
Abstract
Genomic islands interspersed in the chromosome of P. aeruginosa led to inter- and intraclonal diversity. Recently, a particular clone of P. aeruginosa called clone C was isolated from cystic fibrosis (CF) patients, clinical and non-clinical habitats throughout Europe and in Canada. P. aeruginosa clone C strains harbour up to several hundred acquired genes involved in the adaptation of bacteria to diverse niches. Two genes (hp25 and hp18) from one of the hypervariable regions in the chromosome of clone C strains were highly expressed under standard culture conditions as well as under conditions that mimicked CF sputum environment. Protein sequence analysis revealed that Hp25 and Hp18 belonged to small heat shock protein (sHSP) family. Hp25 protein possessed α-crystallin domain, which is a conserved region among heat shock proteins involved in diverse functions. Sequence homology search revealed that in the Methylobacillus flagellatus genome both genes were situated close to each other and the hp25 gene is found among a few other members of Proteobacteria. Expression of hp25 and hp18 by inter- and intraclonal strains of P. aeruginosa suggested that both genes were present in the stable part of the hypervariable region at the toxR locus and might play a role in their adaptation to diverse niches including the CF lung environment.
Introduction
Chronic colonization of P. aeruginosa in the lung of cystic fibrosis (CF) patients and the associated inflammation results in morbidity and mortality.1,2 The worldwide occurrence of a predominant clone of P. aeruginosa, called clone C, in CF patients, ear infection, peritoneal dialysis fluid, clinical environment, sediment, and water from Germany, Belgium, France, England and Canada has been revealed.3,4 Clone C strains of P. aeruginosa were shown to possess a range of DNA insertions acquired by horizontal gene transfer 5 when compared to the laboratory-adapted reference strain PAO. Sequence analysis of inserted DNA revealed the presence of genes involved in novel functions, which equipped P. aeruginosa to inhabit diverse niches.6–8 Microorganisms inhabiting unusual niches respond to a wide range of environmental stresses like temperature, pH, oxidants, host factors, etc by inducing the synthesis of stress proteins including heat shock proteins (HSPs). 9 In general, HSPs are classified according to their molecular weights of 100, 90, 70, 60 and 40 kilo daltons. HSPs that have low molecular weight in the range of 12 to 43 kDa are grouped together as small HSPs (sHSPs). 10 Small HSPs, which lack ATP-binding sites interact with partially denatured proteins, prevent their aggregation and under certain conditions transfer partially-denatured substrates to chaperones possessing ATPase activity.11–13 In some cases, the presence of common epitopes in host proteins and microbial HSPs may elicit autoimmune responses thereby inducing apoptotic/necrotic effect that ultimately leads to tissue destruction. 14 Proteome analysis by 2D-GE has been a powerful tool in the indentification of proteins involved in the colonization of P. aeruginosa in the CF lung.15,16 In this study, two-dimensionally separated proteins extracted from P. aeruginosa clonal variants and the reference strain PAO were compared to identify the presence of novel proteins not synthesized by the latter strain. At least two novel proteins found in the proteome of all clone C strains showed higher similarity to HSPs, which might play a significant role in the adaptation of P. aeruginosa towards biotic and/or abiotic niches.
Experimental Procedures
Bacterial strains and growth conditions
Pseudomonas aeruginosa strains 4 were pre-cultured in trypticase soy broth (TSB) (1% Tryptone, 0.5% sodium chloride) overnight at 37 °C with shaking. The pre-culture was used to prepare a lawn culture on trypticase soy agar (TSA) (TSB containing 1.5% agar) and incubated for 16 h at 37 °C. Artificial sputum medium 17 was inoculated with P. aeruginosa strain C and incubated for 16 h at 37 °C with shaking.
Preparation and purification of whole-cell proteins
Whole-cell proteins from bacteria were extracted as described. 18 Briefly, bacteria were scraped from the plate or harvested from liquid culture, washed twice with 0.01 M PBS, pH 7.5 and centrifuged (6,000 g for 10 min. at 4 °C) to obtain the bacterial pellet. Whole-cell proteins were extracted as described. 19 1 ml volume of cell pellet was thoroughly dispersed in 2 ml of solubilization buffer (7 M Urea, 2 M thiourea, 4% (wt/vol) CHAPS, 30 mM DTT, 2 μM leupeptin, 1 mM PefaBloc SC, 20 μl 1 M Tris (pH 9.5), 0.5% (wt/vol) Pharmalyte (pH 3–10)) and lysed step-wise using a Branson probe sonicator (Danbury, USA) on ice (40% duty cycle; Microtip limit 5) for 30 s. Further sonications were carried out for 3 × 20 s with an interval of 30 s which ensured efficient lysis. The sample mixture was then centrifuged at 50,000 g for 30 min at 10 °C. The supernatant was separated carefully from the pellet and used immediately for phenol-acetone purification of proteins. 18 For the proteome analysis, 600 μg of the whole-cell protein was used for iso-electric focussing across the linear pH range 4–7 after quantification using the Bio Rad protein assay kit (Bio-Rad).
Isolation and purification of membrane proteins
Membrane proteins were isolated from P. aeruginosa strain C and PAO as described previously. 18 400 μg of protein was separated by 2-DE across the linear pH range 3–10.
Two-dimentional gel electrophoresis
Isoelectric focussing was carried out using IPGphor (Amersham Pharmacia) at 20 °C. IPG strips were rehydrated for 4 h and the following scheme was used for focussing: 30 V constant for 10 h, 150 V gradient for 2 h, 300 V gradient for 2 h, 600 V gradient for 2 h, 1500 V gradient for 4 h, 3500 V gradient for 8 h, 8000 V gradient for 3 h, and a final 8000 V constant for a total of 250 KVh. IPG strips were reduced (6 M Urea, 30% glycerol, 1% (wt/vol) DTT, 2% (wt/vol) SDS in 0.05 M Tris-Cl (pH 8.8)) for 12 min and free SH-groups were blocked with iodoacetamide (6 M Urea, 30% glycerol, 260 mM iodoacetamide, 2% (wt/vol) SDS in 0.05 M Tris-Cl (pH 8.8) for 12 min. Then the strips were embedded on top (using 0.5% agarose in running buffer colored with bromophenol blue) of a 12%–15% pore-gradient SDS-PAGE and run in 24 mM Tris, 0.2 M glycine, 0.1% SDS at 6 V/cm at 10 °C until the bromophenolblue dye front reached the end of the gel. The gels were fixed in 40% ethanol and 10% acetic acid mixture for at least 4 h to overnight, successively stained overnight with Coomassie brilliant blue G250 (Biomol, Germany) as described by Neuhoff 20 and destained overnight in distilled water. After scanning (Umax Powerlook III) the gels, protein spots, which were uniquely produced by P. aeruginosa non-PAO isolates were excised and processed for mass spectrometry.
Protein identification and characterization
Preparation of protein spots for mass spectrometry was carried out as described. 21 Briefly, a protein spot was excised from the gel and chopped into tiny pieces. After washing with deionised water the pieces were shrunk with 50 μl acetonitrile. The supernatant was discarded and the gel pieces were reswollen in 50 μl of 10 mM dithiothreitol in 0.1 M NH4HCO3 to reduce the disulfide bonds of the proteins. After incubation at 56 °C for 30 min, 50 μl of acetonitrile was added to shrink gel pieces. The supernatant was removed and the gel pieces were reswollen in 50 μl of 55 mM iodoacetamide in 0.1 M NH4HCO3. After incubation for 20 min in the dark at room temperature, the supernatant was removed completely. Gel pieces were shrunk using acetonitrile as before, dried briefly, reswollen in trypsin solution (2 ng/μl trypsin in 0.05 M NH4HCO3) and incubated overnight at 37 °C for digestion. After incubation 10–15 μl 25 mM NH4HCO3 was added to the gel pieces and incubated at 37 °C for 15 min with shaking. Then 50 μl acetonitrile was added to the previous mixture and incubated at 37 °C for 15 min with shaking. Supernatent was collected and stored. To the gel pieces 40–50 μl 5% formic acid was added and incubated at 37 °C for 15 min. with shaking. 50 μl of acetonitrile was added to the previous mixture and incubated at 37 °C for 15 min. with shaking. The gel particles were spun down to collect the supernatant, pooled with previously collected supernatant and dried completely in a Speedvac (Eppendorf). Dried peptides were recovered in 10 μl of 0.5% formic acid in 5.0% methanol and desalted using C18 ZipTip (Millipore). Elution of an aliquot of peptides on MALDI-TOF target was done with an equal volume of saturated solution of alpha-cyano-4-hydroxycinnamic acid in 0.5% formic acid/ 65% methanol mixture. PMF maps of trypsin-digested peptides generated by MALDI-TOF Reflex II (Bruker-Franzen-Analytik) mass spectrometer were used to identify the protein using MS-Fit of Protein Prospector package (http://falcon.ludwig.ucl.ac.uk/ucsfhtml3.2/msfit.htm). The peptide sequence-tags were generated by ESI MS/MS Q-Tof-2 (Micromass) and the N-terminal sequencing (Procise Protein Sequencer) was carried out with protein spots excised from a polyvinylidene fluorideblotted 2D gel.
Pulsed field gel electrophoresis
For pulsed field gel electrophoresis (PFGE) analysis, high molecular weight DNA was prepared and separated as described previously.5,22 Ethidium bromide stained gels were visualized using a CCD camera system (Fujifilm Image Reader LAS-1000). Transfer of DNA from PFGE gels onto Nylon membranes (Amersham Pharmacia) was carried out as described.22,23 Southern hybridisation was carried out with probes, which detect Uk-282. Probes were created by PCR and labelled with digoxygenin using (CSPD, Anti-digoxygenin-AP-Fab fragments and DIG-high prime, Roche) as described previously. 22 The hybridisation procedure and immunological detection of probe signals were performed according to previous protocols. 22
PCR and sequencing
Polymerase chain reaction was carried out using standard protocols. Primers Hp25(U): 5'-TCAGCC GTTGATGGGGAT-3' and Hp25(L): 5'-GTGAGC GAGAGTCTTCAATTTTT-3' were used to amplify a 609 bp DNA fragment from the genome of P. aeruginosa strain C and SG17M. The amplicon was sequenced using BigDye (Applied Biosystems) terminator. The t-RNA-specific DNA insertions were identified by a method described previously. 3
Genbank accession number
Hp25 sequences from P. aeruginosa C and SG17M were submitted to the Genbank and the accession numbers are DQ305490 and DQ305491 respectively.
Results
Proteomics and identification of novel proteins
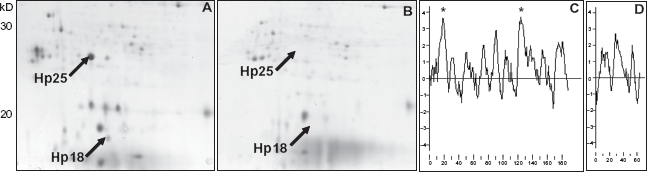
Whole-cell proteins from P. aeruginosa isolates collected from diverse niches were separated in the first dimension based on their isoelectric pH and in the second dimension based on their molecular weight. Two-dimensional protein expression profiles of different P. aeruginosa isolates showed a well-conserved pattern (data not shown) irrespective of their genotype, which is interspersed with numerous genomic islands. 4 Comparison of two-dimensional protein expression profile of the reference strain PAO 18 revealed unique protein spots found only in the proteome of P. aeruginosa non-PAO isolates from different niches (Figs. 1, 2a, 3 and data not shown). Two unique protein spots of relative molecular weight 25 KDa (Hp25) and 18 KDa (Hp18) (Fig. 1) were found in the proteome of all P. aeruginosa isolates (Fig. 3 and data not shown) other than PAO used in this study. Hp25 and Hp18 proteins were among the few highly expressed proteins in the proteome of P. aeruginosa strain C grown in the artificial sputum medium (ASM) (Fig. 1b). 17 Those protein spots were excised from the gels and analysed further for sequence information using mass spectrometry and Edman sequencing.

Whole-cell protein expression profile of P. aeruginosa strain C grown in artificial sputum medium (ASM). Arrows mark the position of novel sHSPs, Hp25 and Hp18. Boxed region is elaborated in Figure 3.
Membrane sub-proteome of P. aeruginosa
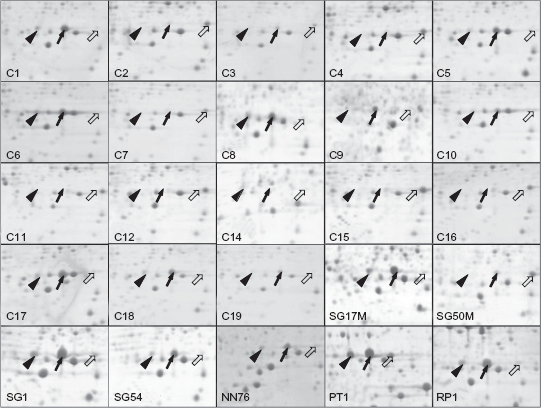
Protein isoforms of Hp25 expressed by intra- and inter-clonal variants of P. aeruginosa isolated from diverse niches. Block arrow indicates native form (pI = 5.54), line arrow and arrowhead indicate post-translationally modified isoforms (pI = 5.34, 5.19). Each window represents the area in a 2D gel that is similar to boxed region in Figure 1.
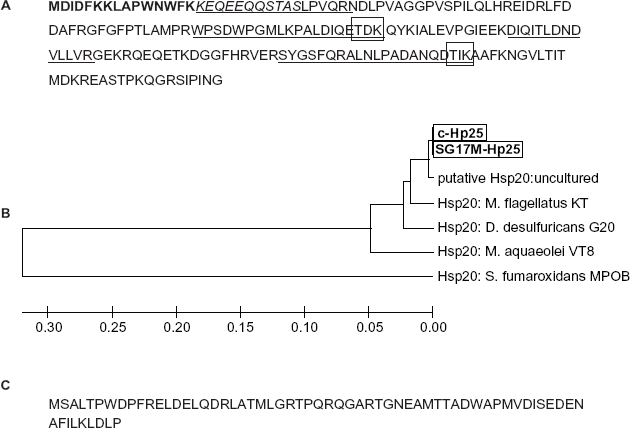
Peptide mass fingerprints of Hp25 and Hp18 obtained using MALDI-TOF were not sufficient for the identification of those proteins. Therefore, peptide sequence tags (Fig. 4a) were obtained using ESI-MS/MS in addition to N-terminal sequencing by Edman degradation. The first 18 amino acids of N-terminal of Hp25 and Hp18 were used to identify homologous proteins in the non-redundant protein database 24 (Figs. 4a, c).
Three isoforms of Hp25 protein were found in the two-dimensionally separated whole cell proteins from non-PAO P. aeruginosa isolates (Fig. 3). Each P. aeruginosa strain showed the up regulation of one of the isoforms only (Fig. 3). In silico analysis showed the presence of two possible threonine phosphorylation-sites in the Hp25 protein (Fig. 4a). Most of the intra- and inter-clonal P. aeruginosa isolates expressed one form of Hp25 (Fig. 3), whereas the expression of other isoforms was variable.
Localization of sHSPs
Whole-cell proteome of P. aeruginosa 18 isolated from diverse niches revealed the presence of novel proteins Hp25 and Hp18 when compared to the proteome of the strain PAO. The membrane sub-proteome of the strain C showed protein spots that corresponded to the novel sHSP proteins as analysed by Q-Tof (Fig. 2a). The hydrophobicity plot (Figs. 2c and d) using Kyte and Doolittle algorithm 25 showed that the Hp25 and Hp18 proteins had hydrophobic regions, which might associate these proteins to the membrane for membrane-associated chaperone activity. Motif-scan (www.expasy.ch) on Hp25 showed the presence of a typical involucrin-repeat like sequence (KEQEEQQ), which is one of the indicators of the membrane associated nature of a protein. Involucrin is present in keratinocytes of epidermis and other stratified squamous epithelia. Involucrin first appears in the cell cytosol, but ultimately becomes cross-linked to membrane proteins by transglutaminase thus helping in the formation of an insoluble envelope beneath the plasma membrane. 26
Homologues of Hp25
Homology search for Hp25 protein using BLAST 24 (1) against bacterial non-redundant protein database provided hits against the genomes of proteobacteriaeae and one hit was from an uncultured bacterial DNA derived from a pooled blood samples of malaria patients from the northern part of India. Multiple protein alignment using CLUSTALW (www.ebi.ac.uk) (Fig. 4) showed that the Hp25 was 99% identical to the putative sHSP from the metagenomic study from the pooled blood sample from malarial patients. Hp25 contained a highly conserved α-crystallin domain at the C-terminus. Alpha-crystallin-domain sHSP superfamily proteins are characterized by the presence of homologous sequence of 80–100 amino acid residues in prokaryotes and eukaryotes involved in diverse functions related to stress-associated survival. 27 Recently sequenced proteobacteria Methylobacillus flagellatus contained two closely-situated genes ZP_00564196 and ZP_00564200 homologous to Hp25 and Hp18 respectively. The genome of Desulfovibrio desulfuricans contained a gene (YP_388230) homologous to Hp25 only. However, the closest homologue was a putative HSP20 (AJ488197) from the gene amplified from the metagenomic study. These observations provide evidence for horizontal gene transfer of Hp25 and Hp18 genes among the members of proteobacteria.
Primer Design and Gene Amplification
The N-terminal sequence of Hp25 showed 100% sequence identity to ZP_00564196 in M. flagellatus. Therefore, the DNA sequence of ZP_00564196, which was homologous to hp25, was taken as reference and primers were designed to amplify the gene from P. aeruginosa isolates. Hp25 gene was amplified from P. aeruginosa strain C, which was isolated during the onset of the colonization process in the CF lung and the strain SG17M, which was isolated from water. 22 PCR amplification of flanking regions of the Hp25 gene in P. aeruginosa strain C and SG17M were similar to that of M. flagellatus (data not shown).
Location of hp25 in the Genome
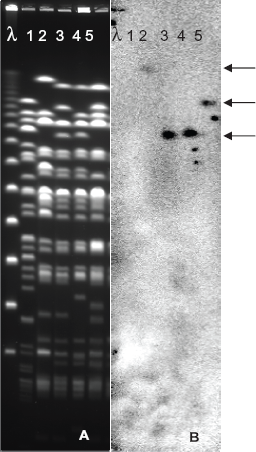
Gene probe was prepared using the PCR amplified hp25 from P. aeruginosa strain C in order to find the location of the gene in the genome of prototype clone C strains. The high molecular weight DNA from P. aeruginosa PAO and the clone C reference strains 3 was digested with the rare-cutting SpeI and separated by pulse-field gel electrophoresis. The macro-restricted DNA fragments were transferred to a nylon membrane and hybridised with the hp25 gene probe. Southern analysis using the probe showed that the genomic island inserted at a non-tRNA locus adjacent to toxR gene in the SpB macro-restriction fragment in the P. aeruginosa strain C harboured hp25 gene (Fig. 5). The gene hp25 representative of that genomic island was found to be present in other clone C strains as well as the European-wide novel clone C strains, 3 which had SpB fragment of variable size than that of strain C. Deletions ranging from 86 to 214 kb within the SpB fragment were reported 28 in several clone C strains. This finding correlates with the various sizes of locus corresponding to SpB fragment among P. aeruginosa isolates originated from various niches (Fig. 5).
Discussion
In this study, the comparison of two-dimensionally separated whole-cell proteins from P. aeruginosa clonal variants revealed the expression of novel proteins that are homologous to protein involved in heat-shock response in bacteria. P. aeruginosa, though a versatile opportunist, exhibits a clonal population structure with a predominant clone, for example clone C, in diverse niches. 4 The genome of P. aeruginosa clone C strains was characterized by the presence of numerous DNA insertions carrying a number of genes involved in niche-specific adaptation. 29 Here it was found that two novel heat-shock proteins (Hp25 and Hp 18) were expressed by all P. aeruginosa clone C strains and a few strains from a distant clone. The expression of Hp25 (α-crystallin domain protein) and Hp18 under standard culture conditions as well as in the artificial sputum-like environment indicated that the proteins might be necessary as a molecular chaperone in the adaptability of non-PAO P. aeruginosa strains inhabiting diverse niches including the CF lung. Isolation of these novel heat-shock proteins in membrane fractions suggested that they might be associated with membrane complexes involved in the stabilization of the bacterial membrane which is in close proximity to the environment. Hsp 16.3 of Mycobacterium tuberculosis was found located on the outer cell wall serving as a major peripheral membrane protein 30 still maintaining the chaperone activity. 31 Membrane association of mammalian α-crystallins is a widespread feature of this superfamily. 32
The sequence identity of Hp25 and Hp18 to the proteins found among Proteobacteria indicated the mobile nature of the DNA block containing the respective genes. The sequence information of pKLC102, a reversibly integrated mega-plasmid in P. aeruginosa clone C strains, showed DNA-sharing with the members of Proteobacteria. 33 The expression of hp25 and hp18 by the intra- and inter-clonal variants of P. aeruignosa reveals its important biological role irrespective of its presence in the hypervariable region of the genome. This suggests that a part of the hypervariable region, located near the toxR gene, consisting of hp25 and hp18 genes is stably maintained among the members of Proteobacteria including P. aeruginosa clones inhabiting diverse niches. The presence of Hp25 in the vicinity of transposase gene in the D. desulfuricans genome and its location in the hypervariable region indicate the recent occurrence of horizontal DNA transfer in these organisms. The consistence in the occurrence and pattern of Hp25 isoforms in the proteomes of all P. aeruginosa non-PAO strains used in this study indicates a possible necessary post-translational modification of this protein. Reversible phosphorylation, translocation and dissociation of sHSPs were reported from prokaryotes and eukaryotes.34,35 This study is the first report on the expression and distribution of identical α-crystallin type heat-shock protein among the members of Proteobacteria. Further experiments on the specific functional properties of Hp25 and Hp18 in P. aeruginosa clones might provide insights about the CF lung-specific adaptation of this organism.
Disclosure
The author reports no conflicts of interest.