
Abstract

Local Impermeant Anions Establish the Neuronal Chloride Concentration.
Glykys J, Dzhala V, Egawa K, Balena T, Saponjian Y, Kuchibhotla KV, Bacskai BJ, Kahle KT, Zeuthen T, Staley KJ. Science 2014;343:670–675.
Neuronal intracellular chloride concentration [Cl(−)](i) is an important determinant of γ-aminobutyric acid type A (GABA(A)) receptor (GABA(A)R)-mediated inhibition and cytoplasmic volume regulation. Equilibrative cation-chloride cotransporters (CCCs) move Cl(−) across the membrane, but accumulating evidence suggests factors other than the bulk concentrations of transported ions determine [Cl(−)](i). Measurement of [Cl(−)](i) in murine brain slice preparations expressing the transgenic fluorophore Clomeleon demonstrated that cytoplasmic impermeant anions ([A](i)) and poly-anionic extracellular matrix glycoproteins ([A](o)) constrain the local [Cl(−)]. CCC inhibition had modest effects on [Cl(−)] (i) and neuronal volume, but substantial changes were produced by alterations of the balance between [A](i) and [A](o). Therefore, CCCs are important elements of Cl(−) homeostasis, but local impermeant anions determine the homeostatic set point for [Cl(−)], and hence, neuronal volume and the polarity of local GABA(A)R signaling.
Commentary
It is now well established that GABA receptors can mediate either excitation or inhibition, depending on the GABA reversal potential (EGABA) of the neuron, which in turn is set by the transmembrane chloride (Cl) gradient. In mature neurons, the Cl concentration outside the cell (Clo) is higher than that inside the cell, so when GABA binds to its postsynaptic type-A receptor and opens chloride channels, Cl flows down its electrochemical gradient into the cell, leading to hyperpolarization. In neonatal neurons, however, the intracellular Cl concentration (Cli) is high enough such that GABA binding-induced chloride channel opening leads to Cl efflux from the neuron, leading to membrane depolarization. Depolarizing GABA action early in development is considered to be important for neuronal plasticity and circuit maturation (1) and has also been used to explain the enhanced susceptibility of the neonatal brain to seizures (2, 3). Phenobarbital (PHB), a GABA receptor agonist, has limited effectiveness as an anticonvulsant in neonatal seizures, an effect that has been attributed to this reversed Cl gradient. In accordance with this mechanism, PHB not only fails to stop neonatal seizures but can even exacerbate them. Beyond the neonatal period, GABA depolarizing actions may have wider ramifications in epilepsy. For example, GABA induces depolarization in amygdala neurons in temporal lobe epilepsy (4).
Two chloride cation cotransporters (CCCs) act in concert to govern a neuron's Cl concentration gradient. The sodium-potassium-chloride cotransporter 1 (NKCC1), predominates in the neonatal period and imports Cl into neurons. Later, expression of the potassium-chloride cotransporter 2 (KCC2) increases, with Cl transported out of the neuron. Thus, over development, KCC2 expression increases and NKCC1 expression decreases, such that when KCC2 predominates, the extracellular concentration of Cl is higher than its intracellular concentration. Until now, the explanation for the reversed chloride gradient and GABA-mediated depolarization early in development has implicated a developmental switch in the predominance of CCCs (3). Is the action of the CCCs sufficient to explain the relative differences in intra- and extracellular Cl concentrations? The present paper by Glykys et al. proposes that CCC actions, while conceptually attractive, fail to account fully for Cli and the observed GABA equilibrium potential, EGABA. Evidence is presented that Cli sets EGABA but that CCC expression does not fully explain Cli and EGABA.
If not CCC, what else could determine Cl concentration and thus the polarity of GABA action? Glykys et al. hypothesize that impermeant anions on both sides of the neuronal membrane contribute to Cl homeostasis. Intracellularly, impermeant anions consist of nucleotides with negatively charged phosphate groups and proteins with negatively charged amino and carboxyl groups. In the extracellular matrix, negatively charged sulfates on proteoglycans comprise the bulk of impermeant anions. Glykys et al. base their experiments on the assumption that the sum of intracellular anions and Cl must equal the sum of extracellular anions and Cl (Ai + Cli = Ao + Clo). There is a reciprocal relationship between Ai and Cli so that their sum must be constant—any increase in Cli must be balanced by a decrease in Ai, such that Cli is “constrained” by Ai. Similarly, in the extracellular matrix, Clo is constrained by Ao. To maintain Donnan equilibrium, an increase in either anion species within the intracellular or extracellular domain must balance. This requirement leads to several physiological predictions (5), which the authors tested experimentally in the current paper.
To determine whether intracellular and extracellular anions help set EGABA and thus determine the polarity of GABA responses, Glykys and colleagues first assessed Cli using Clomeleon, a fluorescent reporter of the intracellular Cl concentration. They found a wide variation in Cli in hippocampal CA1 neurons and neocortical pyramidal cells, in both neonates (in which NKCC1 predominates) and older animals (in which KCC2 predominates). Blockade of either NKCC1 or KCC2 did not alter the average Cli of the imaged neurons significantly, suggesting that some factor other than CCC mediates Cli and, thus, ECl and the direction of the GABA response. Likewise, when they altered the concentration of intracellular or extracellular anions, the chloride gradient varied in the predicted manner. In addition to the wide variation in Cli between cells, the authors also found considerable heterogeneity in subcellular distributions of Cli related to differences in impermeant anion concentrations. For example, Cli varied considerably among neuronal compartments (such as dendrites, somata, and axons), implying a functional role for microdomains of different concentrations of chloride and fixed anions.
These complex physiological observations have clinical relevance for epilepsy and for neonatal seizures in particular. Neonatal seizures are associated with cytotoxic edema (cell swelling and volume increase) and increased Cli, especially in the context of hypoxic–ischemic encephalopathy. The authors investigated the effects of seizure-induced changes in cellular volume by measuring Cli in CA1 neurons in organotypic slice cultures, in which spontaneous ictal-like discharges occur. They found that prolonged seizures in this system increase both neuronal volume and Cli, known to be mediated by NKCC1 and reduced by its antagonist, bumetanide (6). Further relevance to epilepsy would be suggested by an increase in fixed Ao and Ai over development, accompanied by maturational change in the chloride gradient, which will impact GA-BA-A signaling and anticonvulsant efficacy. Gliosis and other changes in the extracellular milieu could also affect GABA-A signaling and contribute to medical intractability in chronic epileptic foci. Finally, damage to the extracellular environment itself might induce changes in the chloride gradient that could compromise GABA-A signaling after injury, leading to acute seizures. Although these comments are speculative currently, the specific type and concentration of Ao may offer a potential new target for seizure control.
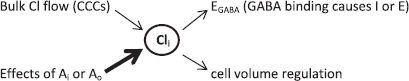
The centrality of Cli in cellular excitability is summarized in the schematic. Both CCCs and fixed anions affect Cl homeostasis, but experiments described here suggest that fixed anions play a prominent role in determining Cli (indicated by the thicker arrow). Cli then directly influences the polarity of GABA action (inhibition I or excitation E) and also regulates cell volume. If a cell is viewed as a container with a finite volume, its capacity to change Cli will be constrained by fixed intracellular anions that limit the influx and efflux of Cl and accompanying water and cations via CCCs. Similarly, the extracellular matrix is not an infinite space but rather a spatially limited, gelatinous admixture of polyanionic biopolymers that can constrain ion mobility (7).
It is clear that Cl ions are not simply passive bystanders in cellular excitability, as once thought. In the 1999 science fiction film The Matrix, the protagonist (“The One”) is confined to a simulated reality or “matrix” by sentient computers that obtain their energy from captive humans. The One is prophesied to set humans free. Similarly, in the brain, impermeant extracellular anions act to constrain chloride ions to the extra-cellular matrix, thus exerting a powerful influence on neuronal excitability. Whether fixed anions are “The One” explanation for GABA action awaits additional validation.