
Abstract
The chytrid fungus Batrachochytrium dendrobatidis is a significant cause of the worldwide decline in amphibian populations; however, various amphibian species are capable of coexisting with B. dendrobatidis. Among them are boreal toads (Anaxyrus (Bufo) boreas boreas) located in Grand Teton National Park (GTNP) in Wyoming, USA. The purpose of this study was to identify cultivable bacterial isolates from the skin microbiota of boreal toads from GTNP and determine if they were capable of inhibiting B. dendrobatidis in vitro, and therefore might be a factor in the toad's coexistence with this pathogen. Isolates from 6 of 21 genera tested were found to inhibit the growth of B. dendrobatidis. These bacteria represent diverse lineages such as the Gammaproteobacteria, the Betaproteobacteria, and the Bacteroidetes/Chlorobium groups. We propose that these bacteria compete via microbial antagonism with B. dendrobatidis.
Introduction
Approximately one-third of the amphibian species around the world face extinction, and chytridiomycosis is believed to be one of the factors responsible for this threat.1–5 This disease is caused by the fungal pathogen Batrachochytrium dendrobatidis, which infects keratinized epithelial cells of amphibians. 6 The fungus causes damage to these cells, which is believed to disrupt osmotic regulation as well as electrolyte levels, eventually leading to the death of the host.7–9 This disruption is caused by the thickening of the keratinized cells, specifically around the drink patch of the amphibian. A thickening of the cells ultimately leads to poor gas exchange, and the disruption of electrolytes can cause deadly dehydration. It is also possible that B. dendrobatidis excretes a toxin before or after infection, contributing to amphibian death. 10
Infection by B. dendrobatidis also appears to be enhanced by the chytrid's ability to inhibit the proliferation of lymphocytes by production of a toxin, which may also prevent the normal function of lymphocytes. Zoospores of B. dendrobatidis become inhibitory to lymphocytes after initial infection and encystment of the zoospore. The matured zoospore can induce apoptosis of lymphocytes. This form of inhibition and loss of function has been seen in other pathogenic fungi, such as Cryptococcus neoformans, Aspergillus fumigatus, and Paracoccidioides brasiliensis. The ability to induce apoptosis in these species has been tied to cell wall components, and it is likely that the inhibitory factor is similar in action in B. dendrobatidis. This may explain why encysted zoospores are inhibitory, as free-swimming zoospores do not contain cell walls. The apoptotic effect is not only tied to lymphoid cells but can also have a broader cytotoxic effect and thus could be responsible for some of the other symptoms of chytridiomycosis, such as disruption of the skin (thickening of the cells preventing gas exchange), lethargy, and loss of righting reflex. 11
Despite the frequent reporting of mortality associated with this pathogen, there are populations of animals infected with B. dendrobatidis that appear to co-exist with the fungus.2,12–14 For example, B. dendrobatidis has been identified on boreal toads, Anaxyrus (Bufo) boreas boreas, in Grand Teton National Park (GTNP) in Wyoming, USA and no acute mass mortalities have been observed in this region.15,16 In contrast, in Colorado the same species of toad has experienced mortality from this pathogen. 17 Genetic analysis studies have shown that different isolates of B. dendrobatidis from areas around the world associated with (and without) mortality events have low genetic diversity within the given populations.2,18–21 More recent studies have indicated that there is a high genetic diversity in B. dendrobatidis when comparing populations from different areas of the world. 22 It is thought that the global decline may be tied to the introduction of different, and more virulent, strains to a new environment, as has been documented with the transport of the American bullfrog. 22
Recent studies conducted in our laboratory have found that B. dendrobatidis isolates from Colorado and Wyoming cause similar mortality in boreal toads from these different locations under laboratory conditions, 16 suggesting that factors other than the host and pathogen may play a significant role in the manifestation of chytridiomycosis within these areas. These other factors may have effects on the pathogen, the amphibian, and/or the skin microbiota of the amphibians.
It has been observed that cultures of B. dendrobatidis are highly susceptible to a range of antifungal peptides and/or antifungal compounds produced by amphibians, many with potential to be used in drug development, such as Manganin I, Bradykinin, Esculentin-1, and Temporin A.5,23–26 Harris et al 27 surveyed Rana muscosa from two populations and found antimicrobial skin peptides as only one factor in the resistance of R. mucosa that inhibited B. dendrobatidis. Harris et al 4 suggest that the motile chemotactic zoospores reach the surface of the skin and attach quickly, but bacteria on the skin release antifungal substances, which inhibit growth of this fungus.
It is also known that the normal microbiota of a host can offer resistance to disease, through microbial antagonism or other mechanisms.28–31 Bacteria in biofilms on the surfaces of organisms, such as amphibians and coral, may contain a higher proportion of antibiotic-producing bacteria than some in the surrounding environment.32–34 Additionally, epiphytic bacteria can produce antimicrobial secondary metabolites, inhibiting the settlement of potential competitors, which can be beneficial to the host.33,35,36 Examples of antimicrobials that inhibit B. dendrobatidis and are produced by microbial flora are 2,4-diacetylploroglucinol and violacein.27,33,37 This suggests that normal bacterial flora may inhibit infection and/or colonization of the skin by this pathogen. 29
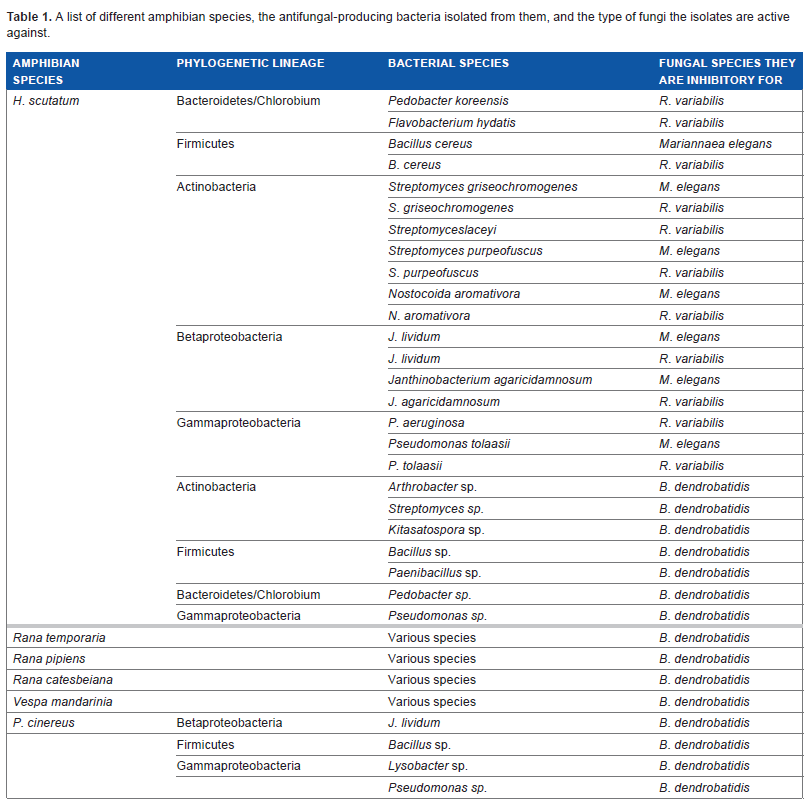
Harris et al 29 identified several bacteria grown in vitro from the skin of two salamander species (Plethodon cinereus and Hemidactylium scutatum), which inhibited the growth of B. dendrobatidis. These and other amphibian species, and the antifungal-producing bacteria found on them, can be seen in Table 1. These substances may be fungicidal (leading to the death of the fungi), fungistatic (inhibiting the growth of the fungi), or fungilitic (leading to the death of the fungi by cell lysis). To further show that it was the natural flora of the amphibians that was capable of inhibiting the growth of B. dendrobatidis, members of the species R. muscosa (a frog susceptible to B. dendrobatidis infection) were inoculated with a culture of Janthinobacterium lividum and growth of B. dendrobatidis was monitored. Harris et al noted that the addition of J. lividum prevented morbidity and mortality in R. muscosa. 27
A list of different amphibian species, the antifungal-producing bacteria isolated from them, and the type of fungi the isolates are active against.
A high percentage of the cells both within and on the exterior of vertebrates are bacteria, and animals often play host to a vast assortment of bacterial species. 38 These bacteria can be viewed as mutualists and are often essential for the survival of the host. Because the skin is continuously being exposed to the environment, the normal flora can have both transient and resident microbial populations. Transient populations inhabit the skin but are unable to grow and multiply (unlike resident populations), and they generally are pathogenic or opportunistic pathogens. 32
A wide variety of bacteria, viruses, and fungi are typically found on the skin, in the digestive tract, and in other tissues of amphibians.39–41 Culp et al 42 compared the natural skin microflora of three different amphibian species via culturing bacterial isolates on agar plates: adult eastern newts (Notophthalmus viridescens), larval bullfrogs (Rana catesbieana), and adult redback salamanders (P. cinereus). Culp et al were able to isolate several phylogenetically distinct bacteria from each of the different amphibians. From N. viridescens, five bacteria were found to naturally occur on the skin: Raoultella terrigena (Gammaproteobacteria), Agrobacterium radiobacter (Alphaproteobacteria), Flavimonas oryzihabitans (Gammaproteobacteria), Chryseomonas luteola (Gammaproteobacteria), and Aeromonas hydrophila (Gammaproteobacteria). Only three bacteria were identified from R. catesbieana: Pseudomonas fluorescens (Gammaproteobacteria), Staphylococcus epidermidis (Firmicutes), and Microbacterium testaceum (Actinobacteria). Four bacterial species were isolated from P. cinereus: Flavobacterium johnsoniae (Bacteroidetes/Chlorobium group), Bacillus cereus (Firmicutes), P. fluorescens (Gammaproteobacteria), and M. testaceum (Actinobacteria). These are vastly different amphibian species that share similar habitats that harbor cutaneous bacterial flora from the same phylogenetic groups, with the exception of A. radiobacter and F. johnsoniae which were from lineages that appeared to be unique to the amphibian they were cultured from. 43 The similarities indicate that the microbial flora of one species may not differ much from another species if environmental conditions are similar.
In another study, approximately 27% of the bacteria isolated from the skin of the salamander Plethodon ventralis exhibited antimicrobial activity against both bacteria and fungi that occurred naturally in the salamander's environment.28,43 This microbial antagonism phenomenon has also been well studied in other systems such as marine free-living and sediment-inhabiting bacteria.33,43
We have identified a diverse group of bacteria on the skin of boreal toads across several collection sites in GTNP, Wyoming, USA, where virulent strains of B. dendrobatidis co-exist with this species of amphibian. Several bacteria were identified to be members of genera known to be inhibitory to B. dendrobatidis, such as Pedobacter and Pseudomonas, as well as genera (Xanthomonas, Stenotrophomonas, Pseudomonas, Flavobacterium, and Pedobacter), which are known to be inhibitory to fungi such as Mutinus elegans and Rhizomucor variabilis. The objective of this study was to identify, among those bacteria isolated from boreal toads in the GTNP, which organisms were capable of inhibiting the growth of B. dendrobatidis in vitro. Identifying bacteria and the compounds they produce that are capable of inhibiting the growth of B. dendrobatidis may lead to the development of new ways to treat this global pathogen.
Methods and Materials
Bacterial isolates
Five adult toads at each of five breeding sites in the Grand Teton area were captured and swabbed at the venter, legs, and feet with a sterile spatula. 16 Latex gloves were worn during sampling procedures. Spatula tips were initially placed in sterile R2 broth in microcentrifuge tubes for transport to the laboratory. During sampling, each toad was contained in sterile containers until capture process was complete to prevent re-sampling of toads, and gloves were changed with each capture and swab. Toads were not rinsed before sampling.
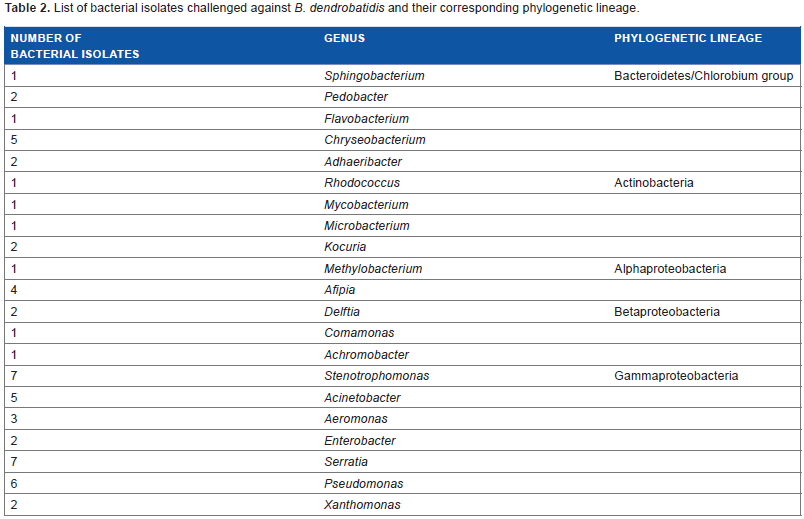
On return to the laboratory, tubes containing the spatula tips were vortexed and 0.1 mL of solution was micro-pipetted onto R2 agar media plates. 44 Bacterial colonies were struck for isolation and on isolation were transferred to R2 broth media. The process of colony selection on R2 agar plates, transfer to R2 broth for growth, and re-streaking for isolation on R2 agar plates was repeated iteratively until axenic cultures were produced. A subset of bacterial isolates identified from the skin of boreal toads from GTNP was challenged against B. dendrobatidis. This subset consisted of 60 bacteria l isolates representing diverse bacterial lineages (Table 2), determined using their SSU (16S) rRNA gene sequences. The bacteria were grown in R2 broth medium until turbidity was seen at room temperature (17–20°C) and then struck out onto R2 agar medium until colonies formed (all isolates were grown on TGhL plates before challenge assays to ensure they were capable of growth). Bacterial colonies were picked in stationary phase (approximately four to–five days old) for challenge assays.
List of bacterial isolates challenged against B. dendrobatidis and their corresponding phylogenetic lineage.
Challenge assays
The B. dendrobatidis strain (isolated from a boreal toad in Black Rock Pond (GTNP, WY)) was used in all challenge assays. B. dendrobatidis was grown in H broth medium (1% tryptone, 0.32% glucose) at room temperature (17–20 °C) for two weeks or until turbidity was seen. Approximately 3 mL of the turbid broth culture was transferred onto hard TGhL agar medium containing 1.6% tryptone, 0.04% gelatin hydrolysate, 0.02% lactose, and 1.5% agar, and distributed across the entire surface. The plates were allowed to dry in a laminar flow hood until they were slightly moist. A single colony of the challenge bacteria (average colony size ~1–3 mm in diameter), grown on R2 medium plates, was struck across the plates in a straight line, and each bacterium was tested in triplicate. The inoculated TGhL plates (100 mm in diameter) were then incubated at room temperature (17–20°C) and were observed every other day up to 10 days for signs of inhibition. They were scored as either inhibitory or not inhibitory based on the presence and size of a zone of inhibition/reduction between the streaked bacterial isolate and the B. dendrobatidis culture. Control plates seeded with only zoospores of B. dendrobatidis were used to verify the viability of the fungus.
Scoring
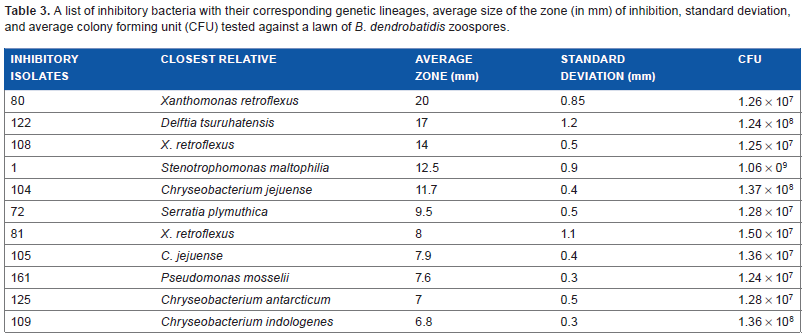
The bacterial isolates that were scored as inhibitory against a lawn of B. dendrobatidis were then re-tested in triplicate to confirm reduction/inhibition. Images were taken using a ProtoCOL HR Model No. 94000 camera and zones of reduction/inhibition using the same protocol described above were measured using the photographs. The average zone of inhibition and standard deviations were calculated for all bacteria with inhibitory properties (Table 3).
A list of inhibitory bacteria with their corresponding genetic lineages, average size of the zone (in mm) of inhibition, standard deviation, and average colony forming unit (CFU) tested against a lawn of B. dendrobatidis zoospores.
Bacterial isolates with a clear zoospore inhibition zone of greater than 12 mm were considered as having strong anti-fungal properties. Isolates with a zoospore clear zone less than 12 mm or with a non-distinct zone of zoospore inhibition were considered to have weak antifungal activity. Bacterial isolates where there was no evidence of inhibition of zoospores (plates covered with active zoospores after 10 days of incubation) were considered not to have antifungal properties.
Results
Control plates showed a complete lawn of B. dendrobatidis across the plates with live swimming zoospores surrounding sporangia (Fig. 1a). Of the 60 bacteria (Table 2) challenged against B. dendrobatidis, 11 bacteria (which are shown in Table 3) had a measurable zone of inhibition (Fig. 1b–1). Microscopic examination of these plates indicated nonmoving zoospores in close proximity to the bacterial species and swimming zoospores outside the zone of inhibition.

Photographs of TGhL plates containing a lawn of B. dendrobatidis taken after 10 days of incubation at room temperature (17–20°C) showing zone of inhibition: (
The bacteria (Isolates 1, 72, 80, 81, 104, 105, 108, 109, 122, 125, and 161) that inhibited growth of B. dendrobatidis are diverse in their bacterial lineages and belong to several genera (Table 3). There are six bacterial isolates (1, 72, 80, 81, 108, and 161) that belong to the Gammaproteobacterial lineage, four bacterial isolates (104, 105, 109, and 125) that belong to the Bacteroidetes/Chlorobium group, and one bacterial isolate (122) that is a member of the Betaproteobacterial lineage. Of these inhibitory bacterial isolates, three had zones of inhibition greater than 12 mm of the plate and are considered strong inhibitors (Table 3) of B. dendrobatidis in the challenge assays. The clearing zones were clear, concise, and evenly distributed around the bacterial streak. Isolates 80 (Fig. 1b) and 108 (Fig. 1d) belong to the genus Xanthomonas, and isolate 122 (Fig. 1c) belongs to the genus Delftia. These bacterial isolates belong to separate phylogenetic lineages; bacterial isolates 80 and 108 are Gammaproteobacteria, whereas bacterial isolate 122 is a Betaproteobacterium.
A total of 5 of the 11 bacteria that inhibited B. dendrobatidis had inhibition zones between 7.6 and 12.5 mm and are considered weak inhibitors (Table 3). These isolates (Fig. 1e–j) belong to three genera (Stenotrophomonas, Chryseobacterium, and Serratia), and are diverse in their phylogenetic lineages, covering the Gammaproteobacterial group (Stenotrophomonas and Serratia) and the Bacteroidetes/Chlorobium group (Chryseobacterium).
The three bacteria tested did not have distinct zones of inhibition but still show evidence of inhibition against B. dendrobatidis consistently (Figs. 1i, k, and l). The sporangia in close proximity to the bacterial streak did not have active zoospores and a distinct clearing zone. These three bacteria all belong to the genus Chryseobacterium and are closely related based on their SSU (16S) rRNA gene sequences.
Discussion
We challenged a subset of bacteria isolated from the skin of B. boreas from GTNP, Wyoming, USA with the pathogen B. dendrobatidis and found 11 of 60 bacteria inhibited the chytrid fungus in vitro. Most of these isolates are considered bacteria of aquatic systems and are likely commensal on the skin of amphibians. Similar findings of commensal bacteria inhibiting B. dendrobatidis have been reported for bacterial isolates from the skin of salamanders.2,4,29,42,45 Given the proportion of bacteria that were found to inhibit B. dendrobatidis, it is possible that under normal environmental conditions, these organisms have a synergistic relationship with each other as well as their host and help reduce the concentration of B. dendrobatidis on the skin of amphibians.
Although we had a high percentage of bacteria that inhibited B. dendrobatidis in vitro, there was significant variation in the level of inhibition among the different isolates. Four of the bacterial isolates produced inhibition zones greater than 12 mm and seven bacterial isolates produced smaller zones of inhibition (Table 3). Isolate 122, belonging to the genus Delftia, was one of the strongest inhibitors of B. dendrobatidis from our collection and the only Betaproteobacterium (Table 3) found to have antifungal properties to B. dendrobatidis. Isolate 122 was the only inhibitory Betaproteobacterium isolated during this study, which was surprising considering that in other similar studies many bacteria with antifungal properties belonged to this lineage.27,38 This genus of bacteria is commonly found to be motile and occurs in aerobic conditions near the surface of the soil. 46 Many members are considered to be highly versatile in their degradation capabilities, for example producing enzymes with chitinase and lysozyme activities. 46
Another relatively strong set of inhibitors in our study belonged to the genus Xanthomonas, a group of organisms that once was a part of the genus Stenotrophomonas. Organisms in the genus Xanthomonas are Gram negative, motile, phytopathogenic pseudomonads. 47 Two of our bacterial isolates (80 and 108) belonging to this group had large distinct clearing zones of inhibition, 20 and 14 mm, respectively, in our assays (Table 3). In addition, Brucker et al 37 reported that the bacterium Lysobacter gummosus, formerly belonging to Xanthomonas, found on the skin of the red-backed salamander, produced the anti-fungal metabolite 2,4-diacetylphloroglucinol, which was effective in inhibiting the growth of B. dendrobatidis.
Interestingly, four of the isolates that mildly inhibited B. dendrobatidis in vitro (non-distinct inhibition zones of less than 12 mm) were from the genus Chryseobacterium. This genus of bacteria is commonly found in freshwater as well as in soil, marine sediment, and plant roots. 48 These four bacteria were found at all five sites tested in GTNP.
Six bacterial isolates belonging to the genus Pseudomonas were evaluated for their ability to inhibit the growth of or kill zoospores of B. dendrobatidis, but only isolate 161 (Table 3) was found to be mildly antifungal. Given that several species of Pseudomonas are known to have antagonistic activity against some pathogenic fungi, 49 we anticipated more of the Pseudomonas isolates to be inhibitory. In fact, some strains of Pseudomonas fluorescens have been used as probiotics against pathogenic bacteria and fungi in aquaculture. 50 In addition, Pseudomonas aeruginosa is known to produce and excrete rhamnolipids, which are compounds that naturally occur as biosurfactants,51,52 which have the ability to lyse fungal zoospores. An unexpected mild inhibitor was found to be a member of the genus Serratia. Isolate 72 had an inhibition zone of 9.5 mm, which is almost 10% to that of the plate. This genus of bacteria is known to be an opportunistic pathogen in humans, and species within this genus are often motile. 53
Overall, 18.3% of the bacterial species isolated from adult toads in five different locations in GTNP were at least partially inhibitory to B. dendrobatidis in vitro, suggesting that these bacteria may help the toads manage fungal infection levels in the environment. Our study design did not permit us to identify the precise antifungal substances that inhibit or kill the zoospores of B. dendrobatidis. Other researchers, however, have identified specific compounds produced by bacteria that have antifungal activity such as violacein4,31 and 2,4-diacetylphloroglucinol. 31 The anti-fungal metabolite violacein is produced by the Betaproteobacterium J. lividum, which is found on the skin of the red-backed salamander.4,31,37
There are other naturally produced antimicrobial compounds found to be inhibitory to B. dendrobatidis, such as chloramphenicol. Bishop et al 54 applied chloramphenicol topically to the skin of adult frogs to remove B. dendrobatidis. After treatment with chloramphenicol, symptoms of chytridiomycosis dissipated and the infected frogs recovered. Although many of the identified natural antifungal products produced by bacteria can inhibit the growth of B. dendrobatidis at high concentrations (ie therapeutic levels), it is not known whether the concentration of these inhibitory products produced by the skin microbiota under natural conditions is sufficient to suppress the growth of B. dendrobatidis. Becker et al, 55 was able to establish that an 18 µM concentration of violacein applied directly to the salamander's skin is able to prevent mortality and morbidity from B. dendrobatidis. Violacein is produced naturally by the bacterium J. lividum. It is plausible that the bacterial community on a toad produces adequate amounts of these compounds to reduce the growth in the immediate area around the skin microbiota; however, further research is required to confirm this hypothesis.
Another research question that remains unanswered is whether boreal toads in different areas have similar anti-fungal microbiota. Amphibians may have selected a group of microbes from their habitat with the functional capacity to resist pathogenic fungi. The boreal toad is a terrestrial amphibian that spends most of its life on land exposing its skin to soil. There are many bacteria found in soil, particularly from the genus Streptomyces and the related genus Actinomyces, which are known to produce antibiotics. 56 Some members of these genera were found in our study, but were not found to be inhibitory to B. dendrobatidis in vitro. However, they may be effective against other pathogens or may only produce antibiotics under certain conditions that were not simulated in this study.
Our results indicate that the resident cutaneous microbiota on B. boreas located in GTNP in Wyoming, USA may have antifungal properties. We propose that this enables boreal toads to coexist with the pathogen B. dendrobatidis. A comparison of the microbial flora of B. boreas from GTNP and microbial flora from populations that are declining in infection by B. dendrobatidis would further support this. Currently studies are underway in evaluating risk factors for chytridiomycosis by considering the microbiota of the animals and the interaction between the animal's microbiota and the environment. Walke et al. 57 conducted a study about how amphibians may become colonized by microbial flora that is inhibitory to B. dendrobatidis. They suggest that the colonization of resident, and inhibitory, bacteria occurs through transmission from parent to offspring, or one individual in a population to another, effectively passing on the resistance to certain pathogens.57–59
Various experiments have been done testing the effectiveness of adding antimicrobial peptides from resistant species, adding antifungal-producing bacteria, exposing to antifungal compounds (such as itraconazole), and treating with temperature changes. Many of these treatments were not effective, however valuable information was gained that can lead to better treatments. 60 In the experiments looking at the addition of antimicrobial peptides, the results indicated that it might help offer some resistance in species that do not produce them. The addition of antifungal compound-producing bacteria shows promise, in the event that the bacteria used can propagate on the new host without conferring disease itself. 60
Further work with the inhibitory isolates from B. bufo in GTNP is currently underway; and peptides believed to be responsible for the inhibition seen are currently being isolated and investigated, as part of a multiyear study.
Author Contributions
STP, SSH, and PPS conceived and designed the experiments. STP and PPS analyzed the data. STP wrote the first draft of the manuscript. AMC, SSH, and PPS contributed to the writing of the manuscript. STP, AMC, SSH, and PPS agree with manuscript results and conclusions. STP, SSH, and PPS jointly developed the structure and arguments for the paper. AMC and PPS made critical revisions and approved the final version. All authors reviewed and approved the final manuscript.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. this article was subject to blind, independent, expert peer review. the reviewers reported no competing interests.