
Abstract
Excess L-tryptophan (L-Trp) in the diet decreases fetal body weight. However, the relationship between L-Trp concentration and its effects on maternal, placental, and fetal growth are not well-understood. We investigated the effects of excess L-Trp intake on maternal, placental, and fetal growth. Female mice were fed a 20% casein diet (control diet) or control diet plus 2% or 5% L-Trp during gestation. Pup weights did not differ between the control (L-Trp intake: 0.04 g/kg body weight (BW)/day) and 2% L-Trp groups (L-Trp intake: 3.3 g/kg BW/day), but were significantly lower in the 5% L-Trp group (L-Trp intake: 7.0 g/kg BW/day) than in the control and 2% L-Trp groups. These results show that less than 3.3 g/kg BW/day L-Trp intake in pregnant mice during gestation does not affect fetal growth or L-Trp homeostasis in the placenta or fetus.
Introduction
L-tryptophan (L-Trp) is an essential amino acid and is an important precursor for vitamin nicotinamide (Nam), and a neuron transmitter, and can be converted into smooth muscle serotonin (5-hydroxytryptamine, 5-HT).1–4 L-Trp supplements are readily available from local supermarkets. 4 Thus, understanding the effects of excess L-Trp on human health is important.
The effects of excess L-Trp have been reported in rodents.5–9 Feeding of a high L-Trp diet to experimental animals induced fatty liver 6 and lung inflammation.7,8 Generally, toxicity due to the excess intake of a particular nutrient is likely to appear during pregnancy.10,11 Good nutrition is particularly important for pregnant animals as well as fetuses to maintain overall health during pregnancy. The requirement for nutrients increases during pregnancy compared with the normal state. Therefore, information regarding safe levels of particular nutrients as well as the amounts that increase the risk of adverse effects is required. In pregnant rodents, fetal body weight was shown to be lower in animals fed a high L-Trp diet compared with rodents fed an normal diet.12,13 However, the relationships between pregnancy and excess L-Trp are not well-documented.12–15
It has been reported that L-Trp levels in maternal serum decrease during normal pregnancy.16,17 However, there have been no studies examining L-Trp metabolism on maternal, placental, and fetal growth in fed a high L-Trp diet. We investigated the relationships between L-Trp intake and L-Trp concentration in maternal, placental, and fetal animals and the subsequent effect on growth. In addition, excess L-Trp in the placenta may restrict fetal growth. Therefore, we investigated the effects of a high L-Trp diet during gestation on placental development and fetal growth. We also measured mRNA expression of enzymes involved in the L-Trp degradation pathway in the placenta.
Materials and Methods
Ethical considerations
The care and treatment of the experimental animals conformed to the University of Shiga Prefecture guidelines for the ethical treatment of laboratory animals.
Animals and diets
Female imprinting control region (ICR) mice (6-week-old) were obtained from Charles River (Tokyo, Japan). They were immediately housed in a large plastic cage (225 × 338 × 140 mm, Clea Japan, Tokyo, Japan). Initially, all mice were fed ad libitum a 20% casein diet (Table 1) for 3 days. They were then divided into three groups. Group 1 was fed ad libitum the 20% casein diet (control), group 2 was fed ad libitum the 20% casein diet plus 2% L-Trp (2% L-Trp group), and group 3 was fed ad libitum the 20% casein diet plus 5% L-Trp (5% L-Trp group) (Table 1) throughout the experiment except for mating with a male mouse. All animals were allowed free access to water. The room in which the mice were kept was maintained at a temperature of approximately 22 °C with 60% humidity and a 12-h light/12-h dark cycle (light on at 6:00 h and off at 18:00 h).
Composition of diets.

Reeves RG: Components of the AIN-93 diets as improvements in the AIN-76A diet. J Nutr. 2008;127:838S–41S.
Mating
Female mice fed more than 10-days feeding of the control, 2% L-Trp, and 5% L-Trp diets were mated with a male adult ICR mouse. Each female mouse in estrus was moved to in a small plastic cage (155 × 245 × 148 mm, Clea Japan) at 16:00 h, and then mated with one male adult ICR mouse fed a commercial non-purified diet (MF; Oriental Yeast Co., Ltd., Tokyo, Japan). The next day, we checked for a vaginal plug at 09:00 h. Female mice were placed back in the large plastic cage and fed the original chemically defined purified diet (control, 2% L-Trp, or 5% L-Trp diet) during gestation. The time was designated as embryonic day (E) 0.5. The normal gestational period of mice is E19.5. Body weights of the pregnant mice and food intakes were measured throughout gestation at 09:00 h each day.
Experiment 1
L-Trp concentrations of the liver, blood, placenta, and fetus
Pregnant mice fed the 20% casein diet (control), 20% casein diet plus 2% L-Trp, or 20% casein diet plus 5% L-Trp were killed at either E12.5 (control group, n = 8; 2% L-Trp group, n = 5; 5% L-Trp group, n = 10) or E17.5 (control group, n = 5; 2% L-Trp group, n = 4; 5% L-Trp group, n = 6). Blood was acquired by cardiac puncture. The maternal liver, fetuses, and placentas were removed and weighed. Non-proteinaceous L-Trp concentrations, which is the total amount of free form of L-Trp and bound form of L-Trp with albumin and macromolecules in the maternal blood, maternal liver, placenta, and fetus, were measured by high-performance liquid chromatography (HPLC). 18
mRNA expression of L-Trp-degrading enzymes and LAT1 in the placenta by real-time RT-PCR
At E12.5 and E17.5, two or three placentas were selected at random from each dam per group. mRNA expression of L-Trp-degrading enzymes of 17 types (Table 2) and large neutral amino acid transporter 1 (LAT 1) mRNA expression levels were measured in the placenta. Total RNA was extracted using the RNeasy® Mini Kit (cat. no. 74124, Qiagen, Hilden, Germany) according to the manufacturer's instructions. Reverse transcription was performed on 4 μg of RNA with oligo-dT primers using the PrimeScript® II 1st strand cDNA Synthesis Kit (6210A/B, Takara Bio, Japan). The FastStart Unucersal SYBR Green Master (ROX) (Roche, Mannheim, Germany) was used for PCR. The expression of mRNA was quantified using the ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). The reverse transcription (RT)-PCR conditions were 50 °C for 2 min, 95 °C for 10 min, 95 °C for 15 s, and 60 °C for 1 min for 40 cycles. Real-time RT-PCR was carried out with the primers shown in Table 2. All data were normalized to the endogenous reference gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels and shown as the fold-change relative to the control.
Primers used to measure mRNA expression of enzymes involved in L-Trp degradation in the placenta at E12.5 and E17.5.

Placental morphometry
The placenta was collected from three dams from the control and 5% L-Trp groups at E12.5 or E17.5. Placentas were immersed in 4% paraformaldehyde for 24 h, embedded in Tissue-Tek OCT Compound (Sakura Finetek Japan Co., Tokyo, Japan), and stored at –80 °C until sectioning. Sections were cut to 5 μm, mounted on glass slides, stained with hematoxylin-eosin (HE), and prepared for subsequent microscopic mounting. HE staining was performed according to the standard protocol. Digitized images were captured from each placenta section using a Pentax digital camera (Pentax Ricoh Imaging Co., Ltd., Denver, CO, USA) equipped with a microscope (Carl Zeiss Japan Co., Tokyo, Japan) at a final magnification. The placenta was classified into the decidua basalis, spongiotrophoblast, and labyrinth zone. 19 For quantification, digitized images were captured on the same scale using the same imaging system described above. The decidua basalis, spongiotrophoblast, and labyrinth zone thickness at the thickest point in the placenta were measured using ImageJ software (Image Processing and Analysis in Java. http://rsb.info.nih.gov/ij/).
Experiment 2
Death rate of pup
To elucidate whether fetuses born from mothers fed the high L-Trp diet could live after birth, pregnant mice fed the 20% casein diet (control), 20% casein diet plus 2% L-Trp, or 20% casein diet plus 5% L-Trp were killed at E 18.5 (control group: n = 4; 2% L-Trp group: n = 2; 5% L-Trp group: n = 8) and the pups were removed by caesarean delivery. Pup body weights were measured and pup behavior was immediately monitored with a video camera (GZ-HM350, JVC Co., Kanagawa, Japan) for 2 min. We then measured respiration frequencies of the pups using the video data.
Lung morphometry
One hour after caesarean delivery, pups in the control and 5% L-Trp groups were sacrificed and their lungs were collected from three dams per group. Lungs were immersed in 4% paraformaldehyde for 24 h, embedded in Tissue-Tek OCT Compound (Sakura Finetek), and stored at –80 °C until sectioning. Sections were cut to 5 μm, mounted on glass slides, stained with HE, and prepared for subsequent microscopic mounting. HE staining was performed according to the standard protocol. We determined the alveolar area according to the method described by Chen et al. 20 Digitized images were captured from each lung section (ie, 30 images per lungs) using a Pentax digital camera (Pentax Ricoh Imaging) equipped with a microscope (Carl Zeiss Japan) at a final magnification of ×500. Six images were chosen randomly, and alveolar airspace in the image was measured using ImageJ software.
Statistical analysis
Data are expressed as the mean ± SE. Maternal body weight changes were compared by two-way analysis of variance (ANOVA) for each day among control, 2% L-Trp, and 5% L-Trp groups. Placental layer thicknesses were compared by Student's t-test between the control and 5% L-Trp groups. Other data were compared by one-way ANOVA and subsequent Tukey-Kramer multiple-comparison post-hoc tests. Differences were considered significant at P < 0.05. Prism 5.0 (Graph Pad Software, San Diego, CA, USA) was used for all analyses.
Results
L-Trp intake and changing maternal body weight during gestation (experiment 1)
L-Trp intake was 0.04 g/kg BW/day, 3.3 g/kg BW/day, and 7.0 g/kg BW/day in the control, 2% L-Trp, and 5% L-Trp groups, respectively, during gestation. Figure 1 shows the body weights of mice during gestation. These values did not include the aborted pregnant mice. The body weights of pregnant mice were not significantly different among the control, 2% L-Trp, and 5% L-Trp groups through gestation (Fig. 1). There were no abortions in the control group (plugging, n = 13; abortion, n = 0) or the 2% L-Trp group (plugging, n = 9; abortion, n = 0). However, 31.3% of mice aborted in the 5% L-Trp group (plugging, n = 16; abortion, n = 5).

Pregnant mice body weight change during gestation by feeding of a high L-Trp diet (experiment 1).
Reduction of placental and fetal body weights by feeding a high L-Trp diet (experiment 1)
Placental weights and fetal body weights at E12.5 or E17.5 are shown in Table 3. Placental weights and fetal body weights at E12.5 were lower in the 2% L-Trp group than the control group (Table 3). At E17.5, placental weights did not differ between the control and 2% L-Trp groups (Table 3). Fetal body weights were lower in the 2% L-Trp group than the control group at E17.5 (Table 3). Placental weights and fetal body weights were markedly reduced only in the 5% L-Trp group but not in the 2% L-Trp group at E12.5 and E17.5.
Inhibition of placental and fetal development in pregnant mice fed a 2% L-Trp or 5% L-Trp diet (experiment 1).

Non-proteinaceous form L-Trp concentrations in the maternal liver, blood, placenta, and fetus (experiment 1)
Non-proteinaceous form of L-Trp concentrations in the maternal liver and total blood, placenta and fetus did not differ between the control and 2% L-Trp groups at E12.5 (Fig. 2A–D) and E17.5 (Fig. 2E–H). Thus, mice fed the 2% L-Trp diet (3.3 g/kg BW/day) were able to maintain L-Trp concentration normal in the maternal body, placenta, and fetus.

Increase in Non-proteinaceous form of L-Trp concentrations in liver, blood, placenta, and fetus at E12.5 and E17.5 in the pregnant mice fed the 5% L-Trp diets (experiment 1).
In contrast, the non-proteinaceous form of L-Trp concentrations in the maternal liver, blood, placenta, and fetus were higher in the 5% L-Trp group than in the control and 2% L-Trp groups at E12.5 (Fig. 2A–D) and E17.5 (Fig. 2E–H). L-Trp accumulation was observed not only in maternal but also placental and fetal tissues in the 5% L-Trp group.
mRNA expression of L-Trp-degrading enzymes and LAT1 in the placenta (experiment 1)
The response patterns by L-Trp were different for Trp 2,3-dioxygenase (TDO), indoleamine 2,3-dioxygense 1 (IDO1), and IDO2 mRNA expression in the placenta (Table 4). TDO mRNA expression was not significantly different among the three groups at E12.5 and E17.5 (Table 4). IDO1 mRNA expression increased with dietary L-Trp content at E12.5 and E17.5 (Table 4). IDO2 mRNA expression increased in a dose-dependent manner of dietary L-Trp at E12.5; however, IDO2 mRNA expression was not observed at E17.5 (Table 4). There were no observed changes in mRNA expression for other enzymes involved in the L-Trp-kynurenine pathway (Table 4).
Composition of mRNA expression related to L-Trp-degrading enzymes in the placenta at E12.5 and E17.5 in pregnant mice fed control and high L-Trp diets (experiment 1).

In the NAD cycle and Nam catabolic pathway, mRNA expression of Nam mononucleotide adenylyltransferase 1 (NMNAT1), NAD kinase, and Nam N-methyltransferase (NMT) were induced according to L-Trp content in the diets at E12.5 (Table 4). NAD kinase mRNA expression was higher in the 5% L-Trp group than the control and 2% L-Trp groups (Table 4).
At E17.5, mRNA expression of Trp 5-hydroxylase 1 (TPH1), which is an initial enzyme involved the L-Trp-serotonin pathway, was significantly lower in the 2% L-Trp group and 5% L-Trp group than in the control group (Table 4). LAT1 mRNA expression was induced according to a L-Trp dose-dependent manner at E12.5. LAT1 mRNA expression was higher in the 5% L-Trp group than in the 2% L-Trp group (Table 4).
Spongiotrophoblast and labyrinth zone development (experiment 1)
The placenta supports fetal growth. Placental thickness is associated with fetal growth (ex, fetal body weight),21,22 so we measured the placental thickness of the control and 5% L-Trp groups. There was no significant difference in decidua basalis thickness between the control and 5% L-Trp groups at E12.5 (Figs. 3A–B and 4A). Placental spongiotrophoblast and labyrinth zone thicknesses were significantly thinner in the 5% L-Trp group than in the control group at E12.5 (Figs. 3A–B and 4B–C). The thicknesses of the spongiotrophoblast and labyrinth zone in the 5% L-Trp group were 49.1% and 69.5% of control thickness, respectively (Figs. 3B and 4C). There was no significant difference in the thickness of placental regions between the control and 5% L-Trp groups at E17.5 (Figs. 3C–D and 4D–F).
Pups body weight and death rate (experiment 2)
The body weights of pregnant mice were lower in the 5% L-Trp group than in the control and 2% L-Trp groups at E 18.5. (P < 0.05) (maternal body weight at E18.5: control group, 55.2 g ± 2.2; 2% L-Trp group, 55.1 g ± 5.0; 5% L-Trp group, 47.4 g ± 2.0). However, the body weights of pregnant mice in the 2% L-Trp and 5% L-Trp groups were not significantly different compared to the control group from E0.5 to E17.5 (data not shown). There was no incidence of abortion in control pregnant mice (plugging, n = 4; abortion, n = 0) and the 2% L-Trp group (plugging, n = 2; abortion, n = 0). However, the abortion percentage was 25.0% in the 5% L-Trp pregnant mice (plugging, n = 8; abortion, n = 2).
Pup weights at E18.5 were not significantly different between the control and 2% L-Trp groups (Table 5). All pups in the control and 2% L-Trp groups lived and moved immediately following caesarean delivery at E18.5. However, the pup weights were significantly lower in the 5% L-Trp group than in the control and 2% L-Trp groups (Table 5). Precise monitoring indicated that some pups in the 5% L-Trp group were born alive, but despite visible efforts to breathe, they became cyanotic and died within minutes. Other pups in the 5% L-Trp group responded to forceps irritation, but showed a very weak response compared to those in the control group. In contrast, the pups in the control and 2% L-Trp groups responded to forceps irritation and contracted their bodies and rolled over. At 1 hour after caesarean delivery, the death rate of pups was 3-fold higher in the 5% L-Trp group than in the control group (Table 5). Respiration frequency in living pups of the control and the 5% L-Trp groups was counted for 2 min. The incidence of pup respiration was 3-fold higher in the 5% L-Trp group than in the control group (Table 5). In the 2% L-Trp group, respiratory frequency did not differ from that of the control.
Pup body weight and death rate (experiment 2).

The data express dead rate until one hour from cesarean delivery;
the pregnant dams of each group took up the pups by cesarean delivery at E18.5. Immediately, numbers of respiration frequency of pups were monitored with a video camera.
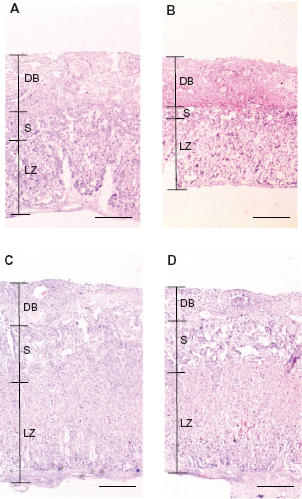
Retardation of the structural maturation of spongiotrophoblast and labyrinth zone in placentas of the pregnant mice fed the 5% L-Trp diet (experiment 1).
Lung morphometry of pups (experiment 2)
We had hypothesized that lungs of the pups in the 5% L-Trp group were damaged by excess L-Trp. The lungs of live and dead pups were macroscopically observed in the control and 5% L-Trp groups at E18.5. There were no abnormalities of the thoracic diaphragm and bronchi in the two groups. Lung weights were not significantly different between the control pups and the live pups in the 5% L-Trp group (data not shown). However, the lungs of dead pups were smaller in the 5% L-Trp group than in live pups of control and 5% L-Trp groups (Fig. 5A). The alveolar air space was narrower in dead pups of the 5% L-Trp group than in live pups of the control and the 5% L-Trp groups (P < 0.01) (Fig. 5B–C). Part of the lung of living pups in the 5% L-Trp group was collapsed (data not shown) and blood cells remained in the alveolar air space (Fig. 5C).

Decrease in thickness ratio of spongiotrophoblast and labyrinth zone in placentas of the pregnant mice fed the 5% L-Trp diet (experiment 1).

Histological comparison of pup's lungs at E18.5 (experiment 2).
Discussion
Understanding the safe consumption of nutrients is important in the prevention of certain health problems caused by excess intake of particular nutrients. Knowing the safe amount of L-Trp is particularly important because of the reported link between eosinophiliamyalgia syndrome and the ingestion of contaminated L-Trp preparations. 23 We previously reported that a high L-Trp diet (5% L-Trp) leads to a decrease in food intake and subsequent reduced body weight gain in weaned male rats. 9 However, feeding a 2% L-Trp diet does not show adverse effects for weaned male rats. 9 In addition, our group reported that supplementing healthy women with up to 5.0 g/day of L-Trp has no adverse effects. 24
Here, we clarified that less than 3.3 g L-Trp/kg BW/day during gestation can maintain the homeostasis of L-Trp concentration in the maternal body (ie, liver, blood), placenta, and fetus. Thus, consumption of up to 3.3 g L-Trp/kg BW/day was safe during gestation. However, 7.0 g L-Trp/kg BW/day was not safe. L-Trp accumulation in the maternal body, placenta, and fetus in pregnant mice fed a 5% L-Trp diet was observed. Thus, 7.0 g L-Trp/kg BW/day during gestation could not maintain the homeostasis of L-Trp concentration in the maternal body, placenta, and fetus. L-Trp is transported from maternal blood to fetus via the placenta, where L-Trp is transported by LAT1 and LAT2.25,26 The expression of LAT1 was higher in mice that had consumed the 5% L-Trp diet than in mice consuming control and 2% L-Trp diets. The increase in the transporter LAT1 may increase the ability to eliminate L-Trp from the body through the urine and amniotic fluid. The increase of L-Trp via LAT1 to the amniotic fluid in the 5% L-Trp group may worsen in addition to the increase in L-Trp intake from the 5% L-Trp diet.
Placental development is one of the most important factors for normal fetal growth. Birth weight is strongly positively correlated with placental weight. 27 The placenta of a low birth weight fetus induced by feeding of a low protein diet is light and shows a restricted ability for amino acid transport. 26 Here, placental weight at E17.5 did not differ between the control and 2% L-Trp groups, and pup body weights in the 2% L-Trp group recovered to control levels at E18.5. Further, the death rate of pups was 0.0% in the 2% L-Trp group. We suggest that the fetus could not grow normally because of the premature placenta of mice fed an excess L-Trp diet.
However, we did not measure the placental ability to transport nutrients, which should be examined in future studies.
The labyrinth zone of the placenta is the most important interface for exchanging nutrients between fetal and maternal tissues. The placenta of low birth weight newborns is light and shows retardation of labyrinth zone development, which is associated with fetal growth restriction. 22 The low birth weight newborn death rate is higher because of problems with respiration, including collapsed lungs. 28 The placenta was thinner and lighter in the 5% L-Trp group than in the control group. This suggests that fetuses could not grow normally because of the premature placenta. Thus, the low birth weight newborns in the 5% L-Trp group died, likely because of lung collapse.
The fate of L-Trp in the placenta may explain the resulting toxicity of excess L-Trp. IDO1 has been implicated in contributing to immunotolerance in early pregnancy to deplete L-Trp. 29 It has been reported that IDO1 and TDO is expressed in the mouse placenta 30 and that kynurenine 3-monooxygenase (KMO) is expressed in the human placenta. 31 L-Trp metabolism is unique in the placenta. TDO and IDO1 mRNA is strongly expressed at mid-gestation, but that expression becomes very weak before birth in mice. 30 TDO mRNA expression is strongly expressed from early to mid-gestation (about E10.5), but that expression becomes very weak after E12.5 until birth in mice. 30 There have been no reports regarding the effect on IDO1 and TDO expression in a high L-Trp diet in pregnant rodent.12,13 We showed that mRNA expression of enzymes of L-Trp degradation in the placenta when the dam was fed 2% L-Trp or 5% L-Trp diets. TDO mRNA expression was not different in the control, 2% L-Trp and 5% L-Trp groups. In contrast, IDO1 mRNA expression increased according to L-Trp content in the diets. IDO1 is known to be induced by various compounds, such as interferon-γ 32 and lipopolysaccharide. 33 Induced IDO1 expression may not be induced by L-Trp in the placenta. L-Trp degrades to N-formylkynurenine, which then is converted to kynurenine by formylase. KMO, kynureninase (KYNase) and kynurenine aminotransferase 2 (KAT2) mRNA expression did not change at E12.5 and E17.5. These results suggest that kynurenine accumulates in the placenta of mice fed a high L-Trp diet.
The role of IDO2 is poorly understood. Here, IDO2 mRNA expression was not dependent on L-Trp content in the diet at E12.5. However, the expression was not different in the control, 2% L-Trp, and 5% L-Trp groups at E17.5. Thus, IDO 2 mRNA expression may be controlled by factors other than L-Trp.
NMT catabolizes the reaction of Nam to N 1 -methylnicotinamide (MNA). We showed that NMT mRNA expression increased in a dose-dependent manner with L-Trp. AOX mRNA expression in the placenta at E12.5 did not change. This implies the possibility of MNA accumulation in the placenta at E12.5. In recent years, the physiological roles of MNA have been reported, such as antiinflammatory action in skin diseases, induction of prostacyclin synthesis via COX-2, aortal endothelium protection in diabetes and hypertriglyceridaemia, and increased survival rate of diabetic rats.34–37 We were unable to determine whether MNA accumulation leads to the inhibition of placental development and intend to focus further investigation in this area.
An increase of serotonin level in placenta has been previously reported. 15 We showed that TPH1 mRNA expression was lower in the 2% L-Trp and 5% L-Trp groups than in the control group at E17.5. However, serotonin level in placenta was not significantly different between control and 5% L-Trp group at E18.5 (Table S1).
Conclusion
We demonstrated that mice fed 3.3 g L-Trp/kg BW/day during gestation were able to maintain homeostasis in the maternal body, placenta and fetus; thus, no adverse effects were observed up to feeding 3.3 g L-Trp/kg BW/day even in gestation. However, mice fed 7.0 g L-Trp/kg BW/day during gestation led to the accumulation of non-proteinaceous form of L-Trp in the maternal body, placenta, and fetus, which may explain the reduction of placental development and fetal growth and the higher death rate of pup of the maternal.
Author Contributions
AT, TF, MS and KS designed the research. AT, CN conducted the research. AT and KS analyzed data. AT and KS drafted the manuscript. All authors reviewed and approved of the final manuscript.
Funding
This investigation was part of the project “Studies on the nutritional evaluation of amino acids and B-group vitamins” (principal investigator, Katsumi Shibata), which was supported by a Research Grant for Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.
Footnotes
Supplementary table
Serotonin content in placenta at E18.5.

| Control | 5% L-Trp | |
|---|---|---|
| Serotonin (nmol/placenta) | 1.07 ± 0.05 | 1.13 ± 0.20 |
| (n = 3) | (n = 6) |