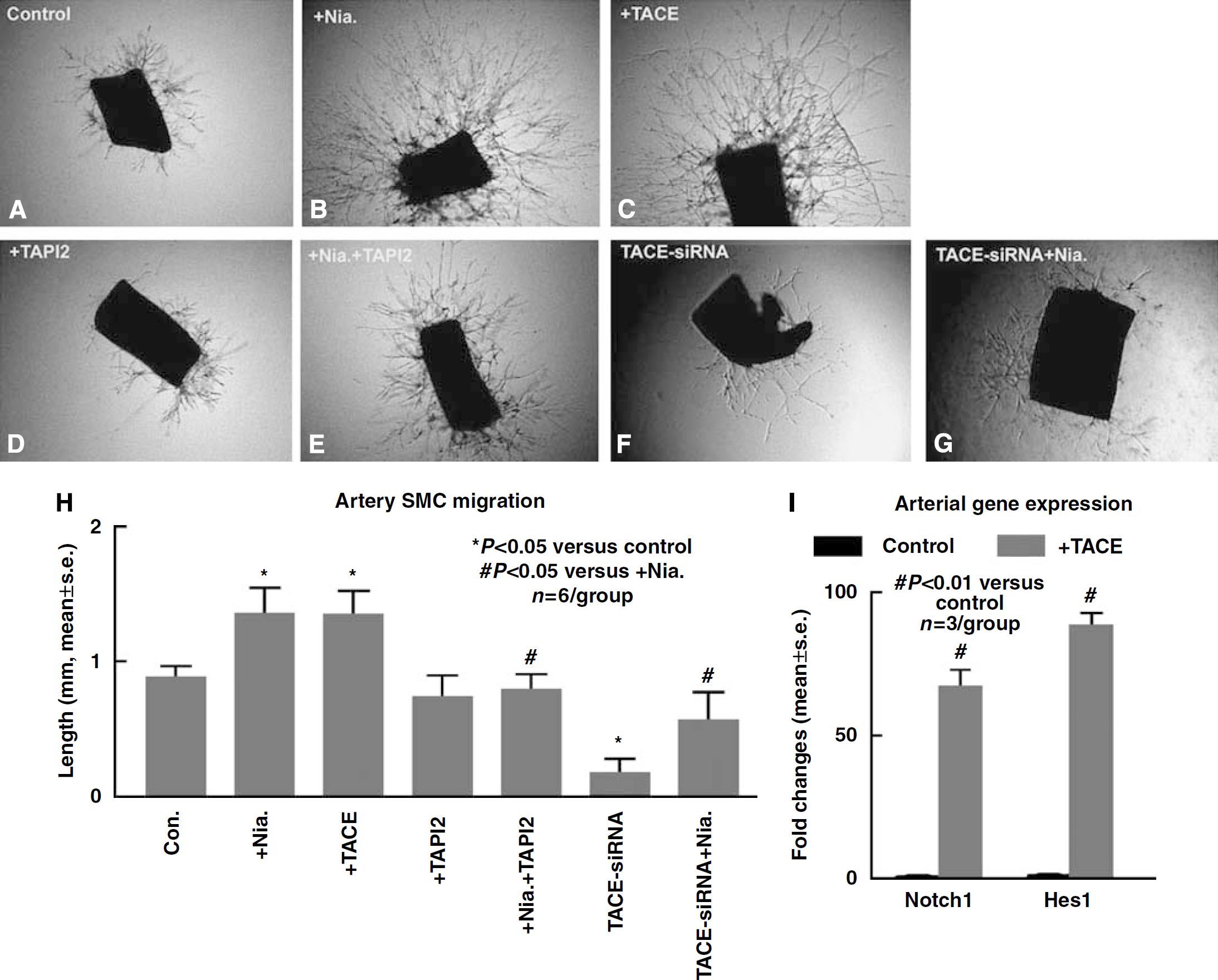
Niacin and TACE increase arterial SMC migration. Inhibition of TACE decreased Niacin-induced arterial SMC migration. (
In addition, TACE treatment (C) also significantly increased arterial sprouting and cell migration compared with nontreatment control (
To further test whether Notchl regulates arteriogenesis, arterial sprouting and VSMC migration were measured in Notchl knockdown arterial segments in culture. The data show that knockdown of Notchl gene expression by siRNA marginally (0.6 ± 0.1 mm,
Discussion
We show that (1) Niaspan treatment of stroke starting at 24 h after onset of MCAO significantly promoted arteriogenesis and increased CBF in the ischemic brain. The increased arteriogenesis significantly correlated with functional outcome after stroke. (2) Treatment of stroke with Niaspan significantly increased TACE, Notchl, and NICD expression in the ischemic brain arteries. (3) Niacin and TACE increased sprouting and cell migration in arterial cultures. Inhibition of TACE or knockdown of TACE gene expression by siRNA significantly decreased Niacin-induced cerebral arterial SMCs migration and arterial sprouting
Niaspan Promotes Arteriogenesis
After arterial occlusion, blood vessels respond by angiogenesis and arteriogenesis. Vascular smooth muscle cells, contractile and matrix-producing support cells that are associated with arteries and, less prominently, with veins, play critical roles in vascular maturation and arteriogenesis (Heil et al, 2006). Arteriogenesis is modulated by many factors and is linked to elevated pressure, which increases radial wall stress, and elevated flow. Occlusion of a major artery and elevated fluid shear stress are important for induction of arteriogenesis. In contrast to the normal brain, the ischemic brain is thus primed for arteriogenesis. Our data show that Niaspan treatment of stroke significantly increases CBF measured by magnetic resonance imaging and upregulates arterial diameter and VSMC proliferation in the ischemic brain. In addition, Niacin also promotes cultured cerebral arterial sprouting and VSMC migration
Tumor Necrosis Factor-α-Converting Enzyme Mediates Niaspan-Induced Arteriogenesis
Our data show that Niasapn treatment increases TACE expression in the ischemic brain tissue and in the cerebral arteries after stroke. The TACE treated artery significantly increased arterial sprouting and arterial SMCs migration in primary artery culture. Inhibition of TACE by TAPI2 and knockdown TACE gene expression by siRNA significantly attenuated Niacin-induced arterial sprouting and arterial SMC migration. Therefore, these data indicate TACE signaling mediates Niaspan-induced arteriogenesis after stroke. Although TACE cleaves and induces Notch signaling activity, TACE also mediates TNF-α shedding. Notch signaling and TNF-α all positively regulate arteriogenesis (Hoefer et al, 2002, 2004), and further studies investigating these downstream effects of TACE on arteriogenesis are warranted.
Niacin Increases Vascular Notch Signaling Activity
Functional studies have shown that the angiogenic growth of the blood vessel network, the proliferation of endothelial cells, and the differentiation of arteries and veins are controlled by Notch signaling (Roca and Adams, 2007). Notch signaling is an ancient intercellular signaling mechanism that plays a myriad of roles in the regulation of artery/vein differentiation in endothelial and VSMCs, and blood vessel sprouting and branching (Gridley, 2007). Arteries are major sites of Notch signaling in mice, and arterial specification is defective in various Notch pathway mutants (Vooijs et al, 2007). Notchl and Hey1/Hey2 knockout mice fail to express arterial endothelial markers in remaining large arteries (Fischer et al, 2004). Mice lacking Notch signaling in VSMCs are intolerant to reduced CBF and incur profound neurologic sequelae and death (Proweller et al, 2007). Dll4, a membrane-bound ligand for Notchl and Notch4, interacts with its cognate receptor Notchl on the surface of endothelial cells, and regulates vascular development and arteriogenesis (Ridgway et al, 2006). Our data show that Niaspan treatment of stroke rats significantly increased Notchl and NICD expression in the ischemic brain, which were specifically localized around the artery in the ischemic brain. Knockdown of Notchl gene expression in cerebral arterial culture by siRNA marginally decreased arterial sprouting and VSMC migration. Therefore, Niaspan regulation of Notchl expression may partially play a role in arteriogenesis after stroke. As Notch signaling pathways include many factors, for example, the four Notch receptors (Notch1-4), Jagged and Delta ligands (Dll1-4), we cannot exclude the possibility that these factors play a role in the regulation of arteriogenesis (Limbourg et al, 2007; Liu et al, 2003).
In summary, we show that treatment of experimental stroke with Niaspan at 24 h after stroke significantly promotes arteriogenesis and increases TACE signaling activity. The TACE plays an important role in Niaspan-induced arteriogenesis after stroke. Therefore, pharmacological attempts to stimulate the growth of arteriogenesis may provide a new treatment option for patients with stroke.
Footnotes
Acknowledgements
The authors thank Cynthia Roberts and Qinge Lu for technical assistance. This work was supported by National Institute of Neurological Disorders and Stroke grants RO1 NS047682, PO1 NS23393, RO1 AG031811, and American Heart Association grant 0750048Z.