
Abstract
Stress plays an important role in drug- and addiction-related behaviours. However, the mechanisms underlying these behavioural responses are still poorly understood. In the light of recent reports that show consistent regulation of many genes encoding stress proteins including heat shock proteins following ethanol exposure in Drosophila, it was hypothesised that transition to alcohol dependence may involve the dysregulation of the circuits that mediate behavioural responses to stressors. Thus, behavioural genetic methodologies were used to investigate the role of the Drosophila hsp26 gene, a small heat shock protein coding gene which is induced in response to various stresses, in the development of rapid tolerance to ethanol sedation. Rapid tolerance was quantified as the percentage difference in the mean sedation times between the second and first ethanol exposure. Two independently isolated P-element mutations near the hsp26 gene eliminated the capacity for tolerance. In addition, RNAi-mediated functional knockdown of hsp26 expression in the glial cells and the whole nervous system also caused a defect in tolerance development. The rapid tolerance phenotype of the hsp26 mutants was rescued by the expression of the wild-type hsp26 gene in the nervous system. None of these manipulations of the hsp26 gene caused changes in the rate of ethanol absorption. Hsp26 genes are evolutionary conserved, thus the role of hsp26 in ethanol tolerance may present a new direction for research into alcohol dependency.
Keywords
Introduction
Alcohol consumption in humans causes long-term physiological changes including tolerance. Tolerance in this sense can develop to both the pleasurable (eg, euphoria and loss of social inhibition) and the aversive (eg, loss of motor coordination and sedation) effects of ethanol, 1 and is thought to encourage increased alcohol intake, development of physical dependence and addiction. 2 Indeed, alcoholics acquire extra-ordinary tolerance to the intoxicating effects of ethanol, usually associated with dependence and uncontrolled craving to continue drinking. 3
Tolerance is defined as a reduction in drug responsiveness seen after repeated exposures to that drug. There are believed to be different types of tolerance: Acute tolerance, which occurs within drug session/experience; rapid tolerance, which occurs after the completion of a single drug exposure/experience; and chronic tolerance which arises from serial drug exposures. 4 Further, two mechanisms of tolerance which are not mutually exclusive have been reported: metabolic/pharmacokinetic tolerance, involving changes in the disposition of ethanol (such as absorption, excretion or metabolism) leading to efficient removal of alcohol from the body and functional/ pharmacodynamic tolerance involving changes experienced at a cellular level and mediated by adaptations in neural function. 1
Drosophila has been shown to acquire rapid tolerance to the sedating effects of ethanol, 5 and in flies many genes have been implicated in ethanol tolerance. For example, flies carrying a mutation in the hangover (hang) gene (a gene encoding a nucleic acid binding zinc finger protein) were implicated in the development of reduced ethanol tolerance as were flies lacking the neuromodulator octopamine owing to a mutation in the gene encoding tyramine β hydroxylase (Tbh).5,6 Further, induction of tolerance was completely abolished in flies simultaneously carrying null mutations in hang and in Tbh, indicating that both genes are involved in different pathways in the induction of ethanol tolerance. 6 Flies carrying slowpoke, a Ca2+-activated K+ channel gene which is a critical modulator of neuronal excitability have also been shown to be required for the acquisition of tolerance to ethanol.7,8 The Drosophila homolog of JWA, a microtubule binding protein, was also reported to be necessary for the acquisition of ethanol tolerance in Drosophila. 9 Finally, a study of flies defective in long-term memory implicated several genes in reduced ethanol tolerance including exba (elF-5C), a translational regulator involved in axon guidance); pumilio (translational regulator) and formin3, involved in actin assembly. 10
Microarray experiments have been recently used to profile changes in gene expression following acute ethanol exposure in Drosophila. 11 13 One of these changes involves the regulation of genes encoding several heat shock proteins (Hsp). Interestingly, one of the most strongly regulated Hsp genes in a recent study of acute ethanol regulation of gene expression in Drosophila was hsp26, 13 a member of the small heat shock proteins (sHsp) that function as molecular chaperones responsible for the prevention of non-native protein aggregation during protein folding/refolding, and facilitation of proteolysis of damaged proteins. 14
Hsp26 is a member of a conserved protein family containing 2 domains: the highly conserved C-terminal region and an N-terminal domain which exhibits sequence divergence ranging from 12 to 40 kDa in different organisms. 15 Unlike the higher molecular weight Hsps, the binding and release of substrates in Hsp26 does not require either ATP binding or ATP hydrolysis. 16 Hsp26 has been shown to be a temperature regulated chaperone in yeast 15 and it is involved in aging and oxidative stress in Drosophila.11,18 Drosophila hsp26 has been shown to be expressed in the neurocytes of the brain and the thoracic ganglion. 19 Hsp26 is transcribed during certain developmental stages in the absence of heat shock 20 and is expressed with a robust circadian rhythm in the adult Drosophila head, 21 indicating a role in sleep homeostasis.
To determine whether the hsp26 gene mediates some aspects of the behavioural adaptations underlying the ethanol response in Drosophila, flies carrying mutations in this gene were tested for ethanol sensitivity and tolerance. Here, we show that whereas flies carrying P-element mutations near the hsp26 gene showed normal ethanol sensitivity, they displayed virtually abolished tolerance to ethanol sedation. The ethanol tolerance defect was also seen when hsp26 gene expression was silenced in glia and the nervous system. Pan-neuronal expression of wild-type hsp26 was also shown to rescue this ethanol tolerance phenotype. Thus, this study has indicated a new role for this member of the small heat shock protein family as an important element in mediating ethanol tolerance.
Materials and Methods
Fly stocks
The wild type strain used in this study was w+; Iso2C; Iso 3I isogenised on the second and third chromosomes and reported to behave similarly to the commonly used Canton-S stock in a range of behavioural tests,
22
and kindly provided by Cahir O’ Kane (University of Cambridge, Cambridge, UK). Mutant strains and P[GAL4] lines used in this study were hsp26EY10556, hsp26KG02086 repo-GAL4 (7415) (chromosome III),
23
elav-GAL4 (III) (8760) and elav-GAL4 (11) (8765) from the Bloomington Drosophila stock centre at Indiana University (Bloomington, IN). UAS-hsp26 (II)
17
flies were obtained from Horng-Dar Wang (National Tsing Hua University, Taiwan) and Tbhnm18 flies from Maria Monastirioti (Institute of Molecular Biology and Biotechnology (IMBB), Greece). The UAS-hsp26RNAi was line v6983 (III) from the Vienna Drosophila RNAi centre. The P[GAL4 lines] were verified for expression in the glia or neurons of the adult Drosophila brain by confocal imaging of the adult brain of flies carrying repo-GAL4 and UAS-GFP or elav-GAL4 and UAS-GFP (Supplementary Fig. 1 and data not shown). Flies were raised on standard maize meal food at 18°C.
Ethanol tolerance measured in the sedation assay. The sensitivity to ethanol vapour of a population of 20 flies was measured in the sedation assay, a simple but highly efficient technique that measures the duration of fly activity after exposure to a sedative dose of ethanol. At 50% standard ethanol concentration, wild-type flies become sedated over a period of about an hour (A) with a mean sedation time (MST) of 53.3 ± 0.3 min (B). When these flies were reintroduced into the vial 4 h after the first exposure, their sedation profile shifted to the right (
Sedation assay
The sedation assay previously described by Wen et al 24 was modified and used to measure ethanol sensitivity in Drosophila. For this assay, 20 active and well fed males (or females as otherwise indicated) were used for each trial. These flies were selected under CO2 anaesthesia and allowed to recover for 24 h before use. 1 ml ethanol solution at 50% concentration was added to a piece of folded Kimwipe tissue (11.4 × 21.5 cm) with edges sealed by transparent tape and laid at the bottom of a 180 ml plastic fly bottle. Flies were then transferred immediately into the bottle and the bottle sealed with a paper lid and parafilm. The active flies remained at the top of the bottle and the sedated flies that dropped to the bottom were counted at 6-min intervals. Counting started immediately flies were introduced into the bottles. The percentage active flies for each time interval of 6 minutes was calculated as the number of flies active at a given time divided by the total number of flies and multiplied by 100 as given by this formula: % Active Flies = xt/N × 100 where xt is the number of active flies at a given sedation time, t and N the total number of sample flies. The Mean Sedation Time (MST) used as a measure of the resistance to the sedative effects of ethanol was calculated as the sum of the number of flies sedated at every 6 minutes multiplied by the time of sedation in minute and divided by the total number of flies sedated as given by this equation:
MST = Σxt × t/N where xt is the number of flies sedated at a given sedation time t and N the total number of flies sedated.
Tolerance assay
Sedation assays measuring rapid tolerance were performed essentially as in the sedation assay for ethanol sensitivity but after initial exposure (MST1), flies were collected in vials and allowed to recover in a humidified room at 18°C on fresh food. They were then exposed to ethanol for a second time. The second exposure (MST2) was initiated exactly 4 h after the start of the first exposure. Tolerance development (ie, percentage increase in tolerance) was calculated relative to the MST of flies following their first and second exposure in the sedation paradigm, using the formula (MST2 - MST1/MST1 × 100).
Heat-shock-ethanol cross-tolerance assay
Flies were incubated at 38°C for 3 min in a vial containing fresh food that had been pre-incubated at 38°C for 18 hrs. After a recovery period of 4 h at 18°C, the flies were exposed to ethanol in the sedation paradigm (MSThs+). Tolerance was calculated with respect to flies that were not heat-treated (MSThs-), using the formula (MSThs+ - MSThs-/MSThs- × 100).
Rescue experiments
P[UAS-hsp26] and P[GAL4-elav]/Cyo transgenic lines were crossed into the hsp26EY10556 mutant background to generate P[UAS-hsp26]/P[UAS-hsp26];hsp26EY10556/hsp26EY10556 and P[GAL4-elav]/ Cyo; hsp26EY10556/hsp26EY10556 flies. For rescue experiments, male flies were Fl progeny of a cross between hsp26EY10556 flies carrying a P[GAL4-elav] driver and UAS-hsp26;hsp26EY10556 flies.
Ethanol absorption and metabolism
Ethanol concentrations were determined from whole fly homogenates of 2 flies per sample using an Analox Instruments (London, UK) ethanol analyser (Alcometer). To study ethanol absorption, flies were exposed to 100% ethanol vapour for 12 min in a recovery assay as previously described 24 and culled at 0, 1, 2, 3 or 4 h recovery time after exposure. Immediately after each recovery period flies were frozen in dry ice and homogenised in 20 μl PBS buffer, pH 7.4. The homogenate was then centrifuged at 12,000 rpm at 4°C for 10 min, and the supernatant was collected. The ethanol concentration in the supernatant was measured using the Alcometer.
Statistical analysis
Statistical significance was assessed by either Student's paired t-tests assuming equal variance or one-way analysis of variance (ANOVA) with Newman-Keuls post-hoc tests.
Results
Rapid tolerance can be measured in the sedation assay
It has previously been reported that adult Drosophila develops tolerance after a single ethanol exposure. 5 Flies upon exposure to ethanol vapour become hyperactive, lose postural control and eventually became sedated. 25 The sensitivity of a population of flies to ethanol can be measured using the sedation assay, a paradigm that quantifies ethanol-induced sedative effects. 24 Briefly, flies are introduced into a vial with a swab soaked with ethanol of a particular concentration. As flies became intoxicated with ethanol, they display hyperactivity before falling to the bottom of the vial and later become sedated. At 50% ethanol concentration (the standard ethanol concentration used in this study) wild-type flies reproducibly become sedated with a mean sedation time of 53.3 ± 0.3 min (Fig. 1B).
To determine whether rapid tolerance could be measured in the sedation assay, flies initially tested in the sedation assay as previously described, were collected in vials containing fresh food and allowed to recover in a humidified room at 18°C before re-testing in the sedation paradigm. The second exposure in the sedation assay (MST2) was initiated exactly 4 h after the start of the first exposure. The 4 h interval was chosen not only to ensure that flies had fully recovered from the sedative effect of ethanol but also to ensure they had completely metabolised all ethanol absorbed and had had time to rehydrate and feed. 5 Tolerance was calculated based on the percentage difference in MST between these two exposures (Figs. 1A, B). It was found that wild-type flies were more resistant to the second exposure to ethanol displaying a MST of 71 ± 4 min, an increase of nearly 18 min from the MST of their first exposure. In this case, tolerance is defined as the relative increase in MST between the first and the second exposure, which for wild-type flies correspond to 33% under the standard experimental conditions.
A previous study using the inebriometer 5 has shown that flies develop rapid tolerance with kinetics of dissipation that was biphasic. To determine the kinetics of tolerance decay using the sedation assay protocol, flies were exposed in the sedation vial twice at a various time intervals and their tolerance acquisition quantified at the appropriate time (Fig. 1C). Consistent with a previous characterisation of rapid tolerance using the inebriometer, it was observed that the kinetics of tolerance dissipation was biphasic, suggesting the involvement of two different processes or mechanisms. Maximal tolerance, a 48.8% ± 1.9% increase in MST, was achieved within a 2 h interval. Tolerance then decreased quickly, reaching ∼12% after 8 h. It then decayed more slowly and was still detecble 24 h after the first exposure, but had disappeared by 36 h (Fig. 1C). Thus, rapid tolerance measured with the sedation assay is very similarly to that previously measured with the inebriometer, an assay that measures the fly's postural control on exposure an intoxicating dose of ethanol.
To validate this present assay Tbhnm18 mutant flies defective for the octopamine biosynthetic enzyme Tbh and previously reported to display impaired rapid tolerance development using the inebriometer assay 5 were tested for rapid ethanol tolerance in the tolerance paradigm. Flies were pre-tested in the sedation assay and were then re-assayed in the same paradigm after 4 h and rapid tolerance was quantified (Fig. 1D). As previously described, the Tbhnm18 mutant showed a marked decrease in ethanol tolerance compared to the wild type control (Fig. 1E), thus validating the tolerance protocol used in this study. This indicates that rapid tolerance, as measured by the sedation and inebriometer assays develops, in part, a common mechanism.
Mutations affecting the hsp26 gene abolish ethanol tolerance
A recent microarray study has shown that the hsp26 gene was strongly upregulated from 0 to 4 h following acute ethanol exposure in Drosophila. 13 Over this time period, the levels of many genes increase and then fall to base-line levels suggesting that they may mediate acute effects of ethanol. Given that the expression level of hsp26 differed from the base line at the end of the 4 h time frame, we wondered if this gene was implicated in ethanol tolerance in Drosophila.
To ascertain a possible role for the hsp26 gene in ethanol tolerance, two mutant alleles carrying different P-element insertions in the same position near the hsp26 gene were tested in the tolerance paradigm. Whereas the ethanol sensitivities of these alleles were normal in the sedation assay, these flies displayed virtually abolished tolerance when tested 4 h after the initial ethanol exposure (Figs. 2A, B) The behaviour of these alleles cannot be explained by an alteration in the kinetics of tolerance acquisition as a similar marked defect in tolerance was observed when these two alleles were exposed to ethanol using protocols earlier shown to produce maximal tolerance in the sedation assay (Figs. 2C, D). Both male and female hsp26EY10556 flies behaved in the same manner. Male flies heterozygous for the hsp26EY10556 mutant were also tested for development of rapid tolerance. The hsp26EY10556 heterozygous males displayed ethanol tolerance that is 75% of wild-type levels suggesting that tolerance is sensitive to the dosage of hsp26. Thus, the two independently isolated hsp26 alleles in their homozygous state exhibited very similar ethanol related behaviours: both showed normal sensitivity and virtually abolished ethanol tolerance development. This indicates the specificity of this behaviour to the hsp26 gene and also rules out any effects due to genetic background at least in the homozygous state.
Tolerance is almost completely abolished in hsp26 alleles. 
Exposure to an acute dose of ethanol induces a stress response.
26
Previous work
6
has indicated that the stress pathway(s) induced by ethanol are linked to, or overlap with, stress pathways induced by other stressors. Hsp26 encodes a cytoplasmic protein involved in the response to several forms of stress, including heat shock.
27
To assess whether prior heat shock of flies might induce ethanol tolerance in the sedation assay, and determine the effect of a heat pulse on the tolerance defect seen in the hsp26 alleles, both the wild-type control and hsp26 flies were tested for ethanol tolerance using the heat shock protocol (see Methods). Heat exposure of wild-type flies (38°C for 5 min) led to a 42% increase in MST when measured in the sedation assay 4 h later. However, the two hsp26 alleles displayed a 10% and 9% increase in MST when treated with the same protocol. Thus, two hsp26 genotypes displayed a substantially reduced level of tolerance compared with the control flies (P < 0.001, Fig. 3). The fact that hsp26 alleles are deficient in the acquisition of tolerance in both protocols indicates that the tolerance produced by ethanol and heat shares some common mechanism(s). hsp26 flies have some capacity for developing tolerance after heat shock, suggesting that other pathways are also involved.
Heat-ethanol cross-tolerance of wild-type and hsp26 flies. 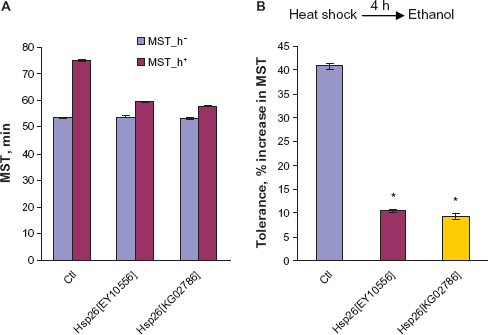
Ethanol absorption and metabolism of hsp26EY10556
One possible explanation for the altered ethanol phenotypes observed for the hsp26 mutants tested in this study is that the modulation of expression of the hsp26 gene is an adaptive response of fly nervous system to compensate for at least some of the sedating effects of ethanol, and this change in expression is at least partially responsible for the altered rapid tolerance to ethanol. However, it is also possible that these mutants exhibit altered ethanol phenotypes as a result of differences in their ethanol absorption and/metabolism when compared to a wild-type strain.
To determine whether there was altered ethanol metabolism in flies, the ethanol concentration in fly extracts prepared from hsp26EY10556 and wild-type flies exposed to constant ethanol vapour (100%) for 12 min and allowed to recover for 0, 1, 2, 3, or 4 h was measured. The ethanol concentration in extracts of hsp26ET10556 flies was indistinguishable from that of the wild-type control flies at all time points (Fig. 4). As shown in the Fig. 4, the results reveal a very similar rate of ethanol clearance for the two genotypes. Therefore, the altered ethanol phenotypes of these mutants were more likely due to pharmacodynamic changes, and not pharmacokinetic changes. The results also give an ethanol content of about 44 mM in wild-type flies immediately after sedation. This corresponds to about 0.20% in human blood alcohol concentration (BAC) which causes severe loss of the response to sensory stimuli and loss of consciousness.
28
Ethanol concentrations in flies after ethanol exposure. No significant differences were seen between the hsp26EY10556 mutant and the wild-type control (Student's t-test; n = 3).
Neuronal expression of Hsp26 on ethanol sedation
The results described above implicate the hsp26 gene in the regulation of ethanol-induced behaviour: hsp26 mutations affect tolerance but not sensitivity of flies to ethanol in the sedation assay. To determine whether hsp26 mediates this behaviour by acting within the nervous system, we used the GAL4/UAS gene expression system 29 to target the expression of the hsp26 gene to the nervous system.
First, to determine whether silencing of Hsp26 expression in neurons can mimic the normal ethanol sensitivity phenotype observed in the hsp26 mutant flies, we used UAS-hsp26RNAi to knockdown the expression of the hsp26 gene in the nervous system of wild-type files using elav-GAL4 driver, which drives expression in all Drosophila neurons. Flies carrying the elav-GAL4 driver and a P[UAS-hsp26RNAi] were generated and tested for ethanol sensitivity in the sedation assay. As with the results from hsp26 mutants, pan-neuronal silencing of the hsp26 gene in all neurons did not result in any significant alterations in ethanol sensitivity (Fig. 5A blue bar). We also determined whether an increase in the endogenous levels of Hsp26 expression in all neurons alters the behavioural response to ethanol. Thus, flies carrying one wild type copy each of the UAS-hsp26 transgene and elav-GAL4 were tested for ethanol sensitivity in the sedation assay. Overexpression of Hsp26 in the nervous systems led to resistance to the sedating effects of ethanol (Fig. 5C blue bar). The overexpression results correlate with the microarray data presented by Kong and colleagues.
13
When flies were exposed to ethanol, transcription of hsp26 rose. This response may reflect a protective function of the gene and probably buffers the cell from further insults, hence, causing the reduced ethanol sensitivity of flies overexpressing this gene in the nervous system.
Hsp26RNAi expression in the nervous system reduced tolerance. Upper and lower left panel (
In addition, we also determined whether silencing of Hsp26 expression in the nervous system can mimic the ethanol tolerance phenotypes observed in the hsp26 mutant flies using the GAL4/UAS gene expression system as described above. As expected, pan-neuronal silencing of the hsp26 gene results in reduced ethanol tolerance from 33.0 ± 1.3 (UAS-hsp26RNAi) or 34.6 ± 0.8 (elav-GAL4) to 18.7 ± 1.3 (UAS-hsp26RNAi + elav-GAL4) (Fig. 5B). Neither the elav-GAL4 driver nor the P [UAS-hsp26RNAl] alone displayed reduced ethanol tolerance, indicating that the tolerance defect observed required the presence of both of these drivers. However, mutations in hsp26 were previously shown to completely abolish the development of tolerance in flies. One interpretation for these results is that the silencing activity of hsp26RNAi did not completely eliminate hsp26 gene activity and the residual hsp26 gene expression resulted in the small level of tolerance seen, suggesting that the acquisition of rapid tolerance is sensitive to levels of Hsp26. Consistent with this is the observation that one wild-type copy of hsp26 gene is sufficient to increase tolerance to 75% of wild-type levels (Fig. 2B).
Although reduced levels of Hsp26 reduce rapid tolerance, increased levels of this protein have no effect when elav-GAL4 was used to drive hsp26 in neurons in a wild-type background (Fig. 5D). The overexpression of hsp26 in the nervous system did not alter rapid ethanol tolerance when compared to the elav-GAL4 driver or the P[UAS-hsp26] transgene alone (Fig. 5D). This suggests that unlike sensitivity to sedation, ethanol tolerance was not affected by raised Hsp26 levels in the nervous system.
Pan-neuronal expression rescues the hsp26EY10556 ethanol tolerance phenotype
To further confirm the role of the hsp26 gene in the development of ethanol tolerance, the ability of a hsp26 transgene to rescue the tolerance defect seen in hsp26EY10556 was examined. elav-GAL4/UAS-hsp26; hsp26EY10556 flies were tested for ethanol tolerance. Pan neuronal expression of wild-type hsp26 restored the ethanol tolerance of hsp26EY10556 flies to wild-type control levels (Fig. 6A, B). The MST of the wild-type flies was not significantly different from the MST of the elav-GAL4; hsp26EY1055/UAS-hsp26; hsp26EY10556 flies. Neither the elav-GAL4 driver nor the P[UAS-hsp26] transgene alone altered the ethanol tolerance of the hsp26EY10556 mutant flies substantially (Fig. 6B). Taken together, both of these experiments show that hsp26 activity is required for rapid ethanol tolerance and that hsp26 regulates rapid ethanol tolerance by acting within the nervous system.
The hsp26EY10556 tolerance defect can be rescued by expression of a UAS-hsp26 transgene in the nervous system under the control of the elav-GAL4 driver. 
Although expression oîhsp26 in neurons fully rescues the tolerance defect, there is a possibility that glia are also involved in this process. Silencing the expression of hsp26 in glia, by driving the expression of hsp26RNAi with repo-GA14, a glia-specific construct, led to a reduced tolerance phenotype (Figs. 7A, B). This raises the possibility that glia partially mediate ethanol tolerance. In contrast, to determine whether increased expression of endogenous Hsp26 in the glial cells also alters ethanol tolerance behaviour, repo-GAL4/UAS-hsp26 flies, in a wild-type background were also tested for rapid ethanol tolerance. The overexpression of hsp26 in the glial cells did not alter rapid ethanol tolerance (Figs. 7C, D) when compared to repo-GAL4 driver or the P[UAS-hsp26] transgene alone suggesting that ethanol tolerance was not affected by raised Hsp26 levels in the glia cells.
Effect of glia on hsp26 ethanol tolerance. Upper and lower left panel (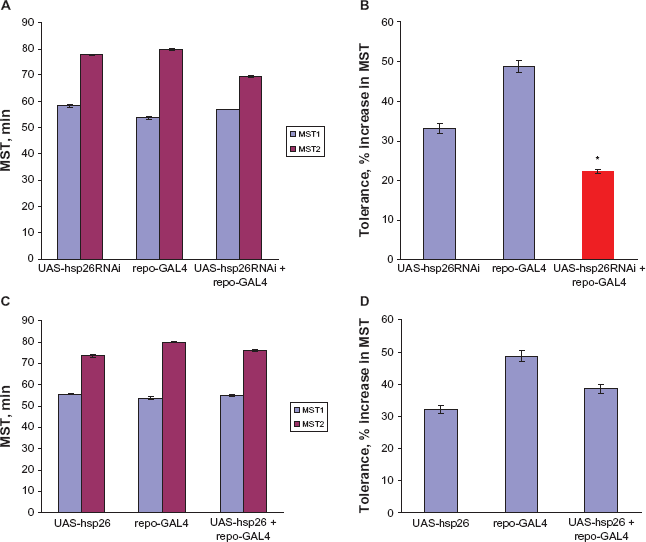
Discussion
The Drosophila hsp26 gene is a member of small heat shock protein family that is induced by a variety of stressors including aging and oxidative stress.17,18 Here, we show that Hsp26 is also required for the development of tolerance to ethanol, although mutations in the gene do not affect the response of Drosophila to the sedating effects of ethanol. Interestingly, the ethanol tolerance phenotype of hsp26 is sensitive to the level of the gene product; however, there is a threshold above which increased levels of Hsp26 have no further effect on tolerance. Hsp26 is required in neurons for normal ethanol-induced behaviour.
Sedation assay measures rapid tolerance in Drosophila
We developed an assay to measure tolerance to the sedative effects of ethanol in Drosophila. Using this assay, rapid tolerance, which is induced by exposure of flies to a single moderately high ethanol dose, was quantified as a change in sensitivity observed upon administration of a second dose of ethanol after the first dose was completely metabolised. 30 The quantitative aspects of ethanol tolerance in flies, such as the extent of maximal tolerance and the kinetics of its decay (dissipation) were similar to those described using the inebriometer. 5 In addition, it was shown that octopaminergic systems previously implicated in a component of rapid tolerance 5 were also involved in impaired ethanol tolerance in this assay. Taken together, these data demonstrate that the sedation assay is a suitable paradigm in which to measure the tolerance of Drosophila to the sedative effects of ethanol.
Hsp26 ethanol tolerance and the molecular pathways involved
Scholz and colleagues 6 have shown that the development of rapid tolerance in Drosophila is mediated by at least two molecular pathways. One is specific to ethanol tolerance and requires the neuromodulator, octopamine. The other pathway is a cellular stress response that is also induced by heat-shock. Using tolerance to sedation, we have also shown the requirement for the cellular stress pathway in the acquisition of rapid tolerance. Pre-exposure of flies to ethanol can be mimicked by heat-shock, resulting in a 42% increase in MST when exposed to ethanol four hours later, an increase that is reduced to only 10% in hsp26 mutants. Mutations in the gene hangover, a nuclear zinc-finger protein, also affect this pathway but do not impinge on the ethanol-specific pathway. Hsp26, however, appears to be required for both of these pathways as mutations in this gene completely abolish tolerance. Although hsp26 mutations eliminate ethanol-induced tolerance, hsp26 flies do have the ability to develop some heat-shock ethanol cross-tolerance. As noted above, there is a small (10%) amount of residual tolerance in hsp26 flies after heat-shock, but not after prior exposure to ethanol. Interestingly, the maximum amount of tolerance induced after heat-shock is about 8% higher than that induced by ethanol (40.8% cf 33.0%). This suggests that the residual tolerance seen in hsp26 flies is due to yet a third tolerance pathway that can be activated by stressors other than ethanol.
Hsp26 regulates distinct ethanol-induced behaviour in Drosophila
Hsp26 is not required for all ethanol-induced behaviours. Sedation is normal in hsp26 mutants, showing that sedation and rapid tolerance are mediated, at least in part, by distinct molecular pathways. The relationship between sedation and tolerance is probably complex, however, as several genes have been identified that disrupt both sensitivity and tolerance to ethanol. 10 13 To untangle the underlying mechanisms will require further study of the action of individual genes.
Why Hsp26 had distinct roles in sensitivity to ethanol and in the acquisition of tolerance was at first, surprising, given that it is widely expressed in Drosophila. However, the sHsp protein family has expanded and diversified in metazoans and it is possible that this diversification may have been driven by the acquisition of specific functions for sHsps during evolution. 31 In Drosophila, for example, although both Hsp26 and Hsp27 are required for an increase in lifespan and elevated resistance to oxidative stress, only Hsp27 can partially block apoptosis. 18
Hsp26 regulates ethanol tolerance by acting within the nervous system
Our present study has shown that silencing of the hsp26 gene in the neurons reduced tolerance to ethanol while the ethanol tolerance defect seen in hsp26 mutant flies can be rescued by pan-neuronal expression of wild-type hsp26, thus supporting the hypothesis that Drosophila Hsp26 regulates ethanol tolerance through functions in the nervous system. This raises the possibility that Hsp26 may be acting through a mechanism to protect the neural cells from ethanol-induced damage, thus maintaining the integrity of the neurons through the promotion of neural cell stabilisation and survival. Our finding agrees with previous studies that have mapped tolerance to specific brain regions in Drosophila including the ellipsoid body neurons 12 and subset of neurons in the central complex. 5 However, we also observed that a reduction in the level of Hps26 using RNAi in glia lowered tolerance to ethanol. Thus, the observation that wild-type levels of tolerance can be achieved by pan-neuronal expression of Hsp26 indicates that any requirement for the function of this protein in glia can be bypassed.
Hsp26 expression and behavioural response to ethanol
Hsp26 is induced in response to stress in a variety of organisms. As ethanol is a cellular stressor, it is not surprising that hsp26 and other stress-induced proteins are highly upregulated following acute ethanol exposure in Drosophila. 13 This upregulation together with our finding that Hsp26 is critical for at least one ethanol-induced behaviour has important functional consequences for the cell. In order to acquire tolerance, Drosophila must first be challenged by a moderate dose of ethanol. Drosophila cells then undergo a series of biochemical and physiological changes not only in attempt to cope with the ethanol stress but also in enhancing the survival of the stressed cells following exposure. Thus, a hypothesis to explain the role of this protein in tolerance would be that it plays a protective role in the nervous system. In addition, the upregulation of this protein following ethanol sedation may occur to ensure cellular protein homeostasis and cell protection from ethanol insults.
In conclusion, further study of the role of hsp26 in neuronal adaptation to ethanol together with identification of its downstream targets could provide a useful hint for understanding the development of tolerance and dependence in alcoholics. Nevertheless, this study has established that Drosophila hsp26 plays a crucial role in ethanol tolerance.
Disclosure
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers of this paper report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material.
Supplementary Figure

Confocal images of the adult Drosophila brain showing P[GAL4] directed expression of GFP in the brain neurons (elav-GAL4) and the glia cells (repo-GAL4).
Footnotes
Acknowledgments
This work was supported by a fellowship from Tai Solarin University of Education to AAA and a BBSRC research grant to JAD.