
Abstract
It is well known that mammalian glycosphingolipids (GSL) play key roles in different physiological and pathophysiological processes. The simplest GSL, glucosylceramide (GlcCer), is formed through the enzymatic transfer of glucose to a ceramide moiety. In mammalian cells this molecule is the building block for the synthesis of lactosylceramides and many other complex GSLs. In fungal cells GlcCer is a major neutral GSL that has been considered during decades merely as a structural component of cell membranes. The recent literature, however, describes the participation of fungal GlcCer in vital processes such as secretion, cell wall assembly, recognition by the immune system and regulation of virulence. In this review we discuss the most recent information regarding fungal GlcCer, including (i) new aspects of GlcCer metabolism, (ii) the involvement of these molecules in virulence mechanisms, (iii) their role as targets of new antifungal drugs and immunotherapeutic agents and, finally, (v) their potential participation on cellular signaling in response to different stimuli.
Introduction
Glycosphingolipids (GSL) comprise a diverse class of molecules involved in membrane stability in different cell types, including fungal pathogens. In the last decades, these molecules have been recognized as relevant structures in processes of cell-cell interaction, cell signaling and lipid raft formation in mammal cells. In fungal organisms a conserved structure was described as the major neutral GSL, the N-2'-hydroxy(hexa/octa)de(ca/ce)noyl-1-beta-D-glucopyranosyl-9-methyl-4,8-sphingadienine or glucosylceramide (GlcCer) (Fig. 1). Although these GSL were first characterized in fungi more than two decades ago, knowledge about their functions remained obscure until very recently. In the present review we discuss the most recent findings that characterize GlcCer as essential structures in fungal cells.
Major structures of fungal GlcCer. Different fatty acids (R1, R2, R3 or R4) can associate to the sphingoid base (9-methyl-4,8-sphingadienine) forming ceramide.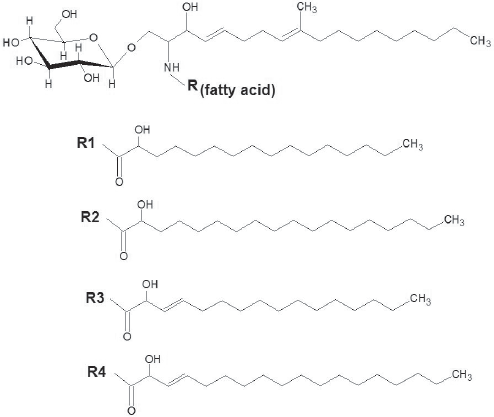
Biosynthesis, Sorting Out and Turnover of GlcCer in Fungal Cells
The elucidation of the pathways required for GSL biosynthesis in eukaryotic organisms originated from studies using mammalian cells (Barreto-Bergter et al. 2004; Guan and Wenk, 2008) and the yeast Saccharomyces cerevisiae (Cowart and Obeid, 2007). Although these studies provided a solid base for the comprehension of how GSL are synthesized, the mammalian and the S. cerevisiae models are not, however, adequate prototypes for the understanding of the synthesis of GlcCer in fungal pathogens. Mammalian GlcCer, for instance, are structurally different from their fungal counterparts, which denote the existence of distinct enzymatic machineries for the synthesis of these glycolipids (Barreto-Bergter et al. 2004). On the other hand, although S. cerevisiae is a fungal species that shares many common characteristics with pathogens, it lacks the gene(s) responsible for the synthesis of GlcCer, as also described in other fungal species (Saito et al. 2006). Importantly, it is notorious that all fungal pathogens studied so far are able to synthesize GlcCer (Nimrichter et al. 2005b) with the single exception of Candida glabrata (Saito et al. 2006). Interestingly, C. glabrata is phylogenetically closer to S. cerevisiae than to other species of the Candida genus (Barns et al. 1991; Hendriks et al. 1991).
The initial steps of GlcCer synthesis in animals and fungi have common features, although with a variety of products being formed. Ceramide synthesis occurs in the endoplasmic reticulum (ER) involving a series of reactions (for more details see (Barreto-Bergter et al. 2004)). Important differences were shown in the ceramide metabolism in animal and fungal cells. The sphingoid bases of the ceramide moiety of GlcCer in fungi have a methyl group at C9 and an extra unsaturation at C8, forming the typical fungal base 9-methyl-4,8-sphingadienine (Barreto-Bergter et al. 2004; Nimrichter et al. 2005b). By using bioinformatics, Ternes et al. (Ternes et al. 2006) identified the gene responsible for the transference of the C9-methyl group to the sphingoid base of fungal GlcCer. The enzyme, which belongs to the superfamily of S-adenosylmethionine-(SAM)-dependent methyltransferases, appears to be membrane-bound and requires a δ4,8-desaturated ceramide as substrate. Since the enzyme is absent in animal cells it could be a potential target for the development of new antifungal drugs. Another potentially relevant difference between mammalian and fungal ceramides is the fact that fungal organisms preferentially use C16 or C18 hydroxylated fatty acids to synthesize GlcCer (Barreto-Bergter et al. 2004).
Through vesicular and non-vesicular routes (Funato and Riezman, 2001; Halter et al. 2007; Vallee and Riezman, 2005), ceramide is transported to the Golgi, where the addition of the polar-head moiety of GlcCer occurs. The transfer of glucose to a ceramide moiety is performed by a ceramide glucosyltransferase (Cer:GlcT), also called ceramide glucosyl synthetase (CGS), an enzyme expressed at Golgi stacks (Halter et al. 2007).
Differently from GlcCer, ceramides that are used for the synthesis of galactocerebrosides (galactosylceramide, GalCer) are glycosylated at the ER (Sprong et al. 2003; Sprong et al. 1998). As discussed previously, with the exception of C. glabrata, all fungal pathogens appear to synthesize GlcCer. It is possible that fungal cells may undertake additional glycosylation of monohexosylceramides, since a dihexosyl ceramide (lactosylceramide) has been reported in lipid extracts of the fungal phytopathogen Magnaporthe grisea (Maciel et al. 2002). The biosynthetic pathways involved in such process are still unclear, since phytosphingosine and a long-chain fatty acid (OH-C24) form the hydrophobic moiety of M. grisea ceramide dihexoside. In comparison, complex phytoceramide- or dihydroceramide-containing GSL are synthesized in yeast (Cowart and Obeid, 2006). The main classes of GSL produced in S. cerevisiae are: inositolphosphoceramides (IPC), mannose-inositolphosphoceramides (MIPC) and mannose-(inositol-P)2-ceramides (M(IP)2C). These molecules are also expressed in other fungal species but they are not the focus of this review (Barr and Lester, 1984; Bennion et al. 2003; Heise et al. 2002; Levery et al. 1998).
Finely regulated mechanisms are required for the correct sorting of biomolecules to their major cell sites, including different organelles and / or the cell surface (Degroote et al. 2004; Watanabe and Riezman, 2004). In the case of fungal GlcCer, these sites include the external layer of the plasma membrane and also the cell wall (Barreto-Bergter et al. 2004; Nimrichter et al. 2004; Rodrigues et al. 2000). The detailed mechanisms of cellular trafficking of fungal GlcCer are still unknown, but recent studies in mammalian cells suggest that cerebrosides that are not utilized for the synthesis of complex GSL reach the cell surface via a non-vesicular mechanism (Halter et al. 2007). The existence of two different pools of ceramides to be used in the synthesis of fungal monohexosylceramides and complex GSL (Leipelt et al. 2001) suggests that sorting and addition of polar head groups could be controlled by the lipid portion, but additional experiments must be performed to confirm this possibility. It has been demonstrated, however, that fungal cells overexpressing glucosylceramide synthases formed glucosylceramides that contained either C18 or very long chain fatty acids (Leipelt et al. 2001), suggesting that the regulation of GSL synthesis depends on cellular compartmentalization of substrates rather than enzyme specificity. It is also well-known that the ceramide moiety is essential to direct the entire GSL molecule towards cell membranes (Tagawa et al. 2002), as well as the fact that it participates in sorting mechanisms (Degroote et al. 2004).
The involvement of GlcCer in sorting and secretion mechanisms was recently suggested for the first time (Rodrigues et al. 2007a). GlcCer and sterols were demonstrated to be lipid components of secretory vesicles in the yeast pathogen Cryptococcus neoformans (see more details in ref. (Rodrigues et al. 2007a). In association, GSL and sterols are components of lipid domains of cell membranes, the so-called lipid rafts. These domains also aggregate biologically significant proteins, such as GPI anchored molecules (Kasahara and Sanai, 2000). Lipid rafts are present in different cell types, including non-pathogenic and pathogenic fungi (Insenser et al. 2006; Martin and Konopka, 2004; Siafakas et al. 2006; Wachtler and Balasubramanian, 2006). Lipid rafts are directly linked to many cellular functions but in fungal cells, since they participate in polarization and compartmentalization steps of the plasma membrane, they seem to be of special relevance (Martin and Konopka, 2004; Siafakas et al. 2006; Wachtler and Balasubramanian, 2006). In addition, a compositional analysis of lipid rafts in membranes of C. neoformans revealed that they concentrate virulence determinants, including phospholipase B1 (PLB1), GPI-anchored proteins, and Cu/Zn superoxide dismutase (SOD1) (Siafakas et al. 2006). Lipid analysis confirmed the presence of sterols and GlcCer, the later representing the major polar component in raft preparations. In C. albicans, another opportunistic fungus that produces GlcCer, staining of yeast cells with filipin revealed that lipid rafts were concentrated at the growing tip of hyphal cells in all stages of hyphal growth and at sites of septum formation in mature hyphae, suggesting a correlation with fungal growth. This profile of filipin-staining in C. albicans was conditioned to a regular synthesis of GSL and sterol, which also supports the supposition that lipid rafts and fungal growth are associated (Martin and Konopka, 2004). Proteomic analysis demonstrated that lipid rafts in C. albicans aggregate proteins involved in glycosylation, lipid metabolism as well as heat shock proteins (Insenser et al. 2006). Interestingly, GlcCer and sterol enriched-secretory vesicles in fungal pathogens, contain different types of protein, including those described as lipid raft components of C. albicans (Rodrigues et al. 2008). The role of GlcCer and lipid rafts during vesicle formation is under investigation in our laboratory.
Since GSL are required for lipid raft formation, inhibitors of Cer:GlcT have been used to study the participation of such domains in various physiological events (Levery et al. 2002; Sottocornola et al. 2006; Yanagisawa et al. 2005). Although these inhibitors are very helpful in studies involving animal cells, their inhibitory effect on the fungal enzyme appears to be controversial. In Aspergillus species, exposure to inhibitors of ceramide glycosylation, D-threo-1-phenyl-2-palmitoyl-3-pyrrolidinopropanol (P4) and D-threo-3P,4P-ethylenedioxy-P4 (EDO-P4), resulted in a significant reduction of GlcCer formation and strongly affected spore germination, cell cycle, and hyphal growth (Levery et al. 2002). In addition, incubation with 1-phenyl-2-decanoylamino-3-morpholino-1-propanol (PDMP) significantly reduced capsule formation in C. neoformans (unpublished data). Interestingly, capsular polysaccharides are also components of vesicles secreted by C. neoformans indicating a potential correlation between GlcCer and polysaccharide secretion (Rodrigues et al. 2007a).
To study the activity of GSL inhibitors in different cell models Hillig et al. (Hillig et al. 2005) overexpressed Cer:GlcT cDNAs from different organisms in the yeast Pichia pastoris, including the Cer:GlcT from the fungi Ustilago maydis, C. albicans, A. nidulans and P. pastoris. Cell-free membranes were prepared and the activity of Cer: GlcT was measured in the presence of EDO-P4. The results showed that the activity of the human enzyme, but not that of fungal enzymes was strongly inhibited by EDO-P4. Although this appears to limit the use of Cer:GlcT inhibitors in studies on the metabolism of fungal GlcCer, it also suggests that different inhibitors could be produced to abrogate the fungal Cer:GlcT activity in a very specific manner. Moreover, it is possible that other fungal species express Cer: GlcT that could be inhibited by PDMP or its derivatives.
It is well known that mammalian GlcCer are constantly internalized and hydrolyzed in endosomal and lysosomal compartments. A perfect equilibrium of synthesis and degradation of GSL is required in higher organisms in which failure in the turnover of lipids has severe consequences (Jeyakumar et al. 2002). The degradation products include long chain bases and fatty acids, or even ceramides, which can be reutilized to form new GSL. Since fungal species are also eukaryotic organisms, GSL turnover is also a likely event. Indeed, some enzymes and organelles required for endocytosis and molecular turnover of fungal lipids have been described through genetic and biochemical studies (Fuchs and Steinberg, 2005; Read and Kalkman, 2003). The transport of GlcCer, and probably of other GSL as well, to the cell wall raises the question of the surface turnover of these molecules. Studies from our laboratory revealed indeed that at least 75% of the total GlcCer content in fungal cells is associated with the cell wall (unpublished data) possibly being expressed in vesicles which may be endowed with a turnover mechanism. It has been suggested that GlcCer is a component of fungal exosomes, which indicates that this molecule participates actively in exocytic events (Rodrigues et al. 2008; Rodrigues et al. 2007a).
Glucosylceramides are Lipid Regulators of Fungal Virulence
As discussed previously, the relevance of monohexosylceramides as structural components of cellular membranes has been well known for several years. New aspects about functions and cellular distribution of GlcCer in fungi, as well as their roles as targets of antimicrobial antibodies and peptides, were more recently described (Barreto-Bergter et al. 2004; Kechichian et al. 2007; Rittershaus et al. 2006; Rodrigues et al. 2007a; Rodrigues et al. 2007b; Rodrigues et al. 2000; Thevissen et al. 2004) and are discussed in this review. Recent approaches to study GlcCer functions in fungi, however, revealed a new and unexpected role of this glycolipid in animal mycoses (Mitchell, 2006; Rittershaus et al. 2006).
Infections caused by yeast forms of the pathogen C. neoformans are acquired by inhalation of desiccated cells, which are available in the environment as basidiospores or poorly encapsulated yeasts (Perfect and Casadevall, 2002). Inhaled cells are first deposited into the alveolar space to then reach the lung interstitium. At the alveolar space, C. neoformans probably interacts with alveolar macrophages and epithelial cells (Barbosa et al. 2007; Barbosa et al. 2006). The infection is normally controlled by cell mediated immunity in immunocompetent patients, but in immunosuppressed patients cryptococci can disseminate to the central nervous system causing a high level of mortality (Perfect and Casadevall, 2002). During dissemination, C. neoformans interacts with different tissues and host microenvironments. Some of these, including the bloodstream and alveolar spaces are characterized by neutral pH. Other niches in which C. neoformans is frequently detected, such as the intracellular environment of macrophages (Alvarez and Casadevall, 2006), have acidic pH values.
Interestingly, Saito et al. (Saito et al. 2006) demonstrated that GlcCer expression is associated with fungal growth in alkaline conditions. The presence of GlcCer and its relationship with the pH appears to be relevant also during fungal infections. In C. neoformans, a mutant was obtained by Rittershaus et al. (Rittershaus et al. 2006) by deletion of GCS1. This is the gene encoding cryptococcal GCS, the enzyme involved in the final step of GlcCer biosynthesis. As expected, the C. neoformans δgcs1 mutant was unable to produce GlcCer, in contrast to the wild type and a reconstituted strain. Interestingly, when the δgcs1 mutant was exposed to high (5%) CO2 levels at neutral pH, fungal growth was inhibited. At an acidic pH, the C. neoformans δgcs1 mutant strain grew normally. These findings established an unexpected and still poorly understood connection between GlcCer synthesis and the fungal response to physiological and CO2 pH. In addition, the susceptibility of the δgcs1 mutant to high CO2 levels at neutral pH raised new concepts about the pathogenesis of C. neoformans (Mitchell, 2006), as detailed below.
Deletion of the GCS1 gene did not affect pathogenesis-related characteristics of C. neoformans, such as capsule production, melanin formation, growth at physiological CO2 and 37 °C (Rittershaus et al. 2006). In an animal model of cryptococcosis, however, the δgcs1 mutant was completely avirulent following nasal inhalation. In contrast, the mutant strain caused lethal infection when inoculated by intravenous injection. The average survival of mice infected intranasally with WT and δgcs1 + GCS1 strains extended to 24–27 days, while all mice infected with the δgcs1 mutant remained alive after 90 days of infection. All animals infected intravenously died after 6 days of infection with strains WT and δgcs1 + GCS1 and 15 days after infection with the δgcs1 mutant (Rittershaus et al. 2006). Histopathological analysis of the brains of mice infected intranasally with δgsc1 showed no sign of yeast cells (Rittershaus et al. 2006). In fact, δgcs1 cells were mostly entrapped at the center of an intense granulomatous response in the lungs of mice challenged intranasally. In contrast, when C. neoformans δgsc1 cells were inoculated intravenously, dissemination to the brain, kidney and liver was observed, which was associated with animal death.
In the context described above, the fact that the C. neoformans mutant lacking GlcCer is avirulent and unable to reach the brain when administered intranasally suggests that lung granuloma formation is a key event in the control of the infection in mice by δgcs1 cells. Using a T-and NK-cell-immunodeficient mice in which alveolar macrophages are not activated and, therefore, granuloma formation is not expected to occur, Kechichian et al. (Kechichian et al. 2007) demonstrated that the δgcs1 mutant proliferates in the lungs and does disseminate to the brain, although its virulence phenotype is dramatically reduced. Given that the δgcs1 mutant grows regularly within acidic lysosomes of macrophages, it has been hypothesized that mutant yeast cells lacking GlcCer proliferate inside alveolar macrophages of immunodeficient mice. In fact, depletion of alveolar macrophages through administration of clodronatecarrying liposomes improved survival of T- and NK-cell-immunodeficient mice and decreased the dissemination of δgcs1 cells to the central nervous system (Kechichian et al. 2007). These results suggest that the growth of δgcs1 in immunodeficient mice is maintained within alveolar macrophages. The δgcs1 cells may have their growth inhibited in the extracellular space of the lung, where CO2 levels are high (5%) and the pH is neutral. After ingestion by alveolar macrophages, C. neoformans mutants would grow in the acidic intracellular pH, which would be decisive for the successful pathogenic process.
The results described above demonstrate that GlcCer is essential for C. neoformans growth in the blood and alveolar spaces of the lung, which in contrast to the acidic intracellular environment in macrophages, have a neutral pH. These studies thus characterized for the first time a microbial lipid as a virulence regulator. In C. neoformans, GlcCer apparently does not control virulence through any of the previously mentioned phenotypic traits, but by a connection between lipid-mediated signaling and the pathogen's response to the CO2 levels and pH of host tissue. In fact, it has been shown that adenylyl cyclase can play a key cellular role acting as a conserved CO2 sensor (Mitchell, 2005). In C. neoformans, a membrane associated adenylyl cyclase can be regulated by CO2 (Mogensen et al. 2006). These observations raise the possibility that GlcCer may modulate adenylyl cyclase activity (Mitchell, 2006), although this has not been experimentally proved.
The contribution of GlcCer to fungal virulence is apparently not restricted to the regulation of microbial growth in different habitats during animal infection. The idea that GlcCer could be involved in different cellular events was first proposed after its cellular distribution was depicted in C. neoformans (Nimrichter et al. 2005b; Rodrigues et al. 2000). These studies demonstrated that the accumulation site of GlcCer was not limited to the plasma membrane, and mainly included the fungal cell wall. A detailed transmission electron microscopy revealed that the glycolipid was a component of vesicle-like structures found at the periplasmic space of C. neoformans (Rodrigues et al. 2000), some of them migrating from the plasma membrane to the cell wall. These vesicles contained a bilayered membrane and were recognized by a monoclonal antibody to GlcCer. Further studies demonstrated that GlcCer-containing compartments were also present at the inner and external layers of the cell wall of cryptococci (Nimrichter et al. 2005b; Rodrigues et al. 2000), as well as in external vesicle like-structures (Rodrigues et al. 2007a). These observations suggested that GlcCer could be a component of fungal secretory vesicles, which has recently been confirmed by Rodrigues et al. (Rodrigues et al. 2007a). Structural analysis of extracellular vesicles produced by C. neoformans revealed the presence of N-2'-hydroxyhexadecanoyl- and N-2'-hydroxyoctadecanoyl-1-βD-glucopyranosyl-9-methyl-4,8-sphingadienine. Interestingly, several virulence determinants were also identified in the extracellular vesicles from C. neoformans (Rodrigues et al. 2008), confirming that GlcCer synthesis is a key event in the secretion of pathogenesis-promoting molecules.
The Therapeutic Potential of Antibodies to GlcCer: in Vitro and in Vivo Antifungal Models
The relative inefficacy of the currently available antifungal drugs particularly in immunosuppressed individuals led several groups to search for new alternatives to control fungal infections. In this context, glycosphingolipid synthesis has been proposed as an attractive target of new antifungal drugs (Dickson et al. 2006).
More recently, it has been demonstrated that glycolipids can also be targeted by antimicrobial antibodies (Barreto-Bergter et al. 2004; da Silva et al. 2004; Nimrichter et al. 2004; Pinto et al. 2002; Rodrigues et al. 2007b; Rodrigues et al. 2000). As discussed below, the control of fungal infections by anti-glycosphingolipid antibodies may represent a new and promising perspective within the field of antimicrobial therapy.
Passive immunization with monoclonal antibodies (mAbs) to fungal antigens can protect mice against lethal mycoses. Targets of protective mAbs include capsular polysaccharides (Taborda et al. 2003), β-glucan (Rachini et al. 2007) and melanin (Rosas et al. 2001) of C. neoformans, mannoproteins (Moragues et al. 2003) and heat shock proteins (Burnie and Matthews, 2003) of Candida, histones of Histoplasma capsulatum (Nosanchuk et al. 2003), and glycoproteins of Aspergillus fumigatus (Chaturvedi et al. 2005) and P. brasiliensis (Buissa-Filho et al. 2008). The mechanisms of mAb-mediated protection are complex and heterogeneous, but it appears that passively administered antibodies enhance the effectiveness of host immune cells (Feldmesser and Casadevall, 1998) or act by a direct antifungal effect (Moragues et al. 2003; Nimrichter et al. 2004; Rachini et al. 2007; Rodrigues et al. 2000; Rosas et al. 2001). Some of the potentially therapeutic anti-fungal mAbs are in clinical trial for the treatment of cryptococcosis and candidiasis (Larsen et al. 2005; Matthews et al. 2003).
GlcCer has recently been described as a lipid component of secretory vesicles that carry a multiple payload of molecules that interfere with the immune function (Rodrigues et al. 2008; Rodrigues et al. 2007a). Although several lipids, including GlcCer, are believed to be poor immunogens (Bertini et al. 2007), antibodies to GlcCer can be isolated from sera of patients with cryptococcosis and other mycoses, but not from sera from healthy individuals (Rodrigues et al. 2000). The use of these antibodies as cell biology tools led to the conclusion that GlcCer is found at the surface of C. neoformans cells accumulating at the neck between the mother cell and the emerging daughter cell (Rodrigues et al. 2000). This observation suggested a connection between glycosphingolipid expression and fungal growth that was further confirmed by molecular biology approaches (Rittershaus et al. 2006). The cell wall has been shown to be the major cell site of GlcCer accumulation in fungal cells, and binding of these antibodies to the C. neoformans surface resulted in a strong inhibition of yeast proliferation (Rodrigues et al. 2000) (Fig. 2).
Fungal GlcCer is synthesized at the Golgi apparatus and then supposedly sorted to the endoplasmic reticulum or to the cell surface. Right panels show that GlcCer located at the cell wall and/or the plasma membrane are potential targets of anti-GlcCer antibodies and defensins; association between GlcCer and the peptide RsAFP2 initiates a signaling cascade culminating in ROS production in cellular sites that remain to be established. A similar response would be expected for anti-GlcCer antibodies.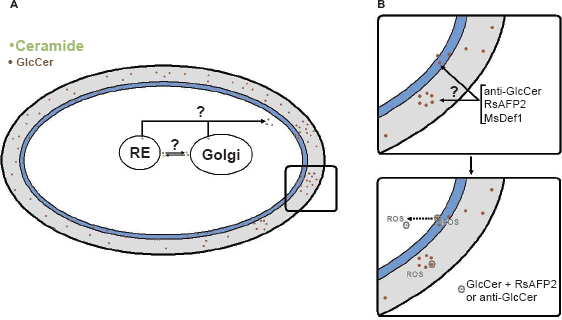
Analysis of antibody-treated C. neoformans cells by transmission electron microscopy revealed intense cellular damage, with organelle destruction, membrane retraction, and increased vacuolization (Nimrichter et al. 2005b) (Fig. 3). These results motivated the use of anti-GlcCer antibodies in other in vitro models of human and plant fungal pathogens. Studies directed to examine the antimicrobial effects of antibodies to GlcCer demonstrated that P. boydii and C. gloeosporioides conidia failed to differentiate into germ tube-like structures in the presence of anti-GlcCer antibodies (da Silva et al. 2004; Pinto et al. 2002). Similarly, pseudohyphae formation in C. albicans was affected by antibodies to GlcCer (Pinto et al. 2002). In F. pedrosoi, treatment of conidia with an anti-GlcCer mAb killed at least 60% of the conidial population (Nimrichter et al. 2004). The anti-GlcCer antibody also opsonized F. pedrosoi conidia and rendered fungal cells more susceptible to destruction by mouse macrophages. These results demonstrated that, besides their immediate antifungal action, GlcCer antibodies can help host cells to eliminate internalized fungi. In F. pedrosoi, the recognition of glucosylceramides by specific antibodies, as well as the fungal sensitivity to these mAbs, is influenced by melanin (Nimrichter et al. 2005a), which seems to negatively modulate antibody binding to the surface of conidia, mycelium and sclerotic cells.
Analysis of C. neoformans cells from normal growth (A) or after treatment with an antibody to GlcCer (B) as seen by transmission electron microscopy. Exposure to the antibody causes evident damage to yeast cells. Images provided by Drs. Kildare Miranda and Anderson J. Franzen. For experimental details, see Rodrigues et al. 2000.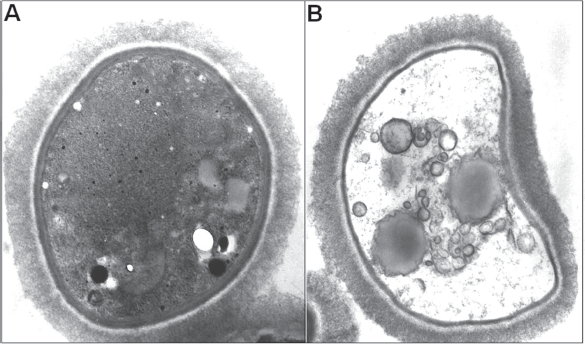
The results described above support the idea that fungal glucosylceramides could represent promising targets of antimicrobial compounds. Based on the previously characterized ability of antibodies to GlcCer to control the growth of fungal cells in vitro, a GlcCer-binding mAb was tested in vivo in a mouse model of cryptococcosis (Rodrigues et al. 2007b). Anti-GlcCer mAb protected mice against lethal C. neoformans infection (Rodrigues et al. 2007b). This study demonstrated for the first time that passive immunization with a lipid-binding antibody protected mice against a lethal fungal infection. Although antibodies to GlcCer were highly toxic for fungal cells in vitro (Nimrichter et al. 2005b), in vivo data showed that fungal loads were not different in mice receiving mAb to GlcCer, PBS or irrelevant antibody (Rodrigues et al. 2007b). This suggested that direct fungal cytotoxicity by the mAb was minimal during infection. However, in comparison to controls, mice given mAb to GlcCer had a dramatically reduced lung inflammatory response suggesting that the mAb-mediated protection could be a consequence of down-regulation of the inflammatory response. It is speculated, therefore, that in the presence of antibodies to GlcCer, the inflammatory response is better controlled by the host, resulting in reduced damage to host tissues and more effective killing of the invading organism by effector cells (Casadevall and Pirofski, 2003). Similar results were described in mice passively immunized with a protective IgG1 to polysaccharides prior to infection with C. neoformans (Rivera and Casadevall, 2005).
Cytokine levels were measured in control animals or mice given the antibody to GlcCer. The reduced inflammation in mice treated with the mAb to GlcCer corresponded to the increased lung concentration of anti-inflammatory cytokines such as IL-4 and IL-6 (Rodrigues et al. 2007b). It has been shown that IL-4 is necessary for mAb-mediated protection and that mice genetically deficient in IL-6 and IL-12 production have increased susceptibility to C. neoformans infections (Beenhouwer et al. 2001). In addition, an association between high levels of IL-6 and resistance to C. neoformans infection has been recently described (Siddiqui et al. 2006). Administration of anti-GlcCer mAb to lethally infected mice also increased lung expression of other cytokines that were previously associated to protection against C. neoformans. For example, the pulmonary levels of IL-12 increased in animals given the GlcCer antibody (Rodrigues et al. 2007b). In fact, IL-12 has been shown to have a protective effect against pulmonary infections by C. neoformans (Kawakami et al. 1996). These results led to the conclusion that, although it is not clear whether the anti-GlcCer antibody has a direct antifungal activity in vivo, it does influence the inflammatory response through the regulation of cytokine/chemokine expression, which is apparently related to host resistance against lethal cryptococcosis in mice.
GlcCer and Antifungal Peptides: Signaling Pathways Activating Cell Death
Anti-GlcCer antibodies are not the exclusive molecules that can bind to GlcCer and impair fungal growth. In 2004, Thevissen et al. showed that the antifungal peptide RsAFP2, a defensin purified from radish seeds, was fungicidal (Thevissen et al. 1999; Thevissen et al. 2004). A series of experiments revealed that the cell target of the defensin was GlcCer (Thevissen et al. 2004). Interestingly, mammalian GlcCer was not recognized by the peptide. In addition, the GlcCer-lacking S. cerevisiae was resistant to the antifungal effects of the peptide.
Exposure to RsAFP2 results in morphological changes, such as multiple budding and swelling of fungal cells (Osborn et al. 1995). It has also been demonstrated that plant defensins induce hyperpolarization of membrane potential (Caaveiro et al. 1997) and membrane permeabilization (Carrasco et al. 1981) in vitro. Although these are major mechanisms by which antimicrobial peptides act, it is not clear whether they sufficiently explain why RsAFP2 exerts its antifungal effects. Plant defensins, such as RsAFP2, do not form ion-permeable pores or directly change the electrical properties of artificial membranes indicating, possibly, a receptor mediated mechanism (Thevissen et al. 1996). To study the mechanism by which RsAFP2 inhibits fungal growth Aerts et al. (Aerts et al. 2007) evaluated its permeabilization capacity on carboxyfluorescence (CF)-containing small unilamellar vesicles (SUVs) enriched with purified fungal GlcCer (Aerts et al. 2007). At concentrations above that used to inhibit fungal growth the defensin was not able to cause CF leakage. These results suggest that, most probably, association of RsAFP2 with fungal GlcCer activates signaling pathways leading to cell death rather than causing direct membrane permeabilization. Further experiments, in fact, revealed that the RsAFP2-GlcCer association activates the production of endogenous reactive oxygen species (ROS) by C. albicans (Aerts et al. 2007). Accordingly, yeast cells lacking Cer:GlcT expression showed no differences in ROS formation before and after exposure to RsAFP2. The capacity of RsAFP2 to induce ROS was dose-dependent and the addition of ascorbic acid, an antioxidant molecule, to the medium abrogated its antifungal effect (Aerts et al. 2007). These results reveal a new mechanism by which a lipid-binding molecule can control the growth of fungal cells (Fig. 2).
Another plant defensin, MsDef1, was purified from alfafa and shown to cause hyperbranching of Fusarium graminearum (Spelbrink et al. 2004), a relevant phytopathogen that causes head blight disease in wheat, barley and other small grains (Bai and Shaner, 2004). Mutants hypersensitive to MsDef1 were generated and indicated that the MAP kinase proteins Mgv1 and Gpmk1 are directly involved with MsDef1 activity (Ramamoorthy et al. 2007b). These MAP kinase proteins are members of a family of serine/ threonine protein kinases that operate activating transcription factors involved in multiple developmental processes such as cell wall integrity, sexual reproduction and pathogenicity (Ramamoorthy et al. 2007a; Xu, 2000). Interestingly, these mutants were also hypersensitive to RsAFP2. This result led the authors to investigate the involvement of GlcCer in MsDef1-mediated signaling. A null mutant lacking the GCS gene, the one encoding Cer:GlcT, in F. graminearum was generated and demonstrated to be resistant to MsDef1, confirming that GlcCer is indeed the target of different plant peptides (Ramamoorthy et al. 2007a).
The data discussed in this section demonstrated the capacity of GlcCer ligands to induce fungal death through previously unknown mechanisms. Although it is clear that binding of external ligands to GlcCer can activate signaling molecules, it remains unknown if such process occurs at the plasma membrane or the cell wall. Further studies must be performed to precisely determine the cellular site where signaling pathways are triggered after the association of GSL to their ligands.
Plant and Animal Responses to Fungal GlcCer
As mentioned before in this review, GlcCer are present in fungal vesicles secreted in vitro and, possibly, in vivo (Rodrigues et al. 2007a). This fact, together with the confirmation that GlcCer is present at the cell wall of fungal pathogens, suggest that these molecules can constantly interact with host cells during infection (Nimrichter et al. 2004; Nimrichter et al. 2005b; Rodrigues et al. 2007a; Rodrigues et al. 2000). In fact, GlcCer from different fungal pathogens stimulate humoral responses (da Silva et al. 2004; Nimrichter et al. 2004; Pinto et al. 2002; Rodrigues et al. 2000). In other models, bacterial glycolipids such as the lipopolysaccharide from Campylobacter jejuni can induce the production of antibodies that cross-react with mammalian GSL, causing a special type of “autoimmune” disease (Yuki, 2007). A series of studies reported on the role of fungal GlcCer in plant infections (Koga et al. 1998; Umemura et al. 2002; Umemura et al. 2000). Glucosylceramides from M. griseae elicit a hypersensitive response in rice inducing the production of H2O2 and microbicidal proteins. The presence of insaturation on C4 and the methyl group at C9 are required for this response. Recently, it was demonstrated that fungal GlcCer stimulates the synthesis of NO and H2O2 by Taxus yunnanensis (Wang et al. 2007). This activation culminated with the production of high levels of taxol, a potent alkaloid, by a mechanism regulated by NO.
Fungal GlcCer also appear to modulate cellular responses in other eukaryotic cells. Kinoshita et al. demonstrated that GlcCer from the yeast Saccharomyces kluyveri regulates melanin synthesis in animal cells (Kinoshita et al. 2007). The yeast GlcCer suppresses the expression of tyrosinase in melanoma cells decreasing substantially their melanin content. An opposite effect was observed when these tumor cells were incubated with animal GlcCer. Previous data demonstrated that metabolites of GSL seem to regulate melanin production. For instance, ceramide and sphingosine-1-phosphate negatively regulates melanin synthesis (Kim et al. 2003) whereas sphingomyelin enhances melanin production (Higuchi et al. 2003). Also, a mislocalization of tyrosinase in glycosphingolipid-deficient GM95 mouse melanoma cell line was described with consequent loss of pigmentation (Sprong et al. 2001). Transfection of tyrosinase with an extended transmembrane domain, thus bypassing the transport block, restored pigmentation. Altogether these results indicate that lipid moieties or intact GSL participate directly on melanin regulation. So far, the correlation between GlcCer and fungal melanization has been suggested in two models. First, in C. neoformans, GlcCer-containing vesicles carry laccase, the enzyme responsible for triggering melanin polymerization in this organism (Rodrigues et al. 2008). Second, in F. pedrosoi, the surface expression of GlcCer is masked by the presence of melanin (Nimrichter et al. 2005a). The molecular mechanisms establishing a relationship between the synthesis of cerebrosides and pigments, however, are not known. Tyrosinase is not the only enzyme regulated by fungal GlcCer. Cerebrosides from Ganoderma lucidum, a basidiomycete, selectively inhibited the activities of replicative DNA polymerases from different eukaryotic species suggesting that fungal GlcCer can be involved in different aspects of eukaryotic cell regulation (Mizushina et al. 1998).
Several of the fungal species mentioned in this work are associated to neurological diseases, including C. neoformans, C. albicans and others. As discussed previously, GlcCer is the building block for the synthesis of complex GSL in animal cells, especially in neuronal cells where gangliosides are expressed in higher concentrations (Schengrund, 1990). Gaucher's disease is a GSL storage illness that occurs due to a mutation on glucocerebrosidase, the enzyme that hydrolyzes GlcCer to ceramide, with a consequent accumulation of cerebrosides in lysosomes (Jmoudiak and Futerman, 2005). During the acute neuronopathic effect, a severe loss of neurons is observed in the central nervous system. The incubation of neuronal cells with condritol-B-epoxide, an inhibitor of glucocerebrosidase, caused a similar accumulation of GlcCer. The consequence of higher levels of GlcCer, that can be transported to the ER (Trinchera et al. 1991), is the increase in ER density and amplified release of [Ca2+] in response to glutamate or caffeine (Korkotian et al. 1999). Although there is no experimental evidence showing that fungal GlcCer interfere with neurological functions, it has been shown that plant cerebrosides disturb Ca++-mediated cellular responses (Kurosu et al. 2007). A possible explanation could be an ionophore-like activity of GlcCer. In fact, the authors demonstrated that GlcCer from soy is able to bind Ca++ resulting in an increase of Ca++ permeability across the membrane. Removal of the hydroxyl group from the fatty acid of GlcCer from soy cerebrosides decreased both metal binding and membrane Ca++ permeability. The ion-cerebroside complex involves oxygen from the amide group, OH-C2 of fatty acid and OH-C2 of the sugar residue (Kurosu et al. 2007). The similar conformation of fungal GlcCer suggests that also in this case the same ionophore-like activity can be functional. Considering the ability of fungal pathogens that colonize the brain to release GlcCer-containing vesicles, such mechanism could interfere with the physiology of neuronal cells, among others (Rodrigues et al. 2008). The possibility that fungal GlcCer activates signaling mechanisms cannot be ruled out, since membrane architecture can be significantly modified by GlcCer accumulation.
Perspectives
Monohexosyl sphingolipids (CMHs) have once been regarded as membrane filling-space components thus participating in the construction of a unified structure mainly built up with proteins and phospholipids. Their polar groups were considered too short to participate in cellular interactions and no mechanisms similar to CD1 presentation of glycolipid molecules applied to most CMHs with the notorius exception of marine-sponge- derived α-galactosylceramide. Any effects related to glucosylceramides in fungi were usually attributed to a plasma membrane target event until there was evidence that the cell wall structure was mainly affected. The present review shows that fungal glucosylceramides and their counterparts in other eukaryotic cells play roles in enzyme distribution and function, are key components in cell division and differentiation and mediate signaling and cytotoxicity based on ROS. Many of these effects were unraveled by using specific anti-GlcCer antibodies and peptides. The path is open for in depth investigation on the role of fungal GlcCer as a key element in cell signaling and enzyme modulating activities. Since most GlcCer has been found in the fungal cell wall, studies should be directed to determine its role in the functional activity of vesicles, transient and secreted, the generation of ROS and signaling kinases on the cell wall, and in the complex machinery of cell division and differentiation. These propositions are in line with the most recent literature that recognizes the fungal cell wall as a dynamic structure rather than a rigid protective coating of the cell.
Footnotes
Acknowledgements
L.N., M.L.R. and E.B.B. are supported by grants from Conselho Nacional de Desenvolvimento Tecnológico (CNPq, Brazil) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil). L.R.T. is supported by CNPq and FAPESP (Brazil). The authors are indebted to Kildare Miranda and Anderson Franzen for preparation of TEM images.
The authors report no conflicts of interest.