
Abstract
Preparation of a single and pure phospholipid via transphosphatidylation has been a much sought after endeavor in the pharmaceutical and nonmedical industries. For this reason, phosphatidylglycerol, a lung surfactant, was produced from phosphatidylcholine with defined fatty acids, ie, dipalmitoyl phosphatidylcholine. Substrate type and concentration, enzyme source, and reaction temperature were investigated. Phospholipase D from two sources, ie, savoy cabbage, was purified in the authors’ laboratory and a commercially available Streptomyces species was used for this study. The substrates used were glycerol, a polyhydric alcohol, and solketal, a monohydric form of glycerol. The progress of the reaction was monitored using thin layer chromatography, and synthesis with solketal, an unusual form of glycerol, was confirmed by liquid chromatography mass spectrometry. Surface response methodology used on four combinations of enzyme and substrate at various temperatures (30 °C–60 °C) and concentration (0.25–1 mM) revealed that yield and selectivity was temperature-driven and predictable. To validate further the thermodynamic attributes, a modified version of the Eyring equation was derived from selectivity and the Arhenius equation. These equations provide some useful insights into the difference in activation of enthalpy change(ΔΔH++) and difference in activation of entropy change(ΔΔS++). Plots of ln[PG]/[PA] versus 1/T gave good linear fits for these four combinations. In addition, a new thermodynamic parameter known as TPG = PA has emerged as a theoretical temperature for equivalent transferase and hydrolase activity.
Introduction
Phospholipids as a group are a unique form of natural lipids in that their molecules possess a dual physical character. Their distinct physical behavior is brought about by a hydrophobic tail and hydrophilic head, to form a myriad of oil and water interactions. Phospholipids range from a simple micelle to a complex liposome, with various phases, including lamellar, hexagonal, and cubic. Phospholipids from natural sources, such as soy lecithin, have limited stability, emulsifying properties, and class-specific concentrations. Commonly found class-specific phospholipids include phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol, phosphatidylserine, and phosphatidylglycerol. Together with their corresponding lyso analogs, such as lyso-phosphatidylcholine, lyso-phosphatidylethanolamine, and lyso-phosphatidylinositol, 1 they form a complex phospholipid generally known as lecithin.
The limitations of phospholipids from natural sources are circumvented effectively by modification of soy lecithin and the enrichment of certain class-specific phospholipids. The latter usually involves physical means while the former involves chemical-assisted or enzymatic-assisted synthesis. The current trend of modification of phospholipids revolves around lipid biotechnology whereby lipase and phospholipase either from plant or microbes catalyze the change. Transesterification by the phospholipase A1 and A2 enzymes mediates the change on the acyl groups at the sn1 and sn2 positions, respectively. However, transphosphatidylation is a reaction that exchanges the polar head group with other primary hydroxyl-containing compounds, ie, primary alcohols 2 (Fig. 1). Phospholipase D (EC 3.1.4.4) catalyzes transphosphatidylation at the phosphodiester bond where the phosphatidyl moiety is transferred to an alcohol as a nucleophile donor to produce phosphatidyl alcohols.3–5 It is interesting to note here that in a non-catalyzed state, hydrolysis of a phosphodiester bond could take an inordinate period, ie, 180,000 years. 6
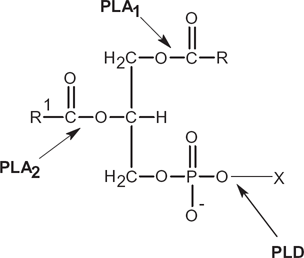
Various phospholipases (D, PLA1, and PLA2) in action on a phospholipid molecule.
Phospholipase D was originally extracted from plant-based sources like cabbage, peanuts, and cotton seed.7–9 Subsequent to its discovery, attempts were made to isolate phospholipase D from microorganisms.10,11 Of the various microbe strains identified, Actinomycetes, and specifically the genuses Streptomyces, Streptoverticilium, and Actinomudra, were commonly employed in transphosphatidylation.12,13
The mechanism of transphosphatidylation proceeds via a phosphohistidine intermediate, whereby a catalytic water molecule is believed to attack the phosphorus apically. The enzyme consists of 18α helices and 17β sheets in an α-β-α-β-α sandwiched configuration and possesses an active site made up of two identical sequence repeats of an HKD motif. 14 Hatanaka et al have investigated further the nature of the amino acid residues critical for thermostability of phospholipase D. 15
Enzyme-mediated transphosphatidylation is not only confined to naturally occurring head groups but also to unnatural polar groups. The factors that determine the candidacy for this change of hydroxyl compounds are type of hydroxyl group (primary or secondary), molecular size, substituent effects, and solubility. 2 The biocompatibility of the phosphatidyl moiety allows it to be used as a novel nontoxic carrier for food, cosmetics, and medical applications. For example, phosphatidylascorbic acid synthesized by Nagoa et al had improved scavenging effects on peroxy radicals. 16 Phosphatidylchromanol synthesized by Koga et al exhibited enhanced vitamin E activity. 17 Arbutin and kojic acid, which controls melanin in epidermal cells, was incorporated into a phosphatidyl group by Takami et al. 18 Shuto et al synthesized a phospholipid with anticancer agents such as 5-fluorouridine. This newly formed compound exhibited antitumor activity superior to that of its parent compound. 19 N-acetylneuraminic acid, a potent antiviral agent, was transphosphatidylated with phosphatidylcholine. A greater potency of 10 3 −10 4 was observed in mitigating rotavirus infection. 20
Phosphatidylglycerol plays an important role as one of the acidic polyglycerophospholipids.21,22 The earliest known study on the formation of phosphatidylglycerol by transphosphatidylation was reported in 1966 by Dawson, who concluded that a transferase reaction occurs simultaneously with hydrolase activity and that acceptor molecules need to possess a primary hydroxyl group. 23 Since then, further research has been carried out, including on the type of reaction medium (micelle and emulsion), 24 enzyme form (immobilized and free), enzyme stereoselectivity towards glycerol, 25 and transphosphatidylation reaction conditions. 26 Apart from its medical use as a lung surfactant, phosphatidylglycerol may be reacted further as a phosphatidylalkanol (head group with one more free primary hydroxyl group) with compounds having amino, carboxylic, halogen, and thiol functional groups. 27
Phospholipase D (EH) may act as a transferase (reaction 1), transferring A in the activated substrateAB from B to NH, or as a hydrolase in the hydrolysis of AB to AOH13,28 (reaction 2, see Fig 2). Analogous to the reactions above, transphosphatidylation of phosphatidylcholine with glycerol or solketal by phospholipase D produces phosphatidylglycerol24,29,30 (see Figs. 3 and 4). However, in the presence of water, hydrolysis competes with transferase activity via a competitive route to produce a phosphatidic acid (Fig. 5).
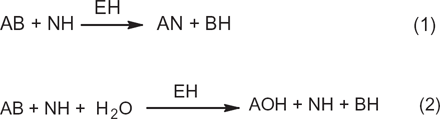
Transferase and hydrolase activity routes of phospholipase D.
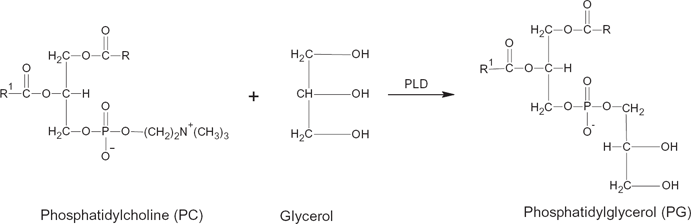
Transphosphatidylation of phosphatidylcholine with glycerol to form phosphatidylglycerol.
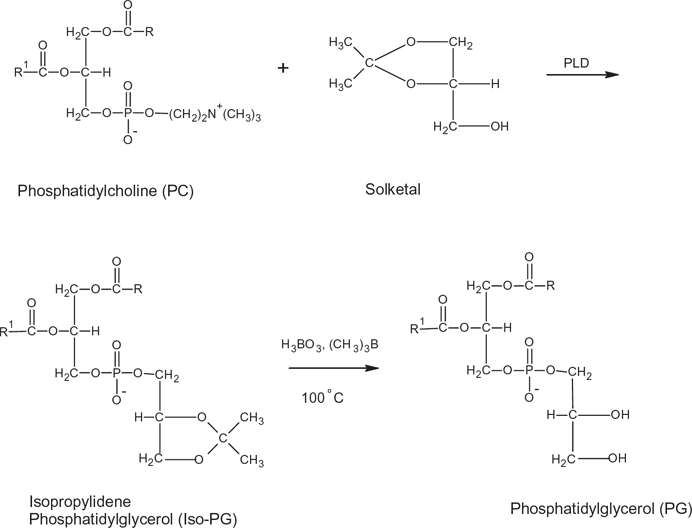
Transphosphatidylation of phosphatidylcholine with solketal to forming phosphatidylglycerol via iso-phosphatidylglycerol.
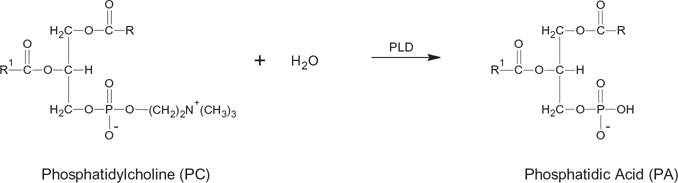
Hydrolysis of phosphatidylcholine forming phosphatidic acid.
The second order rates of AN and AOH formation are defined as follows:
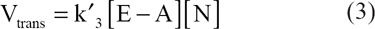
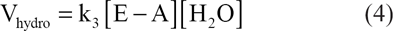
According to the Arrhenius equation, the fraction of molecules, f, having sufficient energy to surmount the activation energy barrier is represented by:

where R is the gas constant, T is the absolute temperature, and ΔEa is the activation energy.
Further treatment to incorporate the rate constant, k, renders the following with A as the collision frequency: 31

Abridged by statistical mechanics, Eyring substituted the activation energy term with the Gibbs free energy in Boltzmann distributed molecules at transition state.
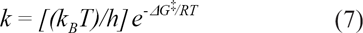
From ΔG‡ = ΔH‡-TΔS‡
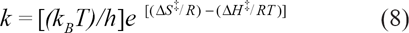
where k is the rate constant, k B is the Boltzmann constant, h is the plank constant, ΔG‡ is Gibbs free energy, and ΔH‡ is enthalpy of activation:32,33
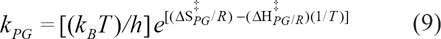
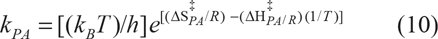
Gibbs free energy, ΔG‡ = ΔH‡-TΔS‡, being a state function, is independent of the paths that inter-convert compounds. Enthalpy controlled reactions occur when ΔH‡ is the major contributor to ΔG‡ and those that are entropy controlled occur when TΔS‡ is the major contributor to ΔG‡. Achieving the transition state is always endothermic because the bond strengths of the activated complex are lower than in the starting material. In general, the change in enthalpy on activation is significantly larger than the change in entropy, because much energy is required to break a bond in the transition state. Therefore, interpretation of absolute values of ΔS‡ has deficiencies. Negative values of ΔS‡ indicate decreasing values of translational and rotational degrees of freedom, or sometimes indicate the merger of two molecules, whereas positive values of ΔS‡ imply an increase in the translational and rotational degrees of freedom or often indicate the creation of two molecules. However, the difference between two competing reaction substrates, ie, ΔΔS‡ (difference in activation of entropy change) made for the reaction of similar molecularity is in order here. 34 An example of this extension was employed in the study of the diastereospecificity ratio, E.35,36
By combining equations (9) and (10) the following is obtained:
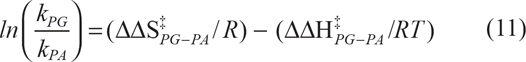
where ΔΔS‡ PG-PA = ΔS‡ PG –ΔS‡ PA and ΔΔH‡ PG-PA = ΔH‡ PG –ΔH‡ PA .
By analogy with the diastereospecificity ratio, E, a new selectivity ratio for transferase to hydrolase may be defined as k PG /k PA where k PG /k PA ≈ [PG]/[PA]. The modified Eyring equation is finally deduced, ie, equation (12) from linear plots of ln([PG]/[PA]) and 1/T
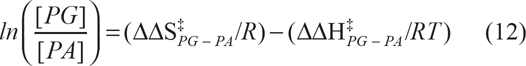
The values of PG/PA can be extracted from the selectivity relationship of transphosphatidylation
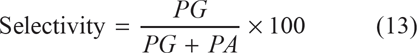
and
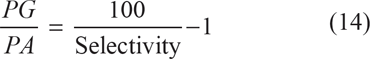
When k3 = k3 the concentration, [PA] = [PG], and its reaction selectivity equals 50%. It follows that ln[PA]/[PG] = 0. Then from equation (12)
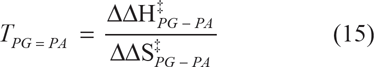
The temperature TPG = PA subsumes equal activities of hydrolase and transferase.
Materials and Methods
Dipalmitoyl phosphatidylcholine (DPPC) was prepared from industrial grade soy lecithin (60% acetone insoluble) via silica column chromatography and later deacylated with tetrabutylammonium hydroxide to form glycerophosphatidylcholine. Further reacylation of glycerophosphatidylcholine with palmitic anhydride and 4-Dimethylaminopyridine (DMAP) as the catalyst was carried out according to the methods described by Regen et al to form dipalmitoyl phosphatidylcholine.37,38
A crude extract of phospholipase D was obtained from fresh savoy cabbage and purified by hydrophobic interaction chromatography using the methods described by Lambrecht et al. 39 The activity of phospholipase D cabbage was determined using choline oxidase and peroxsidase as described by Imamura et al 40 with DPPC produced earlier as a substrate. The amount of protein measured was determined using the Bradford test. Other chemicals, including glycerol, solketal, diethyl ether, calcium chloride, and acetic acid, were of analytical grade from Acros Organic, USA. All other reagents used were of analytical grade unless otherwise stated.
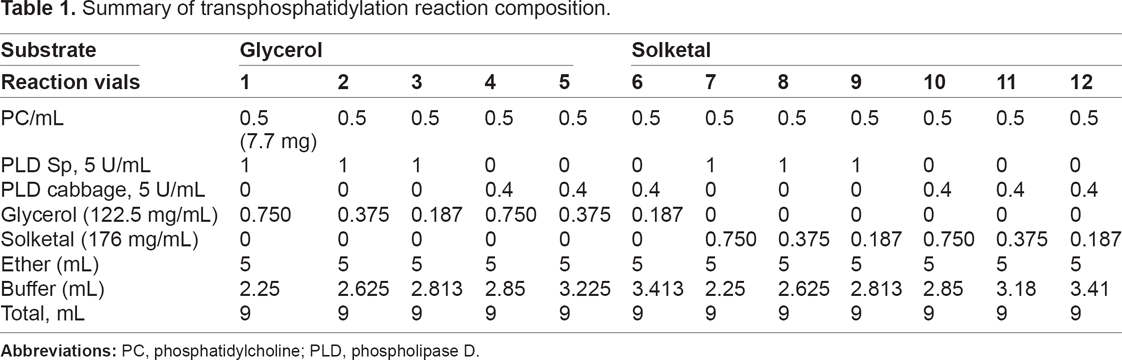
Transphosphatidylation was carried out in a 25 mL Erlenmeyer flask with a Teflon stopper. Briefly, 0.5 mL of DPPC (7.7 mg) was pipetted into the flask, and 5 units of enzyme activity stabilized in 0.2 M acetic acid-sodium acetate buffer at pH 5.6 was added together with 1.3 M substrates of either glycerol or solketal in 80 mM CaCl2 (substrate excess was 100 to 25 times more than DPPC).26,27 The reaction was carried out at four different temperatures, namely 30 °C, 40 °C, 50 °C, and 60 °C. Finally, 5 mL ether was added to initiate the reaction. The flask was kept in a water bath with its working shaker set at 150 rpm. After 12 hours, the reaction was terminated by adding 3 mL of chloroform-methanol (2:1) to the reaction vessel. The upper phase was carefully collected and dried with a stream of nitrogen at atmospheric temperature (30 °C). The dried product was reconstituted with chloroform-methanol (2:1, vol/vol) to a concentration of 1 mg/mL for further quantification by high-pressure liquid chromatography (HPLC) as described below. For the reaction with solketal, an additional step was carried out to remove the ketal protecting group where 1.5 mL trimethyl borate together with 80 mg boric acid was heated up to 100 °C for 3 hours 30 (Fig. 4). The reaction compositions are summarized in Table 1.
Summary of transphosphatidylation reaction composition.
Phospholipid class quantification of phosphatidylcholine, phosphatidylglycerol, and phosphatidic acid was separated by HPLC (Agilent 1100 series) equipped with an ELSD detector (Altech 2000). The column used was a PVA-Sil (5 μm, 250 × 4 mm) from YMC Japan and a corresponding guard column Guardpack PVA-SIL-NP (10 × 4 mm ID 5 μm, 12 nm). The ternary eluent system used was as described by Christie et al 41 and Beermann et al. 42 Briefly, eluent A was n-hexane/ter-methylbutyl ether (98:2), eluent B was isopropanol/acetonitrile/chloroform/acetic acid (84:8:8:8:0.0025), and eluent C was isopropanol/water/triethylamine (49.9:49.9:0.2). All solvents used were of HPLC grade from Fischer Scientific Pty Ltd. The solvent program was as follows: 0–1 minute A/B/C (%) 100:0:0; 1–5 minutes A/B/C (%) 80:20:0; 5–20 minutes A/B/C (%) 42:52:6; 20–27 minutes A/B/C (%) 32:52:16; 27–30 minutes A/B/C (%) 30:54:16; 30–33 Minutes A/B/C (%) 30:70:0; and 33–40 minutes A/B/C (%) 100:0:0. The flow rate was 1 mL/minute and the injection volume was set at 20 μL. A standard calibration curve of phospholipid classes between 0.1 mg/mL and 2 mg/mL was used for the quantification. All standard soy phospholipids were purchased from Sigma-Aldrich.
Mass spectrometry determination via liquid chromatography was carried out to confirm the formation of isopropylidene phosphatidylglycerol (Fig. 4). A Waters Alliance HT 2795 coupled to a Bruker MicroTOF-Q MS was employed with the previous gradient and column setting. Ionization was accomplished using an electrospray ionization in the positive ion mode. Mass range scan was from 50 to 1000 m/z. The nebulizer was set at 2 bar with its dry heater at 190 °C and dry gas (N2) at 7.0 L/minute.
Response surface methodology was carried out for responses, namely percent selectivity of phosphatidylglycerol against phosphatidic acid and percent yield of phosphatidylglycerol. A total of 15 experiments were recommended according to the central composite design, where five were center points and the remaining eight were noncenter points. An addition of two noncentered points was introduced as recommended by the software for model sufficiency. The variable factors were temperature (30 °C–60 °C) and substrate concentration (0.25–1.0 mM). The experimental design was generated by Design-Expert version 6.0.10 (Stat Ease software).
Results and Discussion
Enzyme activity of cabbage phospholipase D
One enzyme unit is defined as 1 μmol of choline liberated in one hour from L-α-phosphatidylcholine at pH 8 and temperature 30 °C. Choline 0.6234 μmol was liberated when 0.05 mL of activity-loaded hydrophobic interaction chromatography solution was assayed. In short, the cabbage phospholipase D activity was 12.5 units/mL and protein content was 5 mg/mL.
Dipalmitoyl phosphatidylcholine
Synthesis was carried out until most of the lyso analog disappeared from the thin layer chromatography plates. After purification and workup, the yield obtained was 97% (Fig. 6).

Thin layer chromatography of dipalmitoyl phosphatidylcholine with traces of its lyso analog.
Dipalmitoyl phosphatidylsolketal
With regard to structural confirmation of synthesized dipalmitoyl phosphatidylsolketal (1,2-dipalmitoyl-sn-glycero-3-fosfo-1′,2′-O-isopropylidien-sn-glycerol), from the mass chromatogram in Figure 7, there are two prominent peaks at m/z 764.57 and m/z 239.22. The peak m/z 764.57 represents the compound [M+H]+ for 1,2 dipalmitoyl-sn-glycerol-3-fosfo-1′,2′-O-isopropilidiene-sn-glycerol, whereas the latter at m/z 239.22 describes the ketene form of palmitic acid [C14H29–CH = C = O + H]+. The ketene form is introduced via a rearrangement of the carboxylic acid R1COOH, forming the said compound and water, R1CH = C = O + H2O where R1 = C14H29.
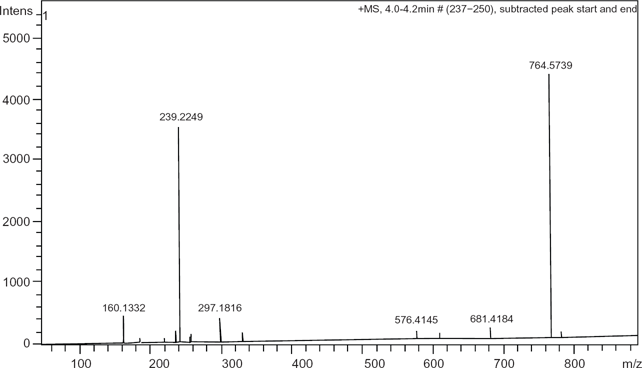
Positive electrospray ionization, electrospray ionization mass spectra of 1,2-dipalmitoyl-sn-glycero-3-fosfo-1′,2′-O-isopropylidien-sn-glycerol generated from a liquid chromatography mass spectrometry run.
Response surface methodology
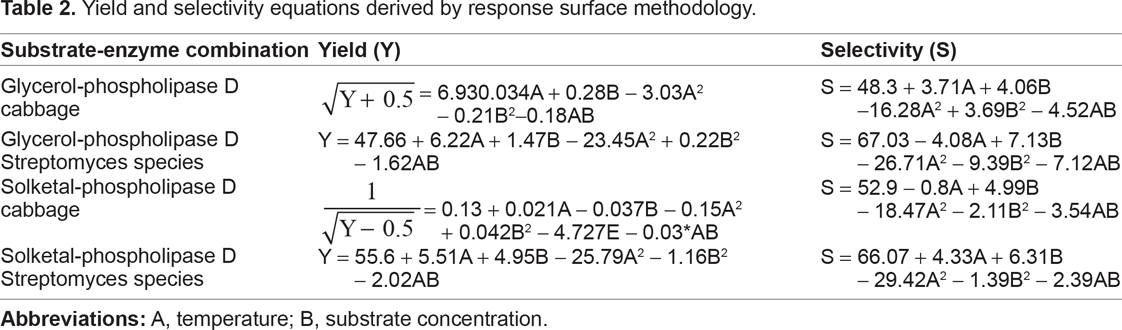
The response surface methodology treatment of glycerol-Phospholipase D cabbage yield obeys the square root of
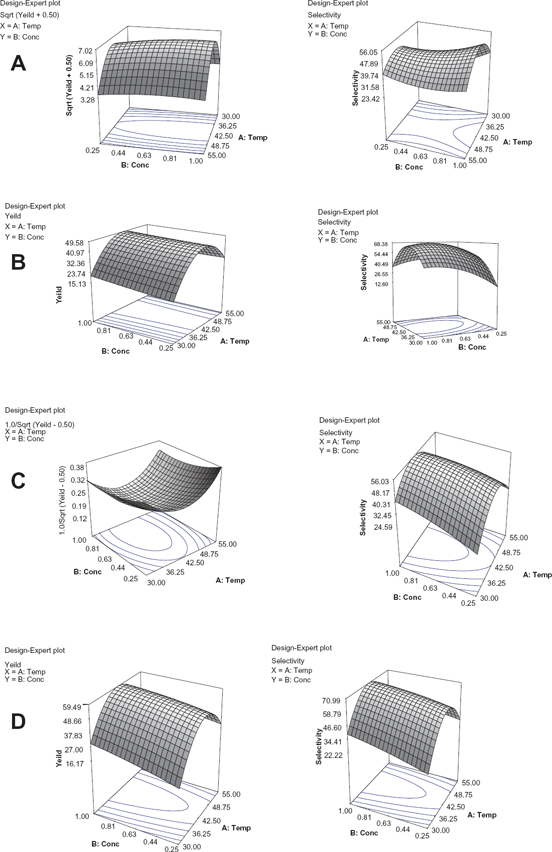
Response surface plot on yield and selectivity with temperature and substrate concentration as variables. (A) glycerol-phospholipase D cabbage, (B) glycerol-phospholipase D Streptomyces species, (C) solketal-phospholipase D cabbage, and (D) solketal-phospholipase D streptomyces species.
Yield and selectivity equations derived by response surface methodology.
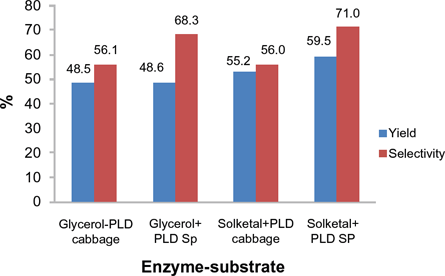
Surface response methodology optimization for maximum yield and selectivity (percentage reading rounded up to one decimal point).
Selectivity increases to a maximum of 68.3% and yield to 48.6% for the glycerol-phospholipase D Streptomyces species combination. This occurs when the temperature and glycerol concentration is 43.3 °C and 0.76 mM, respectively. The loss of linearity seen when the glycerol concentration increases suggests a higher sensitivity of phospholipase D Streptomyces species toward glycerol. This loss of linearity (R2 = 0.6861) could not explain the non-significant lack of fit of the model for selectivity (P > 0.1). One possible reason for this sensitivity is the additional reaction of the second hydroxyl group on glycerol to form cardiolipin. 9
The yield in the reaction of solketal-phospholipase D cabbage is related via a square root transformation
The optimum temperature for highest yield/selectivity when the phospholipase D Streptomyces species was used was the same when either solketal or glycerol was introduced as a substrate, ie, 43.3 °C. However, a relatively higher yield and selectivity was observed for solketal-phospholipase D Streptomyces species, where yield reached 59.5% and selectivity 71.0% compared with yield of 48.6% and selectivity of 68.3% for glycerol-phospholipase D Streptomyces species. For substrate concentration, the amount of glycerol required for optimum temperature was 0.80 mM as compared with 1 mM of solketal.
Thermodynamic treatment
Table 3 gives ΔΔH‡ PA-PG , ΔΔS‡ PA-PG , and T PA=PG for the formation of phosphatidylglycerol and phosphatidic acid as determined from the ln [PA]/[PG] versus 1/T plots (Fig. 10).
Thermodynamic parameters generated via modified Eyring equations.
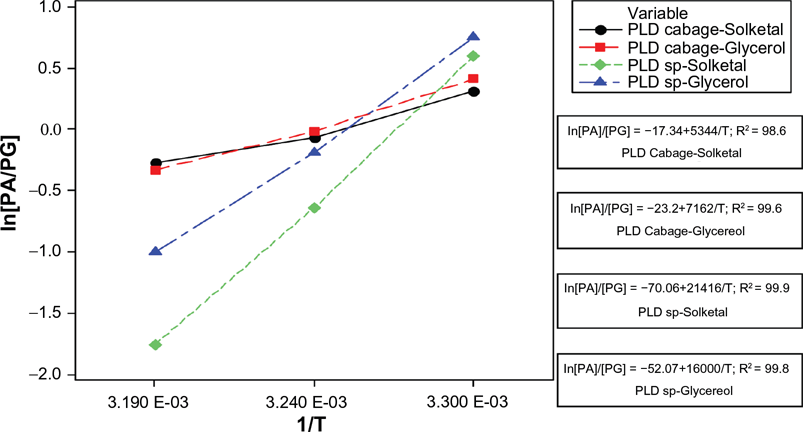
Modified Eyring plot derived from substrate concentration of 1 mM and selectivity at 30 °C, 35 °C, and 40 °C. (results for 35 °C was predicted from surface response methodology).
ΔΔH‡ PG-PA generally is greater for phospholipase D Streptomyces species. This indicates a propensity for transferase during transphosphatidylation. The better selectivity for solketal is delineated in the ΔΔH‡ PG-PA values reported in Table 3, ie. -42.48 kcal/ mol compared with -31.74 kcal/mol for glycerol. Further validation may be gleaned from the selectivity provided by response surface methodology where the selectivity of solketal reached 71% and 68.3% for glycerol (Fig 9).
Another significant parallel with the increased selectivity of solketal is its lower T PG = PA . It is also observed from Table 3 that T PA = PG = 34.96 °C via response surface methodology is similar regardless of the substrate used for phospholipase D Streptomyces species. The less positive values in the entropy difference ΔΔS‡PG-PA for solketal with phospholipase D species suggest a dominance of transferase activity, hence a preferred substrate against glycerol.
ΔΔH‡ PG-PA for cabbage phospholipase D and glycerol was -14.20 kcal/mol. The lower selectivity for glycerol demonstrates subjugation of phospholipase D cabbage. The entropy difference ΔΔS‡ PG-PA = -46.02 cal/mol k is lower (less positive), indicating lesser anchoring of glycerol during the transition state. The value of T PA = PG is lowest for phospholipase D species, derived either by Eyring or response surface methodology.
A greater positive value of ΔΔS‡ PG-PA for cabbage phospholipase D reveals an increase of hydrolase activity as evidenced by the rise in phosphatidic acid concentration. Hence the value of ΔΔS‡ PG-PA = −34.38 cal mol−1 k for phospholipase D cabbage-solketal should show the greatest loss as compared with the other combinations of enzyme-substrate. This intuitively could be caused by the bulky ketal group where two methyl groups form part of the protective group of glycerol (Fig. 11).

Structure of solketal and glycerol in three-dimensional representation (red, oxygen; green, carbon; white, hydrogen).
The ketal group essentially increases the hydrophobic part of the molecule while reducing the H-bond potential and electrostatic interactions, hence the relative positive value of ΔΔS‡ PG-PA . However, the optimum selectivity of phospholipase D cabbage-glycerol and phospholipase D cabbage-solketal is invariant, ie, 56%. The temperature of T PA = PG is relatively higher for those reactions catalyzed by phospholipase D Streptomyces species. This suggests a higher activation temperature for phospholipase D cabbage. Unlike phospholipase D cabbage, the phospholipase D from Streptomyces exhibits broader substrate contact and binding, even for a relatively hydrophobic substrate surface such as solketal.
Conclusion
Eyring-derived thermodynamic parameters constitute a suitable method for deciphering the entropy and enthalpy relationship in transphosphatidylation. It was observed that the larger negative value in the difference of enthalpy of activation, ΔΔH‡ PG-PA for phospholipase D from Streptomyces species meant higher selectivity toward alcohol acceptors (substrate). The smaller negative values for phospholipase D cabbage demonstrate its propensity to form phosphatidic acid by hydrolysis. A larger negative value for the difference in entropy of activation change, ΔΔS‡ PG-PA , indicates that phosphatidylglycerol is tightly bound in the transition state.
The temperature when selectivity is 50%, T PA = PG is a helpful marker to predict the beginning points when phosphatidylglycerol is produced in access when T > T PA = PG transferase activity dominates until maximum selectivity is reached (Fig. 9). The temperature, T PA = PG is another means by which to predict relative activation temperature. The enzyme phospholipase D from Streptomyces species is not affected by type of substrate but is signatured by a lower T PA = PG , indicating a lower activation temperature. However, phospholipase D cabbage possesses a higher T PA = PG .
Footnotes
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.