
Abstract
Hemoglobin A1c (HbA1c) is an important parameter for evaluating long-term (6–8 weeks) control of blood glucose levels in patients with diabetes mellitus. We report a Japanese diabetic case who harbored abnormal Hb and demonstrated falsely low HbA1c levels in the regular ion exchange HPLC-based assay. His abnormal β-globin chain had an amino acid replacement that corresponded to glycine to serine substitution at amino acid position 136 of the β-globin chain (Hb Perpignan). Accordingly, a heterozygotic point mutation replacing guanine by adenine at nucleotide position 406 (β136G > S) was identified in the β-globin gene of the proband and his son. These results indicate that the patient had Hb Perpignan, and this abnormal Hb interfered with the HPLC-based HbA1c assay commonly used in the clinic. The cases presented are the first report of patients in the Japanese population harboring Hb Perpignan (β136G > S).
Introduction
Hemoglobin (Hb) A is a convenient and useful indicator for the long-term (6–8 weeks) control of blood glucose levels in patients with diabetes mellitus. 1 Guidelines for diabetic care are determined by the Japan Diabetes Society which describes values less than 6.5% as a fair control. 2 It is however known that abnormal HbAs sometimes interfere with precise determination of HbA1c levels in these patients. 3 HbA, virtually the sole hemoglobin produced throughout life after birth of normal subjects, is a tetramer consisting of 2 α-and 2 β-globin chains, while HbF expressed in the fetus consists of γ-globins instead of α-globins. 4 α-globin represented as 2 isoforms has 141 amino acids, while β-globin chains are composed of 146 amino acids. 5 So far, over 890 different mutations found in the α-, β- or γ-globin genes are known to produce abnormal Hbs. 6 Sickle cell anemia and thalassemia, which are caused by mutations in the α- or β-globin chain and develop characteristic hemoglobinopaties including hemolytic anemia, splenomegaly and production of fetal Hb, are the most popular disorders associated with abnormal Hbs. In addition, 2 mutations in the β-globin gene cause embolic events through polymerization of mutated HbA, 91 increased affinity of the HbA to oxygen, and 134 destabilized this protein complex 7 . Further, 9 mutations produced metho-Hbs and developed clinical symptoms characteristic to methohemoglobinemia. 7 The rest of the reported mutations appear to be asymptomatic, 7 but their encoding HbAs sometimes cause false measurement of glycosylated HbAs, such as HbA1 and HbA1c, in specific laboratory assays, thus they hamper regular clinical care of diabetic patients. 3
We identified two cases (father and son) in a Japanese family who carried a novel heterozygotic point mutation replacing guanine by adenine at nucleotide position 406 in the β-globin gene that generated abnormal HbA harboring glycine to serine replacement at amino acid position 136 (Hb Perpignan). The proband had type 2 diabetes mellitus and demonstrated unreasonably low HbA1c levels for his blood glucose control in the ion exchange HPLC-based method. We report here detailed characteristic of the patient's clinical profile and his abnormal Hb.
Case Report
Proband, a 78-year old male, had been treated for his type 2 diabetes mellitus using oral hypoglycemic agents for over 40 years. He had peripheral neuropathy without any other complications associated with diabetic micro- and macro-angiopathy. During follow-up, his HbA1C levels determined by a regular laboratory assay fluctuated around 6.0%∼6.8% (normal range (nr): 4.3%–5.8%), while plasma glucose levels at the postprandial period were always above 200 mg/dL (nr: <200 mg/dl), indicating a discrepancy between HbA1c values and his blood glucose control. At presentation in our hospital, the percentage of his HbA1c measured with the ion exchange HPLC, was 5.9% (nr: 4.3%-5.8%), while levels of his postprandial plasma glucose, 1,5AG [1,5-Anhydro-D-glucitol] and glycoalbumin were 229 mg/dL (nr: <200 mg/dl), 12.2μg/mL (nr: <14.0μg/mL) and 28.0% (nr: 11%–16%) respectively, thus his HbA1c value was incorrectly low compared to his overall blood glucose control estimated from the latter indicators. In the same sampling, his red blood cell count (RBC) was 458 x 104/μL (nr: 403 x 104–596 x 104/μL), while his Hb concentration and hematocrit (Ht) were 13.1 g/dL (nr: 13.3–17.5 g/dl) and 39.9% (nr: 38.2%–50.9%), respectively. The mean corpuscular volume (MCV) was 87.1 fL (nr: 81.8–100.0fL), the mean corpuscular hemoglobin (MCH) was 28.6 pg (nr: 28.4–34.7 pg), and the mean corpuscular hemoglobin concentration (MCHC) was 32.8% (nr: 32.7%–36.3%).
Materials and Methods
Methods for determining patient's HbA1c levels
Levels of HbA1c were determined by using the HA-8170 apparatus (Arkray, Kyoto, Japan), which employs a regular ion exchange HPLC method for quantification of this glycohemoglobin, and in an immunological method using the HbA1c Reagent Kit (Bayer HealthCare LLC, Mishawaka, IN, USA) and the DCA2000+Analyzer (Bayer HealthCare LLC, Mishawaka, IN, USA). Instability of Hb was examined in a precipitation method using 17% isopropanol as previously reported. 8 All clinical studies were conducted in accordance with the principles of the Declaration of Helsinki. The “Ethical Guidelines for Clinical Studies” issued by the Ministry of Health, Labor and Welfare of Japan in 2003, with local Ethics Committee approval being obtained at Yokohama Rosai Hospital. All patients were fully informed and written consent was obtained.
Evaluation of the molecular weight and amino acid sequence of patient's β-globin chain
Analysis on the molecular weight of the patient's β-globin peptide was performed by using the TSQ-7000 triple-stage quadruple mass spectrometer equipped with an electrospray ion source (Finnigan MAT, San Jose, CA, USA), as previously reported. 9 For examining the amino acid sequence of patient's β-globin chain, the globin fraction was purified with the acetone precipitation method, 10 and was subsequently cleaved with trypsin. The on-line HPLC electrospray ionization mass spectrometry (ESI/MS) and the tandem mass spectrometry (MS/MS) were then performed to isolate abnormal peptides, and mutated amino acids were identified by using the LCQDECA ion trap mass spectrometer equipped with an electrospray ion source (Thermo Quest, San Jose, CA, USA), according to the previous report. 11 The hemoglobin β-chain gene (HBB) of DNA extracted from leukocytes was amplified using the polymerase chain reaction (PCR) method. 12 The direct sequencing method was employed to determine the DNA sequence on an Applied BioSystems 3130 Genetic Analyzer (Applied BioSystems LLC, Foster City, CA, USA) using a Big Dye Terminator v1.1 Cycle Sequencing Kit (Applied BioSystems LLC) according to the manufacturer's instructions.
Results
Because the patient's HbA1c level was low compared to his blood glucose control, we measured HbA1c using the immunological method in the same sample processed with the ion exchange HPLC method. We found that the HbA1c level in this method was 7.5%, thus there was a clear discrepancy between the values measured with these two procedures. We next examined instability of the patient's Hb as it appeared to be abnormal, but the result was negative (data not shown).
We then examined an elution pattern of the patient's Hb in the ion exchange HPLC (Fig. 1). We found that there was an abnormal HbA1c peak with a notch immediately before the large HbA0 peak (Fig. 1), indicating that there are two types of HbA1c fractions circulating in patient's blood. Consistent with this finding, the transformed ESI/MS spectrum of patient's undigested β-globin revealed that the patient had two β-globin proteins: one was the normal β-globin peptide demonstrating the normal molecular weight [15867.60 Da], while the other was an abnormal β-chain with 29.5 Da larger than the former (Fig. 2).
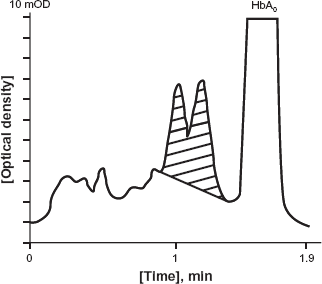
Ion exchange HPLC elution profile of the patient's HbA1c. A split abnormal A1c peak (shaded area) was observed immediately before HbA0 fraction.
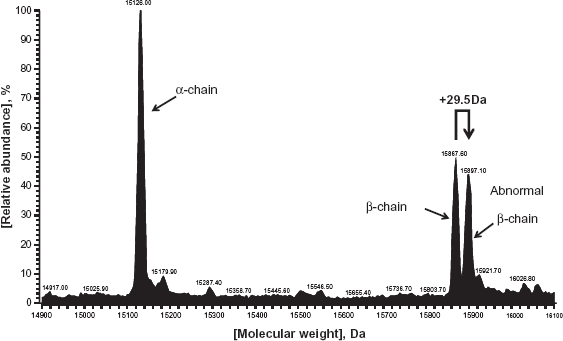
Transformed ESI mass spectrum of the patient's whole HbA. An abnormal β-chain peptide with 29.5 Da larger molecular weight than the normal β-chain was detected.
In the analysis for the tryptic globin digests using the on-line HPLC/ESI/MS, the patient's sample demonstrated two βT-14 elution peaks, a normal peak observed at 18.68 min and an abnormal peak at 19.59 min, together with a normal βT-11 peptide peak recorded at 19.7 min (data not shown). The ESI spectrum of the normal and abnormal βT-14 peaks revealed that the latter peptide had the molecular weight approximately 30 Da larger than the normal peptide, as elution peak of an abnormal βT-14 peptide was recorded at 1,180.3 (m/z) while that of the normal βT-14 was seen at 1,150.4 (Fig. 3). We further performed the MS/MS analysis on the abnormal βT-14 peptide. We found that glycine corresponding to the amino acid located in position 136 of the β-globin peptide was substituted by serine. Figure 4 shows the elution pattern of MS/MS for analyzing β-globin peptide.
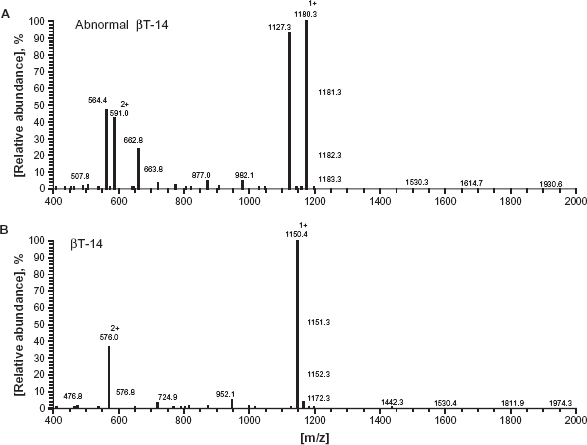
Spectra of the on-line HPLC/ESI/MS obtained with the triptic digests of the patient's β-globin peptide. A) Singly and doubly charged ions of abnormal βT-14 (m/z 1180.3 and m/z 591.0) and normal βT-11 (m/z 1127.3 and m/z 564.4) were recorded at elution time 19.7 min. B) Singly and doubly charged ions of normal βT-14 (m/z 1150.4 and m/z 576.0) were recorded at elution time 18.8 min.
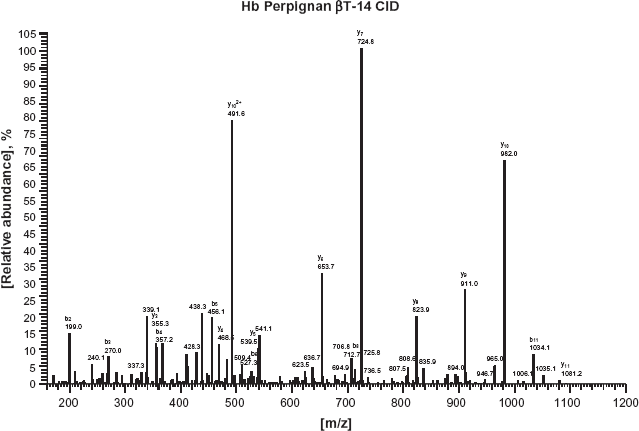
MS/MS pattern of analyzing β-globin peptide. Glycine corresponding to the amino acid located in position 136 of the β-globin peptide was substituted by serine.
We finally performed a DNA sequence of the patient's β-globin gene (HBB) (Fig. 5). Sequence signal peaks of guanine and adenine were both recorded at nucleotide 404, indicating that the patient HBB has a heterozygotic point mutation at this nucleotide position. Thus, the mutated allele produces an abnormal β-globin chain harboring glycine (GGT) to serine (AGT) substitution at amino acid position 136 (β136G > S), while the normal allele encodes the regular β-globin, consistent with our results obtained in the HPLC-based analysis indicating that the patient possessed two β-globin peptides of 30 Da difference. We also identified the same heterozygotic point mutation in the HBB gene of patient's son (data not shown).
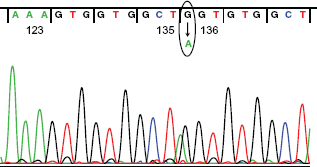
Identification of heterozygotic point mutation replacing guanine by adenine at nucleotide position 406 in the patient's HBB gene. Signals of guanine and adenine were identified at nucleotide position 406 of the patient's HBB gene (circled), which develop abnormal β-globin chain harboring glycine (
Discussion
We identified a 78-year old male case harboring a heterozygotic point mutation (β136G > S) in the HBB gene. The β136G > S mutation of the HBB gene had previously been reported in a Caucasian French man, and the Hb consisting the encoded abnormal β-globin is known as Hb Perpignan. The current report is the second report describing the cases harboring the β136G > S mutation (Hb Perpignan) internationally, while they are the first subjects in the Japanese population. Affinity of this abnormal Hb to oxygen was reported to be normal, while we demonstrated that this abnormal Hb did not show protein instability, thus these subjects do not appear to develop obvious symptoms or signs required for medical attention.
It is well known that carbamyl Hb, acethyl Hb, acetaldehyde Hb, HbF and abnormal Hb, which shows some replacement of amino acids, are demonstrating several abnormal peaks in the elution pattern of HPLC. Thus, it is impossible to determine the exact value of HbA1c when analyzed by the usual HPLC system. However, the turbidimetrical immunoassay system uses a HbA1c peculiar antibody and the antibody can recognize 3 or 4 amino acids from N-terminal of the β chain. Thus, it is suggested that the turbidimetrical immunoassay system can accurately detect the real value of HbA1c despite the presence of abnormal Hb which shows no abnormalities in the N-terminal of the β chain.
Incidence of abnormal Hbs is estimated to be 1/2,000 to 1/3,000 in the Japanese population. 13 Several diabetic patients were reported to demonstrate unreasonably low HbA1c values compared to their plasma glucose control,14–22 while others showed incorrectly high HbA1c levels.23–28 Some rare Hb variants, such as Hb D, Hamaden, Himeji, Hoshida, North Manchester, Riyadh and Sagami, cause such low HbA1c levels in the cation exchange HPLC-based assay, 6 whereas other uncommon Hb variants, including Hb J, Graz, Niigata, Okayama, Old Dominion, Raleigh, Sherwood Forest, and South Florida, originate spuriously high HbA1c in this assay method. 6 Based on our current findings and evidence shown in these previous reports, special attention is needed to the presence of abnormal Hbs, once glycosylated HbA values consistently mismatch to those of other parameters for glucose control in diabetic patients.
Disclosure
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers of this paper report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material. Written consent was obtained from the patient or relative for publication of this study.
Footnotes
Acknowledgements
This work was partly supported by the Grant for Research on Intractable Diseases provided from the Japanese Ministry of Health, Labor and Welfare. We thank Dr. T. Kino (National Institutes of Health, Bethesda, MD) for critically reading and for providing his valuable suggestions.