
Abstract
Fungal cells are encaged in rigid, complex cell walls. Until recently, there was remarkably little information regarding the trans-fungal cell wall transfer of intracellular macromolecules to the extracellular space. Recently, several studies have begun to elucidate the mechanisms that fungal cells utilize to secrete a wide variety of macromolecules through the cell wall. The combined use of transmission electron microscopy, serology, biochemistry, proteomics and lipidomics have revealed that the fungal pathogens Cryptococcus neoformans, Histoplasma capsulatum, Candida albicans, Candida parapsilosis and Sporothrix schenckii, as well as the model yeast Saccharomyces cerevisiae, each produces extracellular vesicles that carry lipids, proteins, polysaccharides and pigment-like structures of unquestionable biological significance. Compositional analysis of the C. neoformans and H. capsulatum extracellular vesicles suggests that they may function as ‘virulence bags’, with the potential to modulate the host-pathogen interaction in favor of the fungus. The cellular origin of the extracellular vesicles remains unknown, but morphological and biochemical features indicate that they are similar to the well-described mammalian exosomes.
Insights into Secretory Mechanisms and Vesicular Transport Systems
Pathogenic mechanisms and secretory processes in microbes are closely associated. Secreted virulence factors and potent extracellular immunogens have been described for a number of prokaryotic and eukaryotic pathogens (Engel et al. 1998; Ghannoum, 1998; Hoegl et al, 1996; Hube, 1996; Liu and Nizet, 2004; McFadden et al. 2006a; Mitchell, 2006; Rodrigues et al. 2008; Rodrigues et al. 2007). Therefore, the understanding of secretory mechanisms and their regulation in microbial pathogens may represent a promising strategy for the design of new drugs and prophylactic agents.
There are numerous secretory pathways in eukaryotic and prokaryotic organisms that utilize diverse mechanisms for secretion (Gorelick and Shugrue, 2001; Lee et al. 2004; Mazar and Cotter, 2007; Rigel and Braunstein, 2008; Salama and Schekman, 1995; Wickner and Schekman, 2005). In eukaryotes, the most well-studied pathway of protein secretion involves vesicular migration from the endoplasmic reticulum to the trans face of the Golgi and then loading into a complex network of vesicles, the trans-Golgi reticulum (Glick and Malhotra, 1998). These proteins are sorted in the trans-Golgi network into transport vesicles that immediately move to and fuse with the plasma membrane, releasing their contents by exocytosis (Glick and Malhotra, 1998). In this regard, the generation of a collection of temperature-sensitive mutant Saccharomyces cerevisiae yeast strains (sec mutants in which secretion and cell surface assembly of proteins were blocked at different steps of the secretory pathway) was extremely important for the elucidation of the sequential events required for secretion (Novick et al. 1980; Novick and Schekman, 1979; Schekman, 2002; Schekman et al. 1983; Schekman and Novick, 2004). In these cells, inhibition of protein secretion at high (non-permissive) temperature is usually accompanied by morphological and biochemical changes, as well as intracellular vesicle accumulation. Other so-called ‘conventional’ mechanisms of secretion involve, for instance, ATP binding cassette type transporters, which are common to both eukaryotes and prokaryotes (Davidson and Maloney, 2007; Niimi et al. 2005).
Proteins that do not use the classical ER-Golgi pathway or membrane transporters can be secreted through various nonclassical pathways, as recently reviewed by (Nickel and Seedorf, 2008). Non-classical protein secretion may require vesicle release to the extracellular space, in a process that involves the formation of the so-called exosomes. During exosome biogenesis, small vesicles are formed by membrane invagination within endocytic compartments (endosomes). The formation of internal vesicles in the lumen of endosomes generates the so-called multivesicular bodies, which usually fuse with lysosomes in degradation pathways. However, multivesicular bodies can also fuse with the plasma membrane, resulting in the release of internal vesicles to the extracellular milieu as exosomes (Keller et al. 2006).
In contrast to most eukaryotic cells, fungi and bacteria are cell wall-containing organisms, making secretion topologically more complex. The presence of the cell wall, at the very least, implies the existence of trans-cell wall mechanisms for the release of molecules to the extracellular space. In prokaryotes, the mechanisms of transport of proteins across the cell wall are multiple. A general protein secretion pathway involving multiple genes (SECA, SECY, SECE, and SECG) and a number of accessory proteins has been firmly documented (Mori and Ito, 2001; Veenendaal et al. 2004). In Gram-negative bacteria, secretion systems have been widely reported (reviewed in (Saier, 2006)). This secretion machinery operates to release proteins extracellularly and, in the case of some pathogens, to inject toxins within host cells. At least six types of bacterial secretion systems have been defined, i.e. type I to type VI. Comprehensive reviews on this topic are available in the literature (Bingle et al. 2008; Cianciotto, 2005; Craig and Li, 2008; Henderson et al. 2004). Other protein secretion pathways in bacteria include two-partner secretion systems (Mazar and Cotter, 2007), which export large exoproteins across the outer membranes using channel-forming β-barrel proteins, twin-arginine transporters (Sargent, 2007), a membrane-bound transport apparatus that translocate proteins in a fully folded conformation, and secretion of extracellular vesicles (Mashburn-Warren et al. 2008). Vesicle-mediated toxin delivery is a potent virulence mechanism exhibited by diverse Gram-negative pathogens.
Trans-cell wall secretion in fungi has remained poorly explored for many years. Although it is well known that fungal cells secrete molecules of different chemical natures and molecular masses, the mechanisms by which extracellular structures leave intracellular compartments and cross the cell wall are virtually unknown. As typical eukaryotic organisms, fungal cells use conventional pathways of secretion involving post-Golgi vesicles that fuse with the plasma membrane to release their cargo (Schekman, 2002; Schekman and Novick, 2004). In fact, it is well known that yeast cells continuously secrete a number of enzymes that remain localized in the periplasm (Wickner and Schekman, 2005). The discovery that the capsular polysaccharide of C. neoformans had a molecular weight that could exceed 1 million Daltons (McFadden et al. 2006b), however, revealed the need for considering new mechanisms of trans-cell wall transport mechanism that could deliver macromolecules from the periplasmic space outside of the cell.
Recent studies reported the characterization of extracellular vesicles in pathogenic and non-pathogenic species of fungi. C. neoformans, Histoplasma capsulatum, Candida albicans, Candida parapsilosis, Sporothrix schenckii and S. cerevisiae were demonstrated to produce extracellular vesicles containing lipid, polysaccharide and protein components (Albuquerque et al. 2008; Rodrigues et al. 2008; Rodrigues et al. 2007). Therefore, extracellular vesicle secretion may represent a eukaryotic solution to the problem of trans-cell wall transport. Remarkably, the vesicles produced by C. neoformans and H. capsulatum contain key virulence determinants (Albuquerque et al. 2008; Rodrigues et al. 2008; Rodrigues et al. 2007), suggesting that, as described for bacteria (Mashburn-Warren et al. 2008), extracellular vesicles in fungi may represent an efficient mechanism of virulence factor delivery that may be crucial for the success of the infection. In this review, we discuss different models of extracellular vesicle secretion, as well as putative pathways of biogenesis and the impact of vesicle excretion on fungal pathogenesis.
Extracellular Vesicles and Trans-Cell Wall Transport: The Cryptococcus Neoformans Model of Polysaccharide and Protein Export
The most distinctive characteristic of the yeast pathogen C. neoformans is the expression of a polysaccharide capsule, a common feature of prokaryotic pathogens which is usually not observed in eukaryotic microbes. Another particularity of C. neoformans is the fact that the synthesis of capsular polysaccharides occurs in the cytoplasm (Feldmesser et al. 2001; Garcia-Rivera et al. 2004; Yoneda and Doering, 2006). In prokaryotes, capsule synthesis usually occurs at surface and extracellular sites. In Escherichia coli, for instance, capsular polysaccharides are initially assembled by enzymes associated with the plasma membrane (Whitfield, 2006; Whitfield et al. 2003).
The capsule of C. neoformans is primarily composed of two polysaccharides, namely glucuronoxylomannan (GXM) and galactoxylomannan (GalXM) (McFadden et al. 2006a). GXM, the best studied capsular component of C. neoformans, is the main virulence factor in this pathogen (McClelland et al. 2005). GXM is not only surface associated, but also continuously secreted to the extracellular space where it accumulates in tissues (McFadden et al. 2006a). Interestingly, GXM has a molecular mass in the range of 1 to 7 million Daltons (McFadden et al. 2006b), suggesting the existence of mechanisms of transport of the polysaccharide from intracellular sites to the extracellular environment. Early and recent studies suggested that the transfer of intracellular polysaccharides to the extracellular space in fungi could require vesicular transport.
Vesicular transport in fungi was hinted at in several earlier reports. Heath and colleagues showed almost 40 years ago that fungal vesicles contained cell wall precursors, suggesting that intracellular vesicles can migrate to the cell surface to release substrates for wall synthesis (Heath et al. 1971). More recently, the cell wall of C. neoformans was described as the major cellular site of the glycosphingolipid glucosylceramide (Rodrigues et al. 2000), which is a membrane component of vesicles that migrate from the plasma membrane to the cell wall (Nimrichter et al. 2005b; Rodrigues et al. 2007; Rodrigues et al. 2000). Early studies by Takeo and co-workers also demonstrated that C. neoformans secrete vesicles outside the cell membrane (Takeo et al. 1973a, b). Interestingly, despite the fact that no molecular evidence was provided at that time, the authors proposed that “capsular material is synthesized in and released via the vesicles”. There the story rested for almost 20 years, until Feldmesser et al. used immunogold electron microscopy and noticed clumped labeling of the cell wall consistent with vesicular transport of GXM (Feldmesser et al. 2001). Those observations were further extended by that of Garcia-Rivera and colleagues who showed that antibodies to GXM recognized cytoplasmic and cell wall clusters resembling vesicles (Garcia-Rivera et al. 2004). In essence, these studies provide support for the suggestion that the capsular polysaccharide was synthesized in the cytoplasm and exported to the exterior of the cell in secretory vesicles that traversed the cell wall.
Based on the classical studies of Schekman and Novick on yeast mutants defective in protein synthesis (Novick et al. 1980; Novick and Schekman, 1979; Schekman, 2002; Schekman et al. 1983; Schekman and Novick, 2004), Yoneda and Doering (Yoneda and Doering, 2006) used site-directed mutagenesis to generate a C. neoformans strain defective in the production of Sav1p, a homolog of the S. cerevisiae small GTPase Sec4p. In S. cerevisiae, sec4 mutants accumulate post-Golgi vesicles under restrictive conditions (Walworth et al. 1989), a morphological feature that was also observed in the C. neoformans mutant (Yoneda and Doering, 2006). More precisely, the sav1 mutant of C. neoformans had defective protein secretion and accumulated exocytic vesicles at the septum and the bud during cell division. Strikingly, these vesicles were strongly recognized by an antibody to GXM (Yoneda and Doering, 2006), confirming and extending previous suppositions that the polysaccharide was synthesized intracellularly and secreted in vesicles.
Since the capsule of C. neoformans enlarges by apical growth (Zaragoza et al. 2006), one might deduce that capsular components are secreted to the extracellular space and incorporated into the external layers of the growing capsule. Therefore, assuming that secretion of intracellularly synthesized GXM is an essential element of capsular growth, one might hypothesize that extracellular vesicles filled up with capsular components may exist in C. neoformans culture supernatants. Indeed, cell wall-bound vesicles, as well as extracellular vesicles in association with surface structures, were observed in vitro and during animal infection by C. neoformans (Rodrigues et al. 2007). In addition, the fractionation of culture supernatants using centrifugation procedures previously designed for the purification of secreted vesicles in mammalian systems allowed the isolation of extracellular lipid-containing fractions. The analysis of these fractions by transmission electron microscopy revealed the presence of round vesicles, in the range of 20 to 400 nm, containing bilayered membranes (Rodrigues et al. 2008; Rodrigues et al. 2007). Vesicle morphology included usually round compartments with different levels of electron-density. Vesicular structures with hyper-dense structures resembling a dark pigment were also observed. Importantly, some of the isolated vesicles were recognized by an antibody to GXM (Rodrigues et al. 2007), confirming that these compartments were involved in polysaccharide trans-cell wall secretion. In addition, GXM detection in vesicles was correlated with capsule enlargement, suggesting that polysaccharide delivery in extracellular vesicles is necessary for assembly of the capsule (Rodrigues et al. 2007).
The production of extracellular vesicles requires cell viability (Rodrigues et al. 2007), confirming that these compartments are actively secreted rather than passively released by dead cells. Transmission electron microscopy of intact C. neoformans cells also revealed that the secretory vesicles were recognized by an antibody to glucosylceramide (Rodrigues et al. 2007). Since glucosylceramide is an important regulator of growth and virulence in C. neoformans (Rittershaus et al. 2006; Rodrigues et al. 2000), the possibility that the extracellular vesicles carry bioactive lipids was raised. The analysis of lipid components of isolated extracellular vesicles by chromatographic and spectrometric methods revealed the presence of glucosylceramide (GlcCer) and other structures (Rodrigues et al. 2007). By thin-layer chromatography, bands with migration rates corresponding to GlcCer and ergosterol were detected in lipid extracts of vesicles produced by both acapsular and encapsulated C. neoformans cells. Vesicle lipids from encapsulated cells were also examined by electrospray ionization mass spectrometry, revealing a complex lipid composition. GlcCer analysis in vesicle fractions revealed the presence of N-2'-hydroxyhexadecanoyl- and N-2'-hydroxyoctadecanoyl-1-beta-D-glucopyranosyl-9-methyl-4,8-sphingadienine. Analysis of sterols showed that ergosterol and 4,14-dimethylergosta-24(241)-en-3β-ol, an obtusifoliol-like molecule, are also vesicle components in C. neoformans (Rodrigues et al. 2007).
In addition to polysaccharides and lipids, the extracellular vesicles of C. neoformans also carried a complex array of proteins to the extracellular milieu. Employing a proteomic approach, 76 different proteins were identified as vesicle components (Rodrigues et al. 2008). Chaperones, including heat shock proteins and superoxide dismutase, signal transduction regulators, anti-oxidant and cytosolic proteins and enzymes were identified. It is intriguing that approximately one third of proteins identified in the C. neoformans vesicles corresponded to molecules that were previously described to compose mammalian exosomes (Rodrigues et al. 2008).
Vesicular Trans-Cell Wall Transport in Histoplasma Capsulatum and other Fungal Species
Following the identification of extracellular vesicles in the basidiomycetes C. neoformans, fungi in the phylum Ascomycota were examined for vesicle production. The ascomycetes H. capsulatum, C. albicans, C. parapsilosis, S. schenckii, and S. cerevisiae each were found to generate heterogeneous extracellular vesicles (Albuquerque et al. 2008). Transmission electron microscopy studies of supernatants subjected to fractional centrifugation revealed that each fungus produced extracellular spherical, bilayered vesicles that varied in their electron density and ranged in diameter from 10 to 350 nm. Vesicles were not present in supernatants from media inoculated with dead fungal cells. Notably, the percentage of large versus small vesicles varied significantly between the different ascomycetes. For example, only 4% of S. cerevisiae vesicles were larger than 50 nm compared to 38 and 54% of vesicles from H. capsulatum and C. parapsilosis, respectively.
The movement of membrane-bound vesicles across C. albicans cell walls was previously proposed by Anderson and co-workers (Anderson et al. 1990) who demonstrated the presence of “pimples” in the cell wall of opaque switch variants of strain WO-1 where vesicles were identified within some channels or appeared to emerge from the “pimples” with their double membrane intact. The analysis of intact H. capsulatum yeast cells by transmission electron microscopy similarly demonstrated vesicular structures engaging the internal aspects of the cell wall, within the cell wall, and emerging from the cell wall consistent with trans-cell wall transport of vesicles (Albuquerque et al. 2008).
To further define the constituents of the ascomycetes vesicles, purified fractions from H. capsulatum were subjected to lipidomic and proteomic analyses (Albuquerque et al. 2008). Using electrospray ionization time-of-flight mass spectrometry, phosphatidylethanolamine, phosphatidylserine (PS), and phosphatidylcholine (PC) were identified as the major phospholipid species comprising the vesicles, which is similar to the typical distribution of lipids in pathogenic yeast (Rattray et al. 1975). The protein components of the vesicles were analyzed by cation exchange chromatography and analyzed by liquid chromatography-tandem mass spectrometry and 283 proteins were validated, with 206 proteins identified by sequence analysis. Notably, several proteins involved in H. capsulatum pathogenesis and host immune responses were detected, including chaperones (Hsp30, Hsp70, and Hsp60), superoxide dismutase, and catalase B. Furthermore, proteins involved in signal transduction, vesicle formation, cell wall and cytoskeleton regulation, cell growth, and sugar, lipid and amino acid metabolism were identified.
The fact that proteins associated with H. capsulatum virulence were identified in the purified extracellular vesicles raised the question of whether the vesicle proteins were recognized by the host immune system. In this regard, hyperimmune human serum reacted strongly with diverse vesicular proteins, including Hsp60 and histone 2B. Notably, H. capsulatum Hsp60 has been associated with virulence (Allendoerfer et al. 1996; Deepe and Gibbons, 2001; Deepe and Gibbons, 2002; Gomez et al. 1995b; Scheckelhoff and Deepe, 2002) and antibody to histone 2B can modify experimental histoplasmosis (Nosanchuk et al. 2003). Hence, it is likely that proteins transported via vesicles are involved in host-pathogen interactions.
Putative Mechanisms of Vesicle Diversity, Biogenesis and Passage through the Cell Wall
The findings of Yoneda and Doering (Yoneda and Doering, 2006) suggesting that GXM-containing vesicles in C. neoformans were derived from the Golgi apparatus was further supported by Hu and co-workers (Hu et al. 2007). These authors reported that exposure of C. neoformans to brefeldin A resulted in a massive reduction of capsule size. The target of brefeldin A in mammalian cells is a subset of Sec7-type GTP-exchange factors that catalyze the activation of a small GTPase called Arf1p (Nebenfuhr et al. 2002). Arf1, in turn, is responsible for the recruitment of coat proteins to membranes, resulting in the formation of transport vesicles (Scales et al. 2000). Arf1p and brefeldin A-sensitive GTP-exchange factors are localized to the Golgi apparatus of mammalian and yeast cells (Spang et al. 2001). Therefore, it seems clear that GXM assembly at the cell surface requires the secretion of post-Golgi vesicles filled up with the polysaccharide. These vesicles would be targeted to the plasma membrane for exocytosis (Yoneda and Doering, 2006), which would result in the release of the polysaccharide to the periplasmic space. Therefore, for capsule assembly, additional mechanisms of trans-cell wall transport of GXM would be required. However, a model of exocytosis with discharge into the periplasmic space did not account for transport of GXM across the cell wall for assembly into a capsule in the extracellular space.
Mutation of the SEC4 homolog that resulted in the cytoplasmic accumulation of post-Golgi vesicles did not lead to any apparent alteration in the capsular expression phenotype (Yoneda and Doering, 2006). This observation could be a consequence of continued polysaccharide secretion before the temperature shift, which blocks secretion, or incomplete inhibition of the secretion caused by the mutation. It was impossible to distinguish in C. neoformans whether GXM was targeted to the cell surface exclusively in post-Golgi vesicles or via recycling endosomes. In fact, endosomes and related multivesicular bodies (MVBs) can be connected to the trans-Golgi secretory pathway, so both pathways could be involved in polysaccharide secretion in C. neoformans. It is therefore possible that the post-golgi GXM-enriched vesicles are sorted to another compartment than the plasma membrane such as the late endosomes and the MVBs.
The existence of MVB-like structures in C. neoformans was suggested in early and recent studies (Rodrigues et al. 2008; Takeo et al. 1973a, b) (Fig. 1). Some of these compartments were found to be clearly merging with the plasma membrane, indicating that the extracellular vesicles of C. neoformans could be related to mammalian exosomes. Exosomes are extracellular vesicles derived from the fusion of MVBs with the plasma membrane. Their isolation was extensively described in several animal cell models such as reticulocytes, dendritic cells, B lymphocytes, mastocytes and epithelial cells (Johnstone et al. 1987; Lamparski et al. 2002; Raposo et al. 1996; Skokos et al. 2003; van Niel et al. 2001).
Typical MVB-like structures in fusion with the plasma membrane in C. neoformans. MVB internal vesicles (arrowheads) are released outside the external layer of the plasma membrane, representing a potential mechanism of exosome biogenesis in fungi. This figure was originally published by Takeo and colleagues (Takeo et al. 1973b). Reproduced with permission from the American Society for Microbiology. CW, cell wall; PM, plasma membrane; Cy, cytoplasm; MVB, multivesicular body.
The molecular machinery implied in MVB formation and sorting is widely known in S. cerevisiae (Hurley and Emr, 2006), but these studies had never shown extracellular exosomes in fungal cells. Early observations by Takeo and colleagues (Takeo et al. 1973a) suggested the occurrence of what the authors defined as an “early stage of formation of spherical invaginations which secrete the vesicles outside the cell membrane”, suggesting the occurrence of exosomes in fungi. That supposition was strengthened by the highly heterogeneous protein composition of C. neoformans and H. capsulatum vesicles (Albuquerque et al. 2008; Rodrigues et al. 2008). This heterogeneous protein profile is also seen in mammalian exosomes, indicating a possible common origin for both extracellular vesicles. Moreover, detergent-resistant lipid microdomains are known to compose exosome membranes at least in some mammalian cell types (de Gassart et al. 2003). Lipid microdomains–-or lipid rafts – are mainly characterized by high sterol content and presence of glycosphingolipids, as described in fungal vesicles (Rodrigues et al. 2007). They are seemingly involved on the sorting of exosomal proteins as well as in vesicle morphology and formation (de Gassart et al. 2003).
Laccase, an enzyme involved in melanin synthesis, is another important component of C. neoformans vesicles (Rodrigues et al. 2008). Interestingly, it was recently shown that a C. neoformans mutant defective in the vps34 gene has a considerable decrease on melanin expression (Hu et al. 2008). Vps34 (vacuolar protein sorting 34) is involved in vesicular sorting of vacuolar hydrolases from the trans-Golgi-network to the late endosomes and in the internal vesicle formation within MVBs (Futter et al. 2001; Kihara et al. 2001). These data strongly support the idea that laccase-containing vesicles originate from the endocytic pathway.
Although different lines of evidence point to a relationship between fungal extracellular vesicles and exosomes, other possibilities can not be ruled out. The methods currently used for vesicle purification do not discriminate between vesicles of different origins, resulting in heterogeneous preparations. Indeed, electron microscopy combined to other approaches demonstrated that vesicle fractions contain different subpopulations which differ in morphological characteristics (Rodrigues et al. 2008), suggesting the existence of different intracellular compartments involved in vesicle biogenesis (Fig. 2). Cellular vesicle secretion strategies other than exosome formation (Fig. 2B) could consist of simple membrane budding (Fig. 2C) and other unconventional mechanisms of vesicle release (Fig. 2D). The existence of different mechanisms of vesicle biogenesis could, in turn, result in the simultaneous generation of extracellular membrane compartments of different cellular origins.
Mechanisms for vesicle biogenesis in fungal cells. (A) Vesicles sorted to the cell surface by conventional secretion fuse with the plasma membrane, releasing their cargo into the periplasm. MVB formation followed by fusion with the plasma membrane (B) would result in the extracellular release of exosomes. Membrane budding (C) could also result in the periplasmic release of vesicles, as also proposed for other unconventional secretion mechanisms (D). For conceptual clarification, see (Kinseth et al. 2007) and (Keller et al. 2006). Of note, vesicle content could vary according with biogenesis pathways. The possibility that different mechanisms result in the simultaneous generation of extracellular vesicles cannot be ruled out.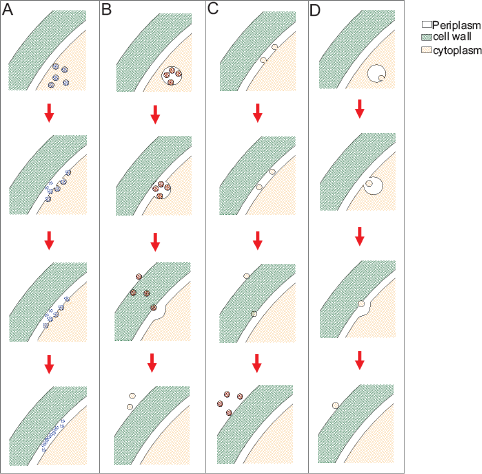
Further investigations at the molecular and ultra-structural levels will need to be done for the development of reliable concepts of vesicle biogenesis. In this context, the discovery of biochemical markers for different vesicle sub-populations is likely to be fundamental criterion for vesicular identification and characterization. In this scenario, genetic approaches will be powerful tools to understand where the vesicles are assembled and how they reach the extracellular space.
It is unknown how vesicles cross the cell wall to reach the extracellular space. Transmission electron microscopy revealed that the cell wall of C. neoformans and H. capsulatum contain numerous vesicle-like structures (Rodrigues et al. 2008; Albuquerque et al. 2008). The cell wall is now described as a compact although malleable structure (Nimrichter et al. 2005a), likely to suffer rearrangements, that could allow vesicle passage. Indeed, it was demonstrated by atomic force microscopy that the cell wall of fungi contain pores that would allow vesicle passage (de Souza Pereira and Geibel, 1999; Eisenman et al. 2005). Trans-cell wall vesicle secretion could also involve vesicle-mediated cell wall remodeling. For example, H. capsulatum extracellular vesicles contain enzymes regulating synthesis and hydrolysis of cell wall components (Albuquerque et al. 2008). In yeast cells, wall remodeling for vesicle passage could be facilitated during budding, when the cell wall is thinner in the bud than in non-dividing areas of the cell surface (Linnemans et al. 1977). Finally, a myosin analog was described as a cell wall component of Aspergillus fumigatus (Esnault et al. 1999). In the cytosolic space, motor proteins like myosin and dynein contribute with the movement of transport vesicles (Schliwa and Woehlke, 2003). However, it is completely unknown if such proteins would be functional at the cell wall microenvironment.
‘Virulence Bags’: Fungal Extracellular Vesicles Concentrate Fungal Molecules Involved in Pathogenesis
It is clear that fungal vesicles are released to the extracellular milieu carrying a complex panel of proteins and lipids and, in the case of C. neoformans, polysaccharides (Rodrigues et al. 2007; Rodrigues et al. 2008). Current data strongly suggest that vesicle secretion occurs not only in vitro, but also in vivo during animal infections by fungal pathogens. Secreted vesicles were detected when murine macrophages were incubated with C. neoformans yeasts and also in sections of lung excised from mice infected with this pathogen (Rodrigues et al. 2007). Vesicles secreted during infection could directly mediate host cell damage and/or modulation of immune response. Indeed, similar functions were proposed for vesicles released from the outer membrane of Gram negative bacteria (Mashburn-Warren et al. 2008).
Lipid components of fungal vesicles have the potential to modulate interactions of fungi with their hosts. GlcCer is required for fungal growth in alkaline pH and high CO2 levels, as well as for the virulence of C. neoformans (Rittershaus et al. 2006; Saito et al. 2006). The fact that the vesicles secreted by S. cerevisiae, which does not synthesize GlcCer, are in a smaller size range (Albuquerque et al. 2008) suggests an involvement of GlcCer in vesicle assembly. As a vesicle-associated compound, GlcCer could be constantly delivered to host cells, facilitating the production of antimicrobial antibodies (Rodrigues et al. 2000). In addition, lipid turnover is known to be a well regulated phenomenon that occurs in mammalian cells. Glycosphingolipids are also involved with sorting of lipids and proteins in mammalian cells (Degroote et al. 2004). Although fungal and mammalian GlcCer are structurally different (Barreto-Bergter et al. 2004), secreted fungal GlcCer could potentially interfere with recycling and sorting of lipids and proteins in host cells. Other lipids, including phosphatidylcholine, phosphatidylethanolamine and phosphatidylserine, were also characterized in fungal vesicles (Albuquerque et al. 2008). Although these are common components of biological membranes, their involvement in immune responses has been already reported. Liposomes carrying phosphatidylserine, for instance, can modulate cytokine production, decrease microbial killing and inhibit nitric oxide production by macrophages (Aramaki, 2000; Gilbreath et al. 1986; Hoffmann et al. 2005).
In bacteria, adhesins, toxins, and immunomodulatory compounds are the components of extracellular vesicles, which directly mediate bacterial binding and invasion, cause cytotoxicity, and modulate the host immune response (Kuehn and Kesty, 2005). In fungal vesicles, several proteins involved with fungal pathogenesis were identified (Albuquerque et al. 2008; Rodrigues et al. 2008). Concentration of pathogenic determinants in the vesicles could provide an efficient mechanism of release of virulence factors into host tissues. Vesicle membranes could also protect fungal structures against hydrolysis by host extracellular enzymes. In addition, membrane fusion involving vesicle elements and the surface of host cells could result in the direct delivery of fungal structures into host tissues.
Different immunogenic compounds are released in fungal vesicles. Chaperones, including heat shock proteins 60 and 70 from H. capsulatum and C. neoformans, respectively, were detected in extracellular vesicles by proteomics and serology (Albuquerque et al. 2008; Rodrigues et al. 2008). Hsp60 is responsible for binding of H. capsulatum to the CD11/C18 receptor in human macrophages (Long et al. 2003). This protein, which was initially described as a glycoprotein present at membrane and cell wall of H. capsulatum, confers protection against lethal intravenous challenge with this pathogen (Gomez et al. 1995a). Besides representing an efficient vehicle for delivery of immunogens during infection, vesicles carrying Hsp60 could sequestrate anti-Hsp60 antibodies produced by the host. In C. neoformans, the role of Hsp70 is still unknown, but this protein is considered to be a major immunogen in cryptococcosis (Kakeya et al. 1997; Kakeya et al. 1999).
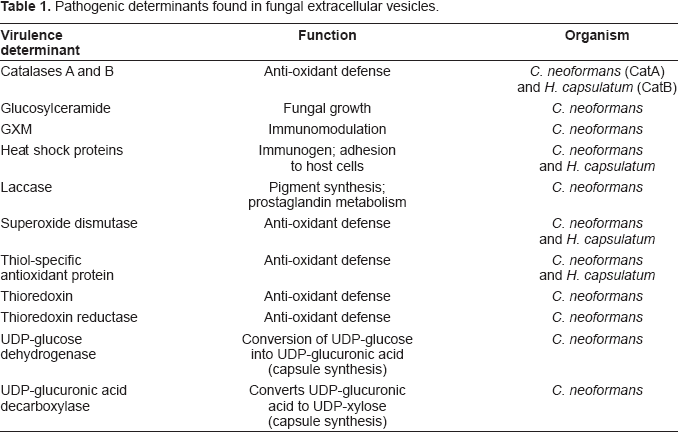
Pathogenic determinants found in fungal extracellular vesicles.
As discussed in previous sections, cryptococcal GXM is also secreted within vesicles that reach the extracellular space. GXM is recognized by different cell types and involved with a series of immunomodulatory effects (Vecchiarelli, 2005). Polysaccharide vesicular export, therefore, could be not only a physiological solution for capsule assembly, but also a fungal strategy to cause tissue damage through the delivery of virulence factors and defense proteins into host tissues. Interestingly, two different enzymes with well-defined roles in capsule synthesis and virulence were also found in cryptococcal vesicles (Rodrigues et al. 2008).
Unsolved Problems and Future Perspectives
The discovery of a trans-cell wall vesicular transport system in C. neoformans, and its subsequent extension to ascomycetous fungi, provides a solution to the problem of exporting macromolecules to the cellular exterior. The finding of similar vesicles in ascomycetes and basidiomycetes suggest that the vesicular transport system is ancient, such that its existence predated the divergence of these fungal branches 0.5–1.0 billion years ago. For C. neoformans the existence of such an export mechanism would appear to be necessary for capsular synthesis given this structure is composed of macromolecules with mass that can exceed 1 million Daltons (McFadden et al. 2006b). The finding that such vesicles contain numerous fungal products associated with virulence suggests that they also have a concentrative function that could allow these compounds to function more efficiently, and possibly synergistically. However, the discovery of this trans-cell wall vesicular transport system also poses a new set of questions for future studies.
How many types of vesicles are there?
Electron microscopy reveals that vesicles differ in shape, size, and inter-vesicular contents. EM surveys suggest the existence of at least four types of vesicles in C. neoformans supernatants (Rodrigues et al. 2008). Classifying vesicular types is important because each type would suggest the existence of a separate cellular synthetic mechanism. Currently, vesicle types have been discriminated based on morphology and size. However, morphological criteria may not be adequate to ascertain the various types of vesicles since similar appearance does not necessarily imply similar content. Consequently, continued progress in this area will require the development of methods for vesicular fractionation and chemical characterization. Furthermore, a full description of the vesicular transport system will require a correlation of vesicular content with morphological characteristics, which is likely to involve combined proteomic analysis, serological reactivity, enzymatic assays, etc.
Where do vesicles originate from?
Analysis of protein components and the finding of vesicles within vacuolar structures led to the proposal that extracellular vesicles originate in exosome-like structures (Rodrigues et al. 2008). However, such structures have yet to be described in fungi. Definition of the various vesicle types combined with mutational analysis could be used to ascertain the number of vesicular synthetic pathways in fungal cells. Understanding the sites of vesicular synthesis and the relationship of extracellular vesicles to cytoplasmic vesicles will undoubtedly be a major area of investigation in future years.
How are vesicles transported across the cell wall?
There is no information on the mechanism used to shuttle vesicles across the cell wall. The existence of pores on the cell wall with dimensions that approximate those of vesicles suggest that these could be used for vesicular transport. Vesicular transport across pores raises a variety of other questions including the energy dynamics of such processes and the types of motors and gears that would be needed to shuttle such large structures across rigid cell walls. This is made even more complex when one considers that certain fungi, such as C. neoformans, H. capsulatum and S. schenckii can form dense melanin layers within their cell walls (for review, (Nosanchuk and Casadevall, 2006)).
What is the fate of released vesicles?
In the case of C. neoformans, vesicular transport has been proposed as the mechanism to deliver macromolecules for capsular assembly to the exterior of the cell (Rodrigues et al. 2007). If that is the case, it raises the question of how vesicles are directed to the capsule surface and their cargo unloaded. Extracellular phospholipases may degrade phospholipids membranes and allow cargo discharge. C. neoformans and most pathogenic fungi have extracellular phospholipase activity (Chayakulkeeree et al. 2008; Ghannoum, 1998; Hruskova-Heidingsfeldova, 2008; Simockova et al. 2008). A related issue is the relative paucity of vesicles relative to the number of cells in solution. This could be interpreted as indicating that vesicles are relatively short lived structures that release their contents in the extracellular space. It is possible that vesicles recovered from supernatants reflect a very small minority of the secreted vesicles and that isolation of the overwhelming majority of vesicles produced by fungal cells would require the development of more sophisticated recovery techniques. Hence, understanding the dynamics of vesicular transport will require studies on the fate of released vesicles.
What is the relationship between vesicles and host cell toxicity?
C. neoformans intracellular pathogenesis is characterized by the appearance of numerous vesicles in the cytoplasm of phagocytic host cells that contain cryptococcal capsular polysaccharide (Feldmesser et al. 2000). These vesicles are believed to be toxic to the host cells since their accumulation is temporally correlated with cytotoxic changes that include membrane blebs and phagosomal membrane leakiness (Feldmesser et al. 2000; Tucker and Casadevall, 2002). The discovery of a vesicular transport system for C. neoformans raises the possibility that these cytoplasmic vesicles originated as fungal vesicles that were released from yeast-containing phagosomes. Hence, knowing about the effect of shed vesicles on host cells and their relationship to the polysaccharide-containing vesicles that accumulate in host cells would seem to be an important question for understanding the pathogenesis of cryptococcal infections. Furthermore, it is possible that shed vesicles are also involved in the pathogenesis of other facultative intracellular fungi such as H. capsulatum.
What is the internal organization of vesicular components?
Given that vesicles contain multiple fungal components that include lipids, polysaccharides, carbohydrates and proteins, it is possible that these are packaged inside vesicles in an organized manner that could reflect their addition following or during vesicular synthesis. For example, it is conceivable that vesicles destined for capsular synthesis contain capsular components arranged in a manner that facilitates their function. Understanding the packaging characteristics of vesicles is likely to be an essential aspect of understanding their function and mechanisms of action. Such studies will almost certainly require the application of new microscopy techniques in combination with specific reagents, such as monoclonal antibodies, that delineate the position of specific components within the vesicular structure.
Finally, there is the notion that knowledge of vesicular transport systems may be exploited in the design of new types of antifungal drugs and vaccines. The complex choreography of vesicular transport implies the existence of numerous steps that could be targets of drug development. One could imagine drugs that reduce the virulence of fungi by interfering with vesicular synthesis and transport. Identification of common pathways used by different pathogenic fungi for vesicular transport could lead to the design of broad-spectrum antifungal drugs. Since trans-cell wall vesicular transport mechanisms have no counterpart on mammalian cells such drugs could potentially have high specificity for fungal cells while concomitantly having limited host cells toxicity. Furthermore, it is possible that vesicular preparations can be developed into vaccine formulations. The finding of many fungal components associated with virulence in vesicles and the fact that vesicle-related proteins are immunogenic (Albuquerque et al. 2008; Rodrigues et al. 2008) suggest that vesicle preparations and/or their components could have utility as fungal vaccines. In this regard, it is noteworthy that exosome preparations from dendritic cells are been investigated as potential anti-cancer vaccines (Hao et al. 2007).
It is now apparent that the discovery of vesicles in cryptococcal culture supernatants has opened a new window into fungal cell biology that promises to revolutionize our thinking on fungal product export and its impact on pathogenesis. It is also clearly apparent that this area of investigation is extremely complex and will require the development of new experimental tools and reagents for continued progress.
Footnotes
Acknowledgements
MLR and LN are supported by grants from the Brazilian agencies FAPERJ and CNPq. AC is supported by NIH grants AI033142, AI033774, AI052733, and HL059842. JDN is supported by NIH AI52733 and AI056070-01A2. We are thankful to Igor C. Almeida, Rosana Puccia and Luiz R. Travassos for helpful discussions.
The authors report no conflicts of interest.