
Abstract
Essential oils (EOs) from plant extracts have been reported to have an antibacterial activity against gram-negative and gram-positive bacteria. Several of the gram-positive bacteria are involved in ruminal biohydrogenation of fatty acids (FAs), thus suggesting that feeding EOs could lower biohydrogenation of FA because of a decrease in the number of bacteria involved in that process. As a result, milk FA profiles are expected to be modified. In addition, monensin was approved as an antibiotic to be fed in dairy cattle, and it was reported that dairy cows supplemented with monensin produced milk containing higher concentration of 18:1 t10 and 18:1 t11. The objective of this study was to evaluate the effects of two EOs (garlic and juniper berry oils) and monensin on FA profiles of milk fat. Four ruminally fistulated Holstein dairy cows were used in a 4 x 4 Latin square experiment. Cows were fed for ad libitum intake a total mixed ration without supplementation (control), or supplemented with monensin (330 mg/head per day), garlic oil (5 g/head per day), or juniper berry oil (2 g/head per day). The FA composition of saturated, monounsaturated, or polyunsaturated was not affected by supplementation of EO and monensin. However, proportion of conjugated linoleic acid trans 10, cis 12 (CLA t10, c12) was higher (P < 0.05) for cows fed EO or monensin than for control cows. Supplementation of monensin increased (P < 0.05) the proportion of total trans FA compared with the control. These results indicate that supplementation of the dairy cow diet with garlic or juniper berry EO or monensin had the potential to increase the proportion of CLA t10, c12 in milk fat with minimal overall effects on FA of milk fat. The results also confirm the increase of 18:1 t10 in milk fat by feeding monensin to dairy cows.
Introduction
Antimicrobial growth promoters such as monensin have been used extensively in ruminant production to modulate competition among different microbial populations for improving the efficiency of energy and protein utilization in the rumen. Monensin is a carboxylic polyether ionophore antibiotic that is produced by fermentation of Streptomyces cinnamonensis. 1 Monensin has been approved for use in lactating dairy cows and feeding monensin to lactating dairy cattle has several benefits, which include improved energy metabolism, increased milk production, and reduced incidence of metabolic disorders. 2 However, the use of antibiotics in animal feeds is facing reduced social acceptance because of the potential appearance of residues and resistant strains of bacteria. For this reason, there is increasing interest in evaluating the potential use of natural antimicrobials, such as plant-derived essential oils (EOs) in ruminant nutrition to favorably alter rumen microbial fermentation in order to improve feed efficiency and animal performance.
The EOs have antimicrobial activities against both gram-negative and gram-positive bacteria, a property that has been attributed to the presence of terpenoid and phenolic compounds. 3 A number of in vitro studies have demonstrated that EOs or their compounds have the potential to favorably alter rumen metabolism.4–6 Garlic oil is one of the EOs that was mostly evaluated and demonstrated beneficial effects on rumen fermentation and animal production.5–7 Garlic oil is a complex mix of many different compounds present in the plant or derived from processing, and it has antimicrobial activity against a wide spectrum of bacteria. 8 Several in vitro fermentation trials with rumen fluid reported that garlic oil reduced the concentrations of acetate, branched-chain volatile fatty acid (VFA) and increased the concentrations of propionate and butyrate.5,9 Ramos-Morales et al 10 reported moderate increases in polyunsaturated fatty acids (PUFAs) and C18:1 t10 concentrations in the continuous culture and suggested the potential of garlic oil compounds to modify lipolysis and biohydrogenation in the rumen. Chiquette and Benchaar 11 showed inhibitory effects of garlic (GAR) and juniper berry (JUN) on the production of methane in in vitro rumen fermentation. However, a few studies to date have investigated the effects of either GAR or JUN or their components on digestion, ruminal fermentation, milk composition, or milk production in dairy cows.12,13
The objective of this study was to compare the effects of three antimicrobial agents including monensin, GAR, and JUN on feed intake, ruminal fermentation, feed digestibility, milk production, milk composition, milk fatty acid (FA) profiles, and methane production of lactating dairy cows. The production data were reported elsewhere.7,14
Materials and Methods
Cows and diets
This experiment has been well described by Yang et al. 7 In brief, four ruminally and duodenally fistulated dairy cows (body weight = 705 ± 50 kg, and day in milk = 113 ± 13), at the beginning of the experiment, were used in a 4 x 4 Latin square design. Cows were fed a diet (Table 1) without supplementation (control), or supplemented with monensin (330 mg/cow/day), GAR (5 g/cow/day; Allium sativum; standardized at 1.5% of allicin), or JUN (2 g/cow/day; Juniperus communis standardized at 35% of α-pinene). The EOs were obtained from AXISS France SAS and mixed into the concentrate. The duration of each experimental period was 21 days long with 11 days of adaptation to new treatments and 10 days of sampling and data collection.
Ingredients and chemical composition of the diet (dry matter [DM] basis).
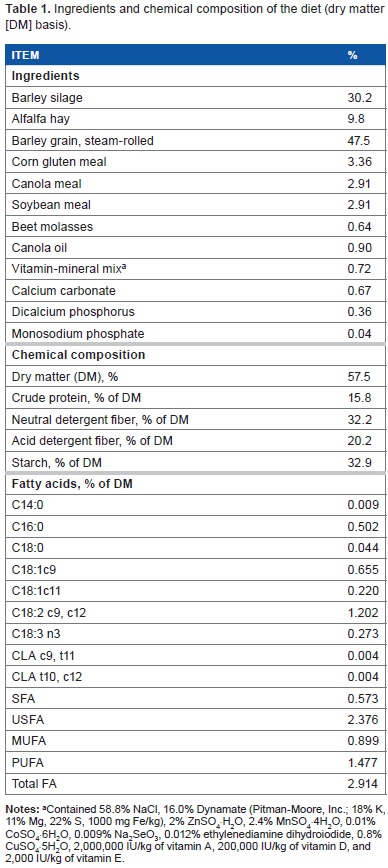
Contained 58.8% NaCl, 16.0% Dynamate (Pitman-Moore, Inc.; 18% K, 11% Mg, 22% S, 1000 mg Fe/kg), 2% ZnSO4·H20, 2.4% MnSO4·4H20, 0.01% CoSO4·6H20, 0.009% Na2SeO3, 0.012% ethylenediamine dihydroiodide, 0.8% CuSO4·5H2O, 2,000,000 IU/kg of vitamin A, 200,000 IU/kg of vitamin D, and 2,000 IU/kg of vitamin E.
Cows were housed in individual tie stalls, fed a total mixed ration ad libitum three times daily at 0600, 1500, and 1800 hours, and milked twice daily at 0700 and 1700 hours. All diets were formulated based on the NRC recommendations 15 to supply sufficient energy and protein for a 700 kg cow to produce 30 kg/day of milk containing 3.5% fat and 3.2% protein. All cows were cared for in accordance with the guidelines established by the Canadian Council on Animal Care (Ottawa, ON, Canada), and all animal-related procedures were approved by the Animal Care Committee of the Agriculture and Agri-Food Canada, Lethbridge, Alberta.
Samples of feeds and refusals were composited by period, then, dried in an oven at 55°C for 48 hours, and ground through a 1-mm diameter screen (Wiley Mill, standard model 4; Arthur Thomas Co.) for subsequent determination of ash, neutral detergent fiber (NDF), acid detergent fiber (ADF), starch, and crude protein (CP). 7 Milk yield was recorded at each milking. During the data collection period, milk was sampled at each milking, preserved with potassium dichromate, and subsequently analyzed for milk composition.
Chemical analyses
Dry matter was determined by oven drying at 55°C for 48 hours (method 930.15). 16 Ash content was determined by combustion at 550°C overnight, and OM content was calculated as 100–-the percentage of ash (method 942.05). 16 The NDF and ADF contents were determined using the methods described by Van Soest et al 17 with α-amylase and sodium sulfite used in the NDF procedure. Starch was determined by enzymatic hydrolysis of α-linked glucose polymers, as described by Rode et al. 18 Total N was determined by flash combustion technique (model 1500; Carlo Erba Instruments). For the analysis of milk FA, methyl esters were prepared by base-catalyzed transmethylation according to Chouinard et al. 19 FAs were analyzed after transesterification with sodium methoxide. FA analyses were conducted using a gas chromatograph (HP 5890A Series II, Hewlett-Packard) equipped with a 100 m CP-Sil 88 capillary column (i.d., 0.25 mm; film thickness, 0.20 µm; Chrompack) and a flame ionization detector, as previously described. 20 Peak area was measured using a Nelson Analytical System 2600 (version 5; PE Nelson). Each peak was identified using pure methyl ester standards (Alltech) on the basis of their retention times. 19 Composition of FA in feed samples was analyzed according to He et al. 21 Lipids in feed were recovered by evaporating the ethyl ether. Nonadecanoic acid (19:0) methyl ester (100 µL, 5.96 mg/mL hexane; Nu-Chek Prep, Inc.) was added to the residues as an internal standard. A combined base/acid methylation procedure using sodium methoxide (0.5 mmol/L in methanol) and boron trifluoride (140 ml/L methanol) was used for methylation. FA methyl esters were quantified using a gas chromatograph (Hewlett Packard GC System 6890) equipped with a flame ionization detector and SP-2560-fused silica capillary column (75 m x 0.18 mm x 0.14 µm; Supelco Inc.).
Statistical analyses
Data were analyzed using the mixed model procedure of SAS (SAS Institute Inc.) to account for effects of period, cow, and treatments. Means were compared using multiple comparison test (LSD), where treatments were considered a fixed effect; period and cow were considered random effects. Differences between each treatment and control were declared significant at P < 0.05 and trends were discussed at P < 0.10.
Results
The data of feed intake, milk yield, and milk composition from this study were reported in the previous manuscript. 7 Intake of dry matter (DM) and milk yield were not affected with GAR, JUN, or monensin supplementation, but milk fat content and yield were lower (P < 0.01) for cows fed monensin than control or EO-fed cows. 7 The milk FA composition of saturated (SFA), monounsaturated (MUFA), or polyunsaturated (PUFA) was affected neither by supplementation of EO nor by feeding monensin (Table 2). Supplementation of GAR or JUN tended (P < 0.07) to increase the proportions of C18:1 t9 and C18:1 t10 in milk fat compared with control cows, and consequently, the proportion of total trans FA tended (P < 0.09) to be greater for cows supplemented with either GAR or JUN than control cows. Compared with control, the cows supplemented with GAR and JUN had also higher (P < 0.04) proportions of conjugated linoleic acid t10, c12 (CLA t10, c12) in milk fat. Supplementation of monensin had higher proportions of C18:1 t6, t8 (P < 0.02), C18:1 t9 (P < 0.01), and C18:1 t10 (P < 0.04), thus greater (P < 0.03) proportion of total trans FA compared with the control cows. The proportion of CLA t10, c12 and the sum of CLA t10, c12 + C18:1 t10 was also higher (P < 0.04) for cows fed monensin than control cows.
Effect of monensin and plant-derived EO supplementation on milk FA profiles (% of total FA) of lactating dairy cows.

Within a row with different letters differ (P < 0.05).
Discussion
Information on the effects of feeding GAR and JUN on milk FA profiles of dairy cows is scarce. That supplementation with GAR and JUN had minor effect on milk FAs in the present study, which agrees with the previous studies13,22 in dairy cows supplemented with eugenol EO. Benchaar et al 13 reported no effect of eugenol supplementation on milk FA in dairy cows fed either high-forage (65%) or low-forage (35%, DM basis) diets. In another study using dairy cows, Benchaar et al 22 also observed that only C4:0, 18:1 t11, 18:1 t16, and 18:4 c6,9,12,15 FA proportions were slightly affected by feeding incremental eugenol doses to dairy cows. Similarly, Hristov et al 23 did not observe variations in milk FAs, when supplementing the diet of lactating cows with three dosages of Origanum vulgare leaves. However, our results of the increased CLA t10, c12 in milk with supplementation of GAR or JUN are somewhat consistent with the studies using dairy goats supplemented with GAR.24,25 Kholif et al 24 found that supplementation with GAR, cinnamon oil, or ginger oil increased UFA and CLA c9, t11 and that supplementation with cinnamon oil also increased C18:3 n3. Boutoial et al 26 supplemented extracts of rosemary leaves to lactating goats and observed that PUFA increased as the dosage of extracts increased. It appears that there are more responses of EO supplementation to milk FA profiles in small ruminants than in large ruminants such as dairy cattle. This difference that is related to animal species could be a result of the high feed rumen passage rate of small ruminants, in comparison with that of large ruminants, which could limit the ability of rumen bacteria to complete the biohydrogenation process in the rumen because of high passage rate of feeds out of the rumen. The consistency between the rumen biohydrogenation of FA and milk FA profiles was reported. 25 In fact, the limited effect of EO addition on milk FA profiles is consistent with the lack of effect on rumen fermentation in the present study. Rumen fermentation pattern and microbial populations were not affected with GAR or JUN supplementation. 7 Calsamiglia et al 8 described the mode of action of EO in the rumen microbial environment and suggested that a change in bacterial growth rates results in changes in the proportion of rumen bacterial populations. Different bacterial populations may result in different proportions of rumen FA and thus difference in milk FA.
Milk FA composition is of great interest for human nutrition, and several strategies have been proposed to improve FA composition, decreasing SFA and increasing UFA. 27 The addition of EO in ruminant diets can be an alternative strategy to improve milk FA profiles. Morsy et al reported that supplying clove EO decreased concentrations of 18:0 and 18:1 c9 and increased PUFA in milk fat of dairy goats. 28 Nevertheless, in the present study, the effects of feeding GAR or JUN on milk FA profiles appeared to be minimal. The lack of effects on milk FA profile is consistent with previous in vivo studies with dairy cows using different sources of EO compounds.12,13 However, contrary to monensin, the higher milk CLA t10, c12 with supplementation of EO did not decrease milk fat content and fat yield, which suggest the advantage of supplementing GAR or JUN over monensin to avoid an induction of milk fat depression.
Monensin was chosen as a positive control because of its approval as a feed additive in dairy cattle and its several benefits, including improved energy metabolism, increased milk production, and reduced incidence of metabolic disorders. 2 However, similar to unsaturated oils, monensin has also been associated with milk fat depression by inhibiting UFA biohydrogenation within the rumen. 29 The milk fat content and yield were decreased by 15% and 13%, respectively, by monensin supplementation versus control cows in this study. 7 Supplementation of monensin increased significantly the concentration of C18:1 t10, which is considered as a potent inhibitor of milk fat synthesis. 30 Monensin supplementation did not affect proportions of the total short-chain and medium-chain SFA in the present study, which agrees with the study by Odongo et al 31 in dairy cow fed 33 mg monensin/kg dietary DM. The increased C18:1 t6-8, C18:1 t9, C18:1 t10, and CLA t10, c12 with monensin addition are also consistent with the studies by AlZahal et al 32 and He et al, 33 but contrast to the observation of Odongo et al. 31 The inconsistent effects of monensin on milk FA proportion among studies could be due to various factors including diet composition, dose and delivery of monensin, and stage of lactation of dairy cows. For example, the dose of monensin (mg/kg DM intake) was about 16.5 and 17.5, respectively, in the present study and the study of He et al 33 which were lower than the dose of menensin (24.0) used in the study of Odongo et al. 31 Similarly, in the study by Benchaar et al 30 in which monensin was fed at a dose of 16 mg/kg, monensin did not affect short- and medium-chain FA in milk, whereas the concentrations of 18:1 t10 and 18:1 t11 were increased. The increased proportions of C18:1 t6-8, C18:1 t10, and CLA t10, c12 with monensin are also consistent with the decrease of milk fat yield in this study, as previously reported. 7 Indeed, CLA t10, c12 is well known for its milk fat depression capacity, C18:1 t10 is potentially antilipogenic, 30 and trans-6 to 8 C18:1 is negatively correlated with milk fat content. 33 The monensin supplementation did not change the proportions of rumen individual VFA, 7 and thus, it explains the no differences in the proportions of odd- and branched-chain FA, which are synthesized by chain elongation of VFA in the rumen. 34
Conclusion
Addition of garlic oil (5 g/cow/day) or juniper berry oil (2 g/cow/day) in the diet of dairy cow increased the proportion of CLA t10, c12 in milk fat. However, overall effects of EO on FA in milk fat were minimal. Supplementing monensin (330 mg/cow/day) in the diet of dairy cows increased the proportion of C18:1 t6-8 FA and the total trans FA in milk. These results suggest the advantage of supplementing GAR or JUN over monensin to not significantly increase trans FA and to avoid an induction of a milk fat depression, although the milk CLA t10, c12 was similar between EO and monensin.
Author Contributions
Conceived and designed the experiments, performed the experiments, analyzed the data, and wrote the paper: WZY and MLH. Agreed with manuscript results and conclusions: WZY and MLH. All the authors reviewed and approved the final manuscript.
Footnotes
Acknowledgments
The authors are thankful to K. Andrews and A. Furtado for their assistance in sample collection and laboratory analyses as well as to the technical staff of Lethbridge Research Centre, Dairy Unit for their continuous help with the care to the cows and collection of milk samples.