
Abstract
Literature is reviewed on the spatial distribution of the eggs and neonate larvae of codling moth on apple trees in relation to research conducted in Nelson, New Zealand. At Nelson, oviposition increased with height and was greater in the north and east of the trees and in those with greater fruit load in some seasons, which matches published reports. All publications and the research recorded high percentages of eggs laid singly within 10–15 cm of the fruit, with most eggs on leaves even within fruit clusters; oviposition on fruit clusters of different sizes was nonrandom because more eggs were laid on those with more fruit, but the aggregation of both per cluster and within clusters was even greater than that caused by the fruit number alone. Oviposition at random with respect to the fruit occurred only at very low population density. The choice of oviposition site between fruit and the adaxial leaf surface and abaxial leaf surface (AbLS) was variable and cultivar related. Cultivars on which eggs predominated on the AbLS were less frequent and characterized by low trichome density. In the literature, neonate larvae from eggs on the AbLS suffered greater mortality, as did those in Nelson that hatched more distant from the fruit. This review discusses the interaction between these distribution characteristics and species-specific host–plant volatiles, egg adhesion to plant surfaces, oviposition deterrents, predation, and their relevance to pest management.
Introduction
In ecological terms, the codling moth,
Fruit odor from apples has a wide range of effects on codling moth behavior: adult activity is increased8,16 as is virgin female pheromone production;
8
it promotes upwind orientation of larvae and adults to the odor source,7,8,17 although not at petal fall when it is repellent;
18
and oviposition is stimulated by apple odor.8,9,16 The first active fruit volatile identified was the terpene α-farnesene, as an attractant to neonate larvae
7
,19–21 and an oviposition stimulant;7,22,23 this compound was later shown to be a female attractant
24
and a stimulant of pheromone production by virgin females.
23
But it is now clear that codling moth responds to a blend of fruit volatiles;8,9,23,25–29 males are attracted to (
These phytochemical relationships have a major influence on the distribution of codling moth eggs by the ovipositing female. A clear manifestation of this is that eggs are laid singly on or close to the fruit, 33 often including >90% laid within 10 cm.4,16,34 Geier 4 never found eggs on apple trees without fruit, and several authors have found a positive relationship between fruit load of individual trees and their codling moth population or infestation.35–37 The laying of eggs singly reduces larval competition for fruit and enables maximum use of the available host fruit by the hatching larvae.
Analysis of the distribution of eggs within fruit clusters of different sizes has produced conflicting results. Geier
4
concluded that eggs were laid at random with respect to the fruit and that this was sufficient to account for both the increase in egg laying on clusters with greater fruit numbers and the distribution of eggs within clusters of different sizes; these conclusions referred to a codling moth population at a very low density (0.14 eggs/fruit cluster). However, studies by Wood
37
suggested that, at higher population densities of codling moth, some contagion of oviposition occurred. Stronger evidence for this aggregation was found in a high-density codling moth population by Jackson,
1
who discussed the relationship between the results of all three studies. In his research, the number of eggs per fruit cluster increased with cluster size, but the number of eggs per fruit decreased. He extended the concept of each fruit being an independent point source of attraction for female moths (as proposed by Geier)
4
to each having a
The relationship of codling moth with its host plant is also expressed in the sites chosen by the female when laying eggs near the fruit,
It has been reported for many years that codling moth is more active or causes more damage in the upper parts compared with the lower parts of its host trees.4,36,37,45–49 Some of these reports indicated that this was due to reduced spray coverage in the upper parts of the trees,4,37 but others found that such height effects occurred in the absence of spraying.47,49 Blomefield et al 34 recorded greater oviposition in the lower area of apple trees during the first generation of codling moth in South Africa, but a change to greater oviposition in the upper areas in the second and third generations. When studying oviposition by a high-density codling moth population on 12–20 m tall unmanaged apple trees, Jackson 1 failed to detect any impact of height on the number of eggs per sample.
There is also some variability in the effects of aspect within the tree on oviposition by codling moth. Most authors report no or minor effects of aspect,4,34,37,43,50 but Stoeckli et al 51 discovered reduced larval infestation, and hence density, on the north-facing side of apple trees compared with the south or east, and this is likely to reflect oviposition differences.
The need to understand the distribution of codling moth eggs in commercial apple trees (
The current study presents previously unpublished research on the spatial distribution of codling moth oviposition and larval fruit entries on apple trees in Nelson, New Zealand. Temporal distribution is included, but limited to data on the relationship between fruit size and the timing of egg laying and fruit entry by neonate larvae. The data were obtained in association with a life table study of a univoltine population of the species over eight generations from 1967 to 1975, 63 and the results complement earlier descriptions of the distributions of fifth-instar larvae leaving the fruit and cocooning on the trees. 64 Related world literature relevant to codling moth spatial distribution in apple trees is reviewed.
Materials and Methods
The research site
A full description of the Appleby Research Orchard, Nelson (41°17′S), where this research was conducted, is given by Collyer and van Geldermalsen, 65 and the study area specifically used for codling moth research from 1967 to 1975 is described by Wearing. 63 The latter mainly comprised three contiguous blocks of mature (50-year old) ‘Delicious’ trees (blocks A, B, and C), and one neighboring block each of ‘Cox's Orange Pippin’ (‘Cox’) and ‘Dunn's Favourite’ (‘Dunns’); all at 5.5 m x 5.5 m spacing, 330 trees per ha, growing in a grass/clover sward. Block A was not sprayed with insecticide in the 1967/1968, 1968/1969, 1969/1970, and 1974/1975 seasons; from the 1970/1971 to 1973/1974 seasons, an additional unsprayed ‘Delicious’ block D and ‘Red Delicious’ block E were substituted because block A was sprayed; the only insecticide applied to block A, and to blocks B, C, ‘Cox’, and ‘Dunns’ from 1967 to 1975, was ryania (Ryanicide 50® or Ryanicide 100®; S. B. Penick & Co.) or codling moth granulosis virus in various schedules as specified by Wearing. 63 The trees were trained to a height of 3.5 m. Neither chemical nor hand thinning was carried out during the study period, and the harvested crop averaged between 600 and 2000 fruits per tree.
Egg and larval sampling on fruit clusters
A specific sampling programme was designed to investigate the distribution of codling moth life stages in the trees. This was in addition to the population sampling detailed by Wearing.
63
Based on the sampling method of Wood,
37
all set fruit clusters (one or more fruit and the surrounding leaves arising from one bud) were counted on the sample trees at the beginning of November each season (1967/1968–1970/1971). The total number of fruit clusters on each tree was divided by 36 to obtain a number
Destructive sampling to estimate egg and preentry larval mortality
Egg and neonate larval distribution data were also collected during destructive sampling of foliage and fruit to estimate egg and larval mortality before fruit entry.
63
Stratified random samples were taken in January each year in the lower (7% of samples), middle (21%), and upper parts (72%) of the trees, immediately after the peak of larval hatching. The percentage distribution of samples was based on the natural distribution of eggs determined from systematic sampling in 1967/1968. From a random starting point within each stratum, every fifth fruit cluster was removed and searched. When an egg was found, the whole fruiting arm was sampled for eggs and larval entries. Sampling of every fifth cluster was then resumed until at least 120 eggs had been found in each block.
63
Results are presented from sampling blocks A, B, C, D, and ‘Dunns’ in 1971/1972 and 1972/1973. Details of the location of each egg were recorded as follows: site (AdLS, AbLS, fruit, and wood), distance to the nearest fruit (cm, 1972/1973), and the number of fruits in the associated cluster and cluster group. A cluster group refers to the fruit clusters whose foliage and/or fruit were close enough to touch or overlap, thereby allowing the potential for mixing of fruit volatiles from the combined fruit numbers to influence the ovipositing female (see Ref. 1 for definition of
Egg distribution in fecundity cages
Wearing 63 estimated the fecundity (eggs laid per female) of codling moth in each year of his population study by caging newly emerged wild adult females with one or two wild males in sleeve cages on the fruiting branches of the trees. A full description of the caging methodology and cultivars used is given in the study by Wearing and Ferguson. 5 Once the female died, she was dissected to determine her mated status, and the cage and its contents were taken to the laboratory to examine all surfaces for eggs. The site of all eggs was noted, including the numbers on individual leaves and fruit (blocks A, B, C, D, ‘Dunns’, and ‘Cox’). In 1972/1973, 20 cages were placed over branches without fruit for comparison with those containing fruit, and in the latter case, the distance of each egg to its nearest fruit was recorded.
Tree to tree distribution and fruit load
In addition to the tagged cluster sampling of the ‘Delicious’ trees in blocks A, B, and C already described, the windfalls and entire crop at the harvest of sample trees (usually 12 per block) were examined for neonate and other larval fruit entries as a part of the life table study from 1967 to 1975 (21 blocks).
63
This provided the data for examining by regression whether the codling moth population (as measured by the number of neonatal larval entries) within a tree was related to the number of fruits in that tree. Population density was known to vary between the blocks that received different treatments,
63
and hence, the annual block data obtained were analyzed by regression independently. While an ideal analysis would have sought to determine the relationship between the egg numbers per tree and the fruit numbers per tree, this was not practical. Analysis of the life table data
63
showed that the estimated neonate larval entries per tree (
Neonate larval entries distribution within the trees
As a result of codling moth oviposition and larval attraction to the fruit, the distribution of neonate larval entries within apple trees is inevitably linked to the distribution of the fruit. In 1967/68 when population density was low, measurements were taken of the distances (cm) from a randomly-selected neonate larval entry to all its neighbours on the same tree. This procedure was replicated four times for unsprayed ‘Delicious’ trees and provided the data on distances to both the nearest neighbor and all the within-tree neighbors. Separate neonates with entries on the same fruit were recorded as having zero distance from one another.
Standard nearest-neighbor analyses indicated that the entries were not random but were aggregated within the areas of the trees. However, correct spatial analyses of these data would require knowledge of the structure and volume of the trees and the distributions of the fruit within them. Moreover, methods for three-dimensional recording of the location of the fruit and their infestation are now available to improve data collection and analysis. 66 In the absence of these methods and only as a preliminary step, the distributions of neonate larval entries and their nearest neighbors were plotted for each tree and compared with a Poisson distribution.
Data analysis
Regression analyses were used to determine the relationships between the following: tree fruit load and codling moth larval fruit entries, height within the trees and egg/larval density per fruit cluster and per fruit, and the proportion of neonate larvae entering the fruit having hatched at different distances away. χ2 analyses were used to compare the observed distribution data with those expected from a Poisson distribution for the following: larval density in different tree aspects, distances to the nearest neighbors of larval entries, eggs and neonate larval entries per fruit cluster and per fruit for clusters of different sizes, and eggs and neonate larval entries per individual fruit. A χ2 homogeneity test was used for the analysis of the distribution of eggs and neonate larval entries per fruit
Results
Temporal distribution
Codling moth in Nelson is univoltine but with a small partial second generation comprising 1.6%–2.1% of the population in some seasons. 37 In the univoltine 1967/1968 season, first eggs were sampled in mid-November and the last in early March, with peak (.80%) oviposition in January and early February, when fruit diameter averaged 4.3–5.3 cm (Fig. 1).
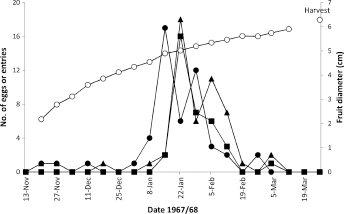
Temporal distribution of total egg laying (•), egg hatching (Δ), and neonate larval fruit entry (■) by codling moth on 576 fruit clusters of 16 ‘Delicious’ apple trees in relation to the mean fruit size (O) in the same clusters at nelson, new Zealand, in 1967/1968.
Egg numbers were too low to analyze site selection or the changes in site selection by the ovipositing females over the season, except to report that eggs were not recorded on the fruit until January 19, 1968; the overall distribution was 10.9% on fruit, 34.8% on the AdLS, and 54.3% on the AbLS (
Sampling of fruit clusters for neonate larval entries in the following three seasons (1968–1971) showed similar timing of the peak of fruit entries (and hence peak oviposition); it occurred predominantly in January and February each year (Fig. 2) at a fruit size range similar to that in 1967/1968. The codling moth population was at high density in 1970/1971, and eggs on the fruit were found from November 20, 1970, until March 5, 1971 (Fig. 2). As a result, egg laying was associated with a larger size range of fruit than recorded in 1967/1968.
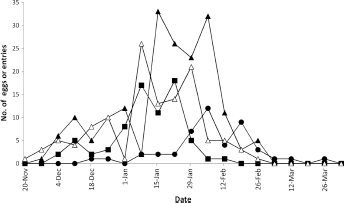
Temporal distribution of total eggs laid on the fruit (Δ, 1970/1971) and neonate larval fruit entries by codling moth on 360 fruit clusters of 10 ‘Delicious’ apple trees in block a at nelson, new Zealand, in 1968/1969 (●), 1969/1970 (■), and 1970/1971 (▲).
Spatial distribution–-fruit load
The relationship between the total fruit (including windfalls) of individual trees and their total neonate larval entries (and hence oviposition) was highly variable between years and blocks (Table 1). Of the 21 blocks/seasons analyzed, 9 blocks were found to have a positive relationship (
Regressions of neonate larval entries per tree on the total number of fruits (including windfalls) per tree for 21 blocks of ‘Delicious’ apples and the estimated egg density per tree in the same blocks 1967–1975 at nelson, new Zealand.
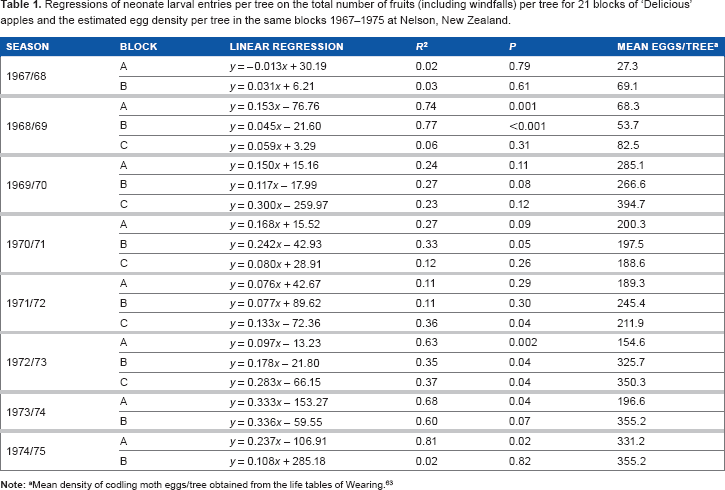
Mean density of codling moth eggs/tree obtained from the life tables of Wearing. 63
Spatial distribution–-height
Every year, most of the fruit clusters were in the height range of 1–2 m above the ground (Fig. 3), and the peak numbers of clusters with >1 fruit were located higher in the trees (>1.5–2 m) than those with only one fruit (>1–1.5 m; Fig. 3).
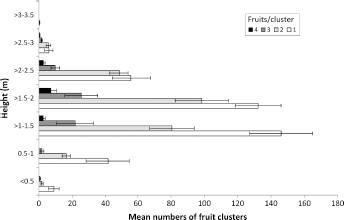
Mean numbers of fruit clusters of different sizes at different heights in ‘Delicious’ apple trees, standardized for a total of 720 clusters sampled using a systematic sampling method of 36 per tree on 20 trees on 20 January each year 1967/1968 to 1970/1971 at Nelson, New Zealand. Bars are ±1 SEM.
The number of eggs
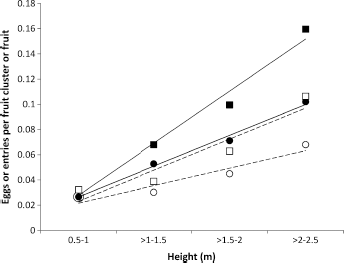
Regressions of the codling moth eggs laid on the height of fruit clusters (–-■–-) and their fruit (–-●–-) in ‘Delicious’ apple trees in 1967/1968 and the equivalent regressions for neonate larval fruit entries on the same fruit clusters (- -□- -) and fruit (- -○- -) (■:
In each of the subsequent years, the neonate larval fruit entries per fruit cluster and per fruit also increased with cluster height, and this effect increased with population density (Fig. 5). With significant numbers of fruit clusters above 2.5 m in 1968/1969 and 1969/1970, it was possible to show that the greatest entry density per fruit cluster occurred at the tops of the trees in those years.
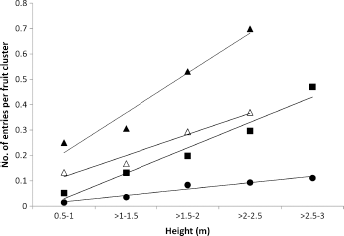
Relationship between the height of fruit clusters in ‘Delicious’ apple trees and the neonate larval fruit entries per fruit cluster in 1968/1969 (●:
Eggs were not recorded on the leaves in the systematic sampling of 1970/1971, but records were obtained for eggs laid on the fruit (
Spatial distribution–-aspect
Egg numbers sampled in 1967/1968 were insufficient to determine if aspect within the tree affected their distribution by the ovipositing females. However, sampling of neonate larval entries provided four years of aspect data (Fig. 6).
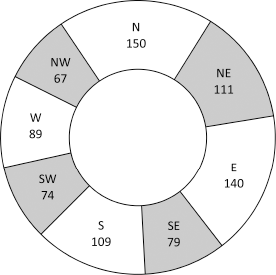
Distribution of the aspect of neonate larval entries on a total of 3096 fruit clusters of ‘Delicious’ apple trees sampled over four years 1967/1968–1970/1971 at nelson, New Zealand.
The aspect distribution of larval entries did not differ significantly between years (χ2 analysis four years x 8 sectors, χ2 = 28.98, 21 degrees of freedom (
Spatial distribution–-neonate larval entries within the tree
The distributions of distances between neonate larval entries and their nearest neighbors were similar in all four ‘Delicious’ sample trees, exemplified by the tree in Figure 7. The nearest neighbors were not distributed randomly, with more than expected in the range 0–4 cm and above 10 cm, and less than expected at 6 cm, close to the mean (χ2 = 2.74 x 1011, 19
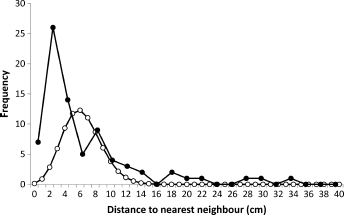
Distribution of distances to the nearest neighbor for neonate larval entries (
Distance of eggs from the nearest fruit
Measurement of the distance of eggs to their nearest apple was undertaken for 46 of the eggs found on the ‘Delicious’ trees during the systematic sampling of 1967/1968. More than 50% were within 3 cm and 96% were within 12 cm of the fruit. Similar results were obtained with greater numbers of eggs in 1972/1973, using both destructive sampling and fecundity cages (Fig. 8).
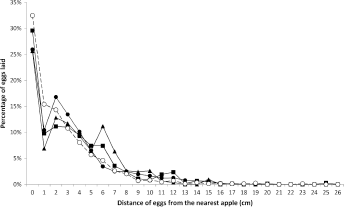
Distance of codling moth eggs from the nearest fruit when laid in fecundity cages on apple trees of the cultivars ‘cox’ (■,
For eggs recorded on ‘Delicious’ trees during destructive sampling, 62.2% were within 2 cm of the fruit and 98.7% were within 12 cm. Of the 844 eggs recorded, 274 were on the fruit themselves, and an additional 130 eggs were within 1 cm. In the fecundity cages, 50.5%, 53.2%, and 45.5% of eggs were within 2 cm, and 97.9%, 97.8%, and 98.9% were within 12 cm on ‘Cox’, ‘Delicious’, and ‘Dunns’, respectively. In 1972/1973, 11 of the 20 fecundity cages operated on ‘Delicious’ branches without fruit had females that mated, but they produced a mean of only 2.5 (95% CL 0.5–7.3; range 0–31) eggs compared with a mean of 40.7 (95% CL 32.4–51.2; range 0–197) by 167 females in cages on branches with apples [detransformed mean values from log (
The 1967/1968 mean population density on ‘Delicious’ trees was only 48.2 eggs per tree, 63 with little competition for fruit clusters between hatching larvae. The labeled cluster sampling data enabled preliminary assessment of the survival of larvae hatching at different distances from their nearest fruit. Survival declined with increasing distance (Fig. 9).
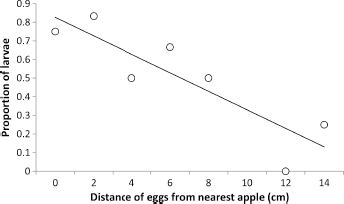
Proportion of neonate codling moth larvae (
Spatial distribution–-site selection for oviposition
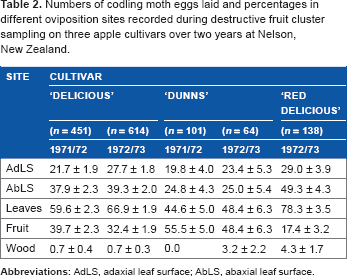
Sampling confirmed that female codling moths laid more of their eggs on the leaves than the fruit of the cultivars ‘Delicious’ and ‘Red Delicious’ (Table 2), as in the systematic sampling data of 1967/1968 previously noted (see “Results: Temporal distribution”). This was also the result in the fecundity cages from 1967 to 1973, despite the artificial conditions, and this included the additional cultivars ‘Cox’, ‘Jonathan’, ‘Kidds D8’, and ‘Sturmer Pippin’. The only exceptions were the cultivars ‘Dunns’ (Table 2) and ‘King of Tompkins County’ (in fecundity cages, 37.7% on leaves and 62.3% on fruit) whose oviposition on the fruit was equal to or greater than that on the leaves. The cluster sampling consistently showed that a higher proportion of eggs were laid on the AbLS than on the AdLS of ‘Delicious’, ‘Dunns’, and ‘Red Delicious’ cultivars (Table 2), as noted also from the systematic sampling of ‘Delicious’ (see “Results: Temporal distribution”).
Numbers of codling moth eggs laid and percentages in different oviposition sites recorded during destructive fruit cluster sampling on three apple cultivars over two years at Nelson, New Zealand.
Position of eggs on leaves
Systematic sampling in 1967/1968 included 40 eggs on leaves whose details were recorded. The mean leaf length selected for oviposition was 6.93 ± 0.39 cm (range 2.5–12.5 cm). Only one of the leaves used for oviposition was horizontal compared with 25 that were vertical, with the remainder at intermediate angles (overall mean = 72.5° ± 3.9°). The eggs were laid at a mean of 2.66 ± 0.30 cm (range 0.3–7.5 cm) from the base of the leaf and a proportional mean distance of 0.56 ± 0.04 (range 0.10–1.00) from the midrib to the leaf edge.
Distribution of eggs and larval entries on fruit clusters
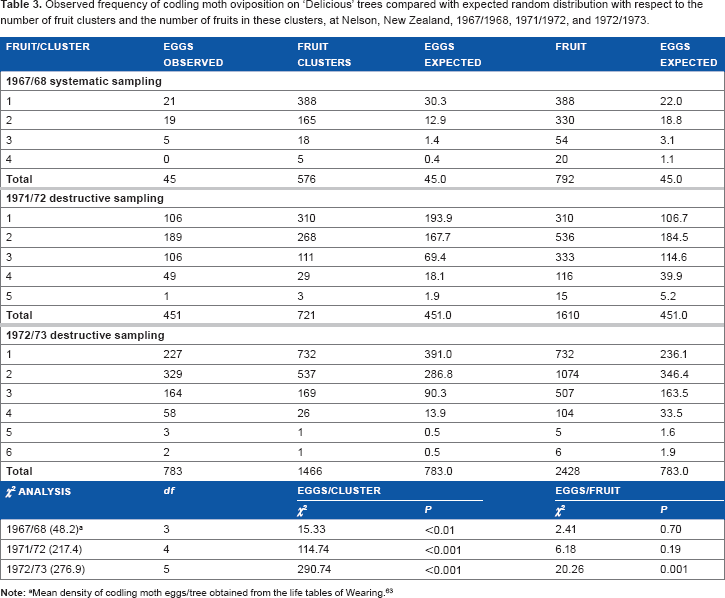
The distributions of eggs on ‘Delicious’ fruit clusters of different sizes are shown in Table 3. It is not possible to compare statistically the density of the egg populations in the three years because of the changing sampling practices, but the egg sampling in 1967/1968 found only 0.08 eggs per fruit cluster, whereas destructive sampling in 1971/1972 and 1972/1973 yielded 0.63 and 0.53 eggs per fruit cluster, respectively. The higher egg density in the two later years reflected the known increase in population densities (compared with 1967/1968) from life table studies (see Table 3 and “Materials and methods” section).
Observed frequency of codling moth oviposition on ‘Delicious’ trees compared with expected random distribution with respect to the number of fruit clusters and the number of fruits in these clusters, at nelson, New Zealand, 1967/1968, 1971/1972, and 1972/1973.
Mean density of codling moth eggs/tree obtained from the life tables of Wearing. 63
There was a strong agreement between the three fruit cluster data sets. The distribution of eggs with respect to fruit clusters was not random (
The increase in egg numbers with increasing numbers of fruit per cluster every year was exponential (Fig. 10). On the other hand, egg numbers per fruit either failed to increase with increasing cluster size (
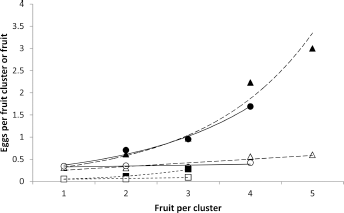
Codling moth egg numbers per fruit cluster (solid symbols) and per fruit (open symbols) laid on ‘Delicious’ fruit clusters of different sizes in 1967/1968 (- - ■, □- -), 1971/1972 (–-●, ○–-), and 1972/1973 (- -▲, Δ- -). ■:
When the 1971/1972 and 1972/1973 fruit clusters were amalgamated into fruit groups (see “Materials and methods” section), the relationships between the size of the groups and egg deposition were similar to those shown in Figure 10. In 1971/1972, eggs per fruit group (up to 15 fruits/group) rose exponentially with increasing group size (
χ2 analyses were also applied to neonate larval entries per fruit cluster and per fruit (Table 4). All analyses showed that the entries per fruit cluster were nonrandom as found for oviposition. The distributions of larval entries derived from systematic sampling over four years showed that the entries were consistently random with respect to the fruit. However, the distributions of entries per fruit obtained from destructive sampling did not have a good fit to Poisson, suggesting that bias had occurred during sampling. Such an effect may have also occurred during egg sampling in 1972/1973 (Table 3). The destructive sampling method was designed principally to measure egg and neonate larval pre-entry mortality; it focused sampling on areas of the trees where eggs and entries were found, and avoided areas where they were absent (see “Materials and methods” section). This practice, and the deliberate bias toward the middle and upper strata of the trees during destructive sampling, probably account for the nonrandom distribution of eggs and entries per fruit. Differences in population density between years are unlikely to account for these changes in larval entry distribution because the four years of systematic sampling included the lowest (1967/1968) and the highest (1969/1970) population densities (Table 4).
Observed frequency of codling moth neonate larval fruit entries on ‘Delicious’ trees compared with expected random distribution with respect to the number of fruit clusters and the number of fruits in these clusters, at nelson, New Zealand, 1967/1968, 1968/1969, 1970/1971, 1971/1972, and 1972/1973.
Mean density of codling moth eggs/tree obtained from the life tables of Wearing. 63
Distribution of eggs and larval entries within fruit clusters
Oviposition within clusters of different sizes is presented in Table 5. Systematic sampling in 1967/1968 gave observed distributions that were primarily a good fit with Poisson. The 2-fruit/cluster data were exceptional in that there were too many clusters in the 0 and 2 eggs/fruit cluster categories and too few with 1 egg/fruit cluster (
Observed frequency of codling moth oviposition on ‘Delicious’ trees compared with the expected random distribution within fruit clusters of different sizes, at nelson, New Zealand, 1967/1968, 1971/1972, and 1972/1973.
The analysis of neonate larval entries within fruit clusters was possible with data replicated over four years using unbiased systematic sampling (Table 6). With the exception of the data of the 4-fruit/cluster class that were few in number, the results indicated that nonrandom distribution of entries within clusters was the norm, especially in 1969/1970 and 1970/1971 when population density was much higher. Random distributions were found only in 1968/1969 and in clusters with one fruit in 1967/1968. In the absence of any change in the sampling method, these results suggest that changes of distribution toward aggregation within clusters over the four years were density dependent. As for the egg data, the departures from nonrandom were associated with too few entries in the 1 entry/cluster class, and too many in the 0 and 2+ classes.
Observed frequency of codling moth larval entries per fruit cluster (E, 0–4+) on ‘Delicious’ trees compared with the expected random distribution within fruit clusters of different sizes (F, 1–4+), at nelson, New Zealand, 1967–1973.
Fruit/cluster (F) and eggs/cluster (E).
In parenthesis refers only to 1971/1972 and/or 1972/1973.
The analyses of the neonate larval entries data from destructive sampling in 1971/1972 and 1972/1973 (Table 6) gave results similar to those from systematic sampling when population densities were also high (1969/1970 and 1970/1971). In addition, the destructive sampling provided records for clusters as big as 6 fruit (truncated to 4+ fruit/cluster for the analyses) and instances of up to 6–12 entries on a single cluster (truncated to 6+ entries/cluster for the analyses). Overall, the analyses confirmed that aggregation usually occurred within the fruit clusters and suggested that this failed to occur only when population densities were low or data were sparse.
Distribution of eggs and neonate larvae on individual fruit
In 1972/1973, the destructive sampling provided sufficient data on the numbers of eggs and entries on individual fruit that their distribution could be analyzed. The samples from unsprayed ‘Delicious’ trees revealed that the distributions of eggs and neonate larval entries were random on the individual fruit (eggs: χ2 = 0.60, 3
Unsprayed block E (‘Red Delicious’) was also sampled destructively in 1972/1973 and confirmed that egg distribution on individual fruit was random (χ2 = 0.03, 3
Analysis of the distribution of eggs on the individual leaves and fruits of the fecundity cages (1970/1971 and 1972/1973) showed that they were very highly aggregated on both substrates. Despite the similarity of egg laying distances to the fruit in cages compared to the wild population (Fig. 8), the ovipositing females confined in cages did not lay randomly on individual leaves or fruit, regardless of whether they were in sprayed or unsprayed blocks (Table 7). On fruits, there were too many without eggs (62%), too few with 1–5 eggs, and too many with >5 eggs compared with a random distribution. On leaves, there were too many without eggs (86%), too few with 1–2 eggs, and too many with >2 eggs compared with a random distribution.
Distribution of eggs on the individual fruit and leaves in fecundity cages on unsprayed and ryania-sprayed ‘Delicious’ trees in 1970/1971 and the results of χ2 analysis when compared with a random distribution.
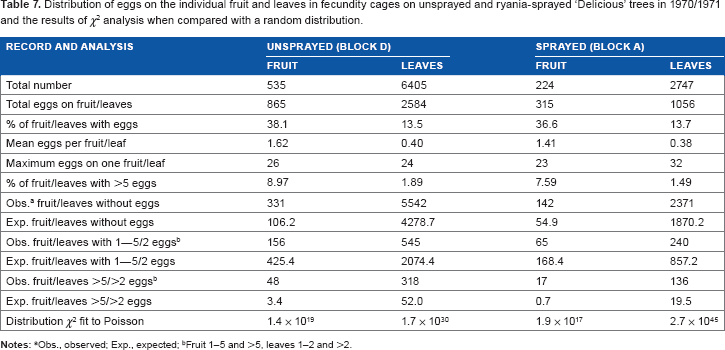
Obs., observed; Exp., expected;
Fruit 1–5 and >5, leaves 1–2 and >2.
Discussion
Temporal distribution
The temporal distribution of codling moth in relation to fruit size (Figs. 1 and 2) showed that in Nelson, New Zealand, the females mainly oviposited on ‘Delicious’ when the apples were 4.3–5.3 cm in diameter. However, first and last eggs were recorded when the fruit was about 2.2 and 5.6 cm in diameter, respectively; first eggs to be laid on the fruit were recorded on 20 November (in 1970). In South Africa, where codling moth is multivoltine, Blomefield et al 34 repeatedly recorded the first eggs on the fruit in the period 19–24 November (1986, 1987, and 1990), and in 1990, the fruit size at that time was 2.66 cm in diameter. As in New Zealand, oviposition occurred in response to a wide range of fruit sizes through the season.
Apple production of α-farnesene, an oviposition stimulant and a larval attractant of codling moth, was shown by Sutherland et al 67 to increase steadily through the season, but production per unit area of fruit surface (including ‘Delicious’) was maximal at fruit set, dropped to a minimum immediately after (late November), and had a further peak over January/February. Although the fruit set peak can be seen as coinciding with the oviposition of the first generation in South Africa and the later peak with the oviposition of the later generations (and with the single generation in Nelson), any causal association between these events remains to be proven. 67 Vallat and Dorn 18 found peaks of volatile attractant chemicals in June and August in the northern hemisphere, when apples were 4.5 and 6.5 cm in diameter, and suggested a relationship to peaks of codling moth oviposition. However, oviposition close to or on the fruit throughout the season suggests that attraction/stimulation is adequate at all times and probably involves a blend of volatile chemicals;9,18,25 the variations in α-farnesene production per unit area may be more important for short-range fruit location by the neonate larvae 7 than for oviposition.
Spatial distribution in the trees
Analysis confirmed some positive relationships between the number of fruits in a tree and their infestation by codling moth (Table 1), as reported by several authors.35–37 However, this was not consistent, despite confining the regression analyses to blocks of trees subject to the same treatment. Wearing
63
showed that immigration was a key factor in the population dynamics of codling moth in the study site, and trees within the blocks varied in their distances from the source of immigrants. Another key factor was variation in the fecundity of codling moth,
63
particularly individual females (0–284 eggs), such that similar numbers of moths per tree could result in different egg numbers per tree, especially at low population density. Both these factors could have reduced the strength of the relationship between fruit load and within-tree egg density, particularly as codling moth adults are mainly sedentary and females lay most of their eggs on one or two neighboring trees.4,68 Edge trees can also act as a
In controlled experiments using the same number of codling moth females released onto caged trees, Brahim et al 69 attributed the greater oviposition on ‘Golden Delicious’ compared to that on ‘Starkrimson’ to the higher fruit load carried by the former. However, they also recorded the behavior of females when ovipositing on these apple cultivars, and this included greater speed of locomotion, greater distance walked, and more time taken to lay each egg on ‘Golden Delicious’.
The increases in codling moth oviposition and neonate larval entries with height in the 3.5 m high ‘Delicious’ trees of the present study were unequivocal (Figs. 4 and 5). These occurred similarly on unsprayed trees and those sprayed with ryania, and confirmed the published results,37,45,47,49 for unsprayed apple trees, and the results of Geier
4
and Wood
37
for ryania-sprayed trees. At least a part of this effect can be attributed to the measured increase with height of the proportion of fruit clusters with >1 fruit (Fig. 3), but even when corrected for cluster size, the data showed an increase with height in the number of eggs and entries per fruit (Fig. 4). Direct observations have found that adult codling moths occur in greater numbers in the upper parts of apple trees,46,70 and this leads to more mating activity,
60
and higher catches in bait traps.
47
In the ‘Delicious’ trees of the present study orchard, Wearing
63
rotated alternate bait traps (
But not all authors have recorded increased oviposition and larval fruit entries with height. 41 Jackson 1 found no such effect on unmanaged apple trees 12 and 20 m tall containing a codling moth population producing 1.96 eggs/fruit cluster, a mean of more than two larval entries per apple, and 95% infestation of the crop. These conditions are very different to those in the present study, and it is conceivable that the population density was so high that height gradients were overridden at the limits of the carrying capacity of the crop. Stoeckli et al 51 studied the effects of height on codling moth infestation in dwarf trees averaging 2.5 m tall. Despite their small stature, significant height effects occurred, with greatest infestation at either the top or middle stratum at different locations. It is not possible to know if these effects reflected differences in fruit density as this was not measured.
The evidence from the current study that eggs laid
In the northern hemisphere, Wei et al 41 and Stoeckli et al 51 found reduced codling moth oviposition and infestation, respectively, on the north-facing side compared with the south- and east-facing sides of apple trees, and MacLellan 36 similarly reported an early-season ovipositional preference for the southeast aspect. On the other hand, Blago and Dickler 50 and Wei et al 43 found no significant relationship within apple trees between oviposition and aspect, and this has also been more commonly reported in the southern hemisphere.4,34,37 The present findings that larval entries were greater on the north- and east-facing aspect compared with the south- and west-facing aspect of the trees (Fig. 6) agree with the studies by Stoeckli et al 51 and Wei et al 41 in the opposing northern hemisphere. With respect to north and south, both probably indicate an oviposition response to the warmer areas of the tree, as shown by Kührt et al, 71 which may include response to more advanced fruit development and associated greater α-farnesene production. 67 In Nelson, the prevailing westerly winds may have contributed to greater oviposition on the more sheltered eastern side of the trees, and this may have occurred in other localities. Greatest damage on the eastern aspect of pear trees was also reported by Du et al. 40
Studies of the genetics and sibship of codling moth within orchards led Franck et al 68 to conclude that females usually cluster their egg deposition in the trees. This was also suggested by the distribution of eggs and larval entries within trees observed during the systematic sampling of the present research. The nearest-neighbor data provided only a first indication that there is probably significant aggregation of these codling moth stages within trees, but whether this is in addition to, or different from, the aggregation of fruits themselves within the trees remains to be investigated. The methodology of Everhart et al 66 appears suitable for such research. Aggregation is likely to be assisted by the known increase in the attractiveness of fruit to ovipositing females and neonate larvae caused by earlier codling moth infestation;31,32,72 on the other hand, aggregation may be reduced by the deterrence of oviposition caused by freshly laid eggs. 15
Distance of eggs from the nearest fruit
Geier, 4 Wearing et al, 16 Jackson, 1 and Blomefield et al 34 presented graphs showing the distribution of distances of codling moth eggs from their nearest fruit. When eggs laid directly on the fruit are included in these graphs (as in Fig. 8),1,16 it is clear that the proportion of eggs declines immediately away from the fruit, and then further with increasing distance, providing a good fit to a negative binomial distribution. 1 Measurements have shown that 90% of eggs are laid within 10 cm of the fruit, 4 91.3% within 20 cm, 1 or 96% within 12 cm. 34 Wearing et al 16 recorded that 99% of eggs were within 11 cm of ‘Delicious’ apples and that 47% were laid on the fruits themselves. The proportion of eggs laid on the fruit is usually less than that on the leaves, but the latter are spread over a radius of at least 24 cm, 4 17 cm, 16 or 16 cm. 34 Jackson 1 recorded eggs up to 50 cm from the fruit, which may be a result of the competition for fruit at a high mean density of 1.96 eggs/fruit cluster. Blomefield et al 34 noted that more eggs were laid on the fruit and on the leaves closer to ‘Granny Smith’ than ‘Golden Delicious’ fruit, which may reflect the much higher production of α-farnesene per unit area of fruit by the former cultivar. 67 But in all these cases, a very high proportion of eggs were laid within 15 cm of the fruit. The consistency of this behavior is further shown by the oviposition behavior of the females in fecundity cages, who laid their eggs at similar distances from the fruit to those under natural conditions, despite being confined (Fig. 8). In the total ‘Delicious’ cages of 1970/1971, 350 of the 495 apples and 4458 of the 4949 leaves were not used for oviposition.
Sutherland et al 7 pointed out that McIndoo73,74 and later Garlick (1948, cited in Ref. 75) were first to raise the possibility that neonate codling moth larvae moved toward apples in a directed manner involving a positive response to odor. Geier 4 observed the behavior of neonate larvae released onto fruit clusters at different distances from the fruit and was able to show that they located the fruit more efficiently than would occur at random. He also posited the presence of attractant odors from the fruit, which were later demonstrated,7,19,76 but several hours were frequently involved in fruit location. The preliminary data on Figure 9 suggest that there is an increased risk of the mortality of neonate larvae that hatch at a greater distance from the fruit, even in New Zealand where they have few natural enemies.37,77 In other locations where predators of neonates are important in population dynamics,36,78 major effects on mortality may be expected, particularly for eggs on the AbLS and wood. 79 In laboratory experiments, Sutherland et al 7 showed that neonate codling moth larvae in Petri dishes were attracted within five minutes to 100 ng of α-farnesene from a distance of at least 6.75 cm away, but the numbers of larvae reaching the source within that time had declined significantly compared with the larvae that began 5.00 cm away. In another laboratory experiment, Sutherland 19 showed that neonate larvae in almost still air oriented and moved within five minutes to a whole apple up to 1.5 cm away but not further. Given the greater distances, obstacles, air movement, and hazards in the field, it is not difficult to envisage some larvae requiring several hours to find the fruit, as observed by Geier. 4 Wei et al 43 reported that neonate larvae released on leaves up to 10 cm from the fruit were as successful at infesting the fruit (67.7%) as larvae released directly on the fruit (67.3%); moreover, this success rate was similar for larvae released on the AdLS or AbLS. However, it is unclear whether these experiments were conducted in the laboratory or field.
Spatial distribution–-site selection for oviposition
Variation in site selection for oviposition has been well documented. For instance, Jackson
1
recorded 57.3% of eggs on the AdLS versus 34.7% on the AbLS, with only 8% on the fruit. Blomefield et al
34
recorded 35.6% of eggs on the fruit of ‘Granny Smith’ compared with 10.8% on those of ‘Golden Delicious’, and of those eggs on the leaves of ‘Granny Smith’, 72.9% were on the AdLS, whereas only 30.2% were on the AdLS of ‘Golden Delicious’. Two major factors dominate the literature that analyzes oviposition site selection (fruit, AdLS, AbLS, and wood) by codling moth,
Wei et al 43 investigated whether the ovipositional preference for the AbLS on their experimental trees (‘Golden Delicious’ and ‘Red Delicious’) was a female preference for shade by turning the leaves over for presentation to ovipositing females. This procedure reduced oviposition on turned AbLSs compared with normal AbLSs, and increased oviposition on turned AdLSs compared with normal AdLSs. Wei et al 43 also reported higher hatch rate for eggs laid in the shade compared to those in the sun, and higher oviposition on the shady side of fruit clusters compared with the sunny side; they concluded overall that the shady versus sunny side of egg-laying surfaces was more important in the expression of preference than the physical properties of the surface. However, this fails to take account of the ovipositional preference of females for the AdLS of many apple cultivars. It seems probable that the expression of preference for egg laying in the shade is possible only on cultivars with low trichome density, whereas on cultivars with dense trichomes on the AbLS, the females lay preferentially on the AdLS.
Subinprasert and Svensson
79
reported that no eggs of codling moth were laid on the AbLS of apple leaves in a Swedish orchard. In addition, when eggs were artificially attached there, they suffered more predation than those placed on the AdLS or fruit. Greater numbers of insect predators are found on the AbLS,
79
and the mobility of neonate larvae of codling moth is reduced by the pubescence of the surface,
82
which could make them more susceptible to predation. Al Bitar et al
13
proposed that these factors, and reduced mobility of ovipositing females on the AbLS, caused a shift in their preference toward oviposition on the AdLS. They also noted the great ability of ovipositing females to walk on the smoother fruit and AdLS surfaces compared with predators and suggested that laying eggs directly on the fruit could be a mechanism to avoid predation. Laying on a variety of surfaces may be of similar benefit, as it is possible to envisage selection for a predator species well adapted to walking on the fruit surface if all eggs were laid there. In New Zealand, the dearth of predators of the eggs and neonate larvae of codling moth37,63,77 means that the preference of females to oviposit on the AbLS of some cultivars (eg, ‘Golden Delicious’, ‘Delicious’) is of little benefit to biological control. Moreover, there is little selection pressure from predation in New Zealand for ovipositing females to lay their eggs on sites other than the AbLS. It should also be noted that predatory species vary in their mobility and effectiveness as predators on different leaf surfaces; a predator of codling moth
The adhesion of eggs to the substrate is strongest on the fruit, then the AbLS, and the least on the AdLS;13,14 hence, the need for strongest adhesion does not determine the distribution of eggs. Specialized adaptation to make a strong bond between the egg and fruit 14 may be needed to counter the risk of dislodgement from this smooth surface that grows rapidly at times and is often exposed to leaf brushing (during wind) as well as inclement weather.
The position of codling moth eggs on the leaf has been little studied. Hagley et al
39
reported similar results for both the AdLS and AbLS on which 44%–48% were on the edge of the leaf, 35%–36% were interveinal, and only 17%–20% were near the midrib (
Spatial distribution between and within fruit clusters
Authors have consistently reported that codling moth oviposition per fruit cluster of different sizes does not fit a Poisson distribution because of greater egg laying on clusters with more fruit.1,4,34,37 This was also the result for eggs and neonate larval entries in the present research. When oviposition was expressed as the number of eggs per fruit, Geier 4 found that the distribution was random, with each fruit carrying an equal probability of egg deposition, albeit at very low density (0.09 eggs/apple). Results obtained by Wood 37 were similar when analyzing data from a low-density population (0.17 eggs/ apple), but not at higher densities (0.48 eggs/apple), reporting mild contagion of oviposition with too few instances of one egg laid per fruit cluster than would occur at random. Jackson, 1 working with a population at very high density, found even stronger evidence that egg distribution per fruit was aggregated, and he pointed out that low density can result in random distributions by default because of the low incidence of insects in the samples. At a mean density of 1.37 eggs/ apple, Jackson 1 obtained a curvilinear decrease in the oviposition per apple with increasing cluster size. The analyses of the current data are similar to those in the study by Wood. 37 A random distribution was obtained in 1967/1968 at a mean low density of 0.05–0.09 eggs/apple, whereas in 1972/1973 at a mean density of 0.31–0.60 eggs/fruit, the distribution was no longer random (Table 3), and unlike Jackson, 1 the eggs/fruit increased with increasing cluster size. This could be attributable to the effects of height within the tree on the size of clusters and the distribution of eggs, as suggested by Wood, 37 although no such height effect was reported by Jackson. 1
The foregoing may be a reasonable interpretation of the current oviposition results, but the larval entry data, which came from systematic sampling over four years, show that the sampling method also played a role. Neonate larval entries were random with respect to the fruit in each of those four years across a wide range of densities (Table 4), and nonrandom distribution occurred only when destructive sampling was used. By focusing this stratified random sampling on the known height distribution of eggs and entries, a bias to more heavily infested larger clusters in the upper stratum was probably introduced.
Similar considerations concern the distribution of eggs and entries
After comparing his results with those of Geier
4
and Wood,
37
Jackson
1
concluded that codling moth oviposition was not random with respect to fruit clusters or fruit and proposed a hypothesis of overlapping
There is additional evidence from the current research that the deterrent effect of freshly-laid eggs on further oviposition, as reported by Thiéry et al, 15 had little influence on the ovipositing females at the population densities being studied. This comes from the analysis of the number of eggs laid directly on the fruit. In 1972/1973, when the sampling obtained 1.22 and 0.78 eggs/fruit cluster on the unsprayed ‘Delicious’ and ‘Red Delicious’, respectively, the number of eggs laid directly on the fruit fitted a random distribution, with no evidence of dispersive behavior. Aggregation was found in the other cultivar blocks, but only in the presence of ryania deposits from spraying; this may be attributable to the known repellency of ryania deposits to the larvae, 63 potentially leading to aggregation of larval entries on fruits with poorer spray coverage. Highly aggregated oviposition on both fruit and leaves occurred when moths were confined in fecundity cages, such as in 1970/1971 (Table 7) at mean densities of about 1.5 eggs/fruit and 0.4 eggs/leaf. Under these conditions, females laid up to 26 and 32 eggs on a single fruit and leaf, respectively; and only 37–38% of fruit and 14% of leaves were used for oviposition. Jackson 1 similarly reported that eggs laid per leaf did not fit a Poisson distribution under natural conditions (too many leaves without eggs and too many with >1 egg) where the mean density was 1.37 eggs/fruit, and Wei et al 41 reported that aggregation of codling moth eggs increased with population density.
Subinprasert and Svensson 79 investigated the survival of codling moth eggs attached to leaves in the field at different densities and in different configurations of the same density and showed that lowest predation occurred with one egg per leaf, and was generally associated with eggs placed in the most widely dispersed configuration. They commented that the strategy of laying eggs singly also reduced larval competition for fruit and hence potential cannibalism, which added to the benefits of laying on the AdLS or fruit and the associated low predation rates mentioned earlier. They also noted that the searching costs for the female of laying eggs singly in an orchard were not high because of the abundance of fruit in close proximity to each other. Similar conditions may have occurred in the area of origin of codling moth in Eurasia, such as the apple forests of Kazakhstan. 86 Even outside orchards, the sedentary behavior of most females is such that their searching activity is largely confined to one or two neighboring trees. 68 Subinprasert and Svensson 79 concluded that laying eggs singly was the most advantageous strategy for codling moth egg and larval survival.
Conclusions and their implications for codling moth management
The research described in this study was conducted about 40 years ago, but it remains as relevant today as when the work was done. The results reinforce the findings of earlier and more recent research and add a number of new findings. These include the extremely low fecundity of wild gravid female codling moth (2.5 eggs laid/female) caged in the field on apple branches without fruit; the limitation of egg numbers laid directly on the fruit, despite increasing egg deposition on leaves and stems nearby; the uniformity of the distribution of eggs by distance from the nearest fruit, even when females were confined in cages; the increased mortality of neonate larvae that hatched more distant from the fruit; exponential increase in eggs laid per fruit cluster as cluster size increased; sampling method effects on egg distribution analysis; aggregated distribution of neonate larval entries on individual fruits; and aggregation of neonate larval fruit entries in apple trees as shown by the nearest-neighbor analysis.
While tree training, tree height, and apple cultivars vary between apple-growing regions and over time, integration of the current research data with a literature review has enabled reconciliation of differing published reports of codling moth distribution. For example, population density differences account for much of the conflicting evidence of egg and larval distributions within and between fruit clusters. And leaf pubescence can explain most of the differences between reported egg distributions on the AbLS and AdLS of apple cultivars.
Overall, this analysis has revealed the underlying characteristics of codling moth behavior and host plant structure/physiology that together determine egg and larval distribution in its host trees. The adult and larval response to fruit is of overriding importance and results in similar fruit-centric spatial distribution on a wide range of cultivars and in many locations (Fig. 8). Higher cropping trees of all cultivars attract greater numbers of codling moths that are likely to be most active in the upper canopy and in the sunnier (north or south, depending on hemisphere) and eastern aspects of the trees. Larger fruit clusters attract even greater oviposition and larval attack than can be explained by greater fruit number alone, particularly in the upper canopy and in high-density codling moth populations. Within fruit clusters, certain fruits are especially vulnerable for reasons that have yet to be delineated but are likely to include more advanced development and greater production of attractant volatiles. Thus, egg and larval aggregation are characteristics of codling moth distribution both within and between trees, within and between fruit clusters, and within fruit.
Yet despite the dominant importance of the fruit, codling moth has an oviposition strategy that limits the proportion of eggs laid on the fruit, with >50% being on leaves close by for almost all cultivars. Within the distribution on the leaves, trichome density has a primary influence in determining the proportion of eggs laid on the AdLS and AbLS of cultivars, both old and new. Less pubescent cultivars are invariably associated with more eggs on the AbLS, favored by some insect predators, while the reverse is true for hairier cultivars. The current research has provided first field evidence that eggs laid on leaves more distant from the fruit suffer greater mortality, be this from natural enemies or desiccation before fruit entry.
Codling moth management can exploit this knowledge of egg and larval distribution in a number of ways. Pruning and thinning practices can be used to obtain predominantly 1- or 2-fruit clusters that attract less egg laying than larger clusters and enable better spray coverage with ovicidal chemicals. 34 Even in the absence of such sprays, preliminary observations in the current research recorded the highest mortality of neonate larvae that hatched on 1-fruit clusters (46.4%) compared with zero mortality of those hatching on 4-fruit clusters, and intermediate mortality on cluster sizes in between. Hand thinning is used by many organic growers, and removal of codling moth-damaged fruit should pay particular attention to large fruit clusters, groups of fruit clusters, and the upper areas of the trees.
Less hairy apple cultivars that permit greater oviposition on the AbLS (eg, ‘Golden Delicious’, and ‘Delicious’) are likely to retain the preference of predatory insects to feed there, and this could result in greater predation of eggs and larvae. These cultivars have the further advantage of permitting better ovicidal or larvicidal spray coverage than those with dense trichomes. Blomefield et al 34 suggested that, for all cultivars but especially those with more pubescent leaves, surfactants could be important in improving ovicidal efficacy when spraying. The importance of the effect of the surface of the fruit, AdLS, and AbLS on the ability of female codling moths to walk and oviposit13,14 suggests that compounds may be found that interfere with this process, as a control option. In the current research, ryania was found to affect egg and larval distribution, and other insecticides may have such effects.
This review has shown that spray machinery should be configured and calibrated to ensure that, in most apple-growing regions, applications against codling moth target the mid-upper strata of the trees. The results of Blomefield et al 34 showed that this could not be recommended universally. They found that in South Africa the height distribution of eggs changed from being primarily in lower strata early in the season to upper strata later and concluded that spraying practices should change accordingly. But this is exceptional in the literature, and greater codling moth activity in the upper areas of apple trees is still widespread and already affecting the application of pheromone for mating disruption and trapping of this pest.60,61 The trend to the increased planting of dwarf apple trees, especially in Europe, is reducing the significance of tree height in codling moth distribution. This includes the use of centrifugal training that results in higher codling moth damage in the inner areas of the trees than that occurring in more compact tree forms. 62
Cultivars have been shown by this review to have a wide variety of effects on codling moth and its distribution. There is recent renewed interest in the breeding and selection of apple cultivars with reduced susceptibility and/or antibiotic resistance to codling moth.84,87,88 Research has shown that cultivars or
Author Contributions
Conceived and designed the experiments: CHW. Analyzed the data: CHW. Wrote the first draft of the manuscript: CHW. Developed the structure and arguments for the paper: CHW. Made critical revisions and approved final version: CHW. Author reviewed and approved of the final manuscript.
Footnotes
Acknowledgments
The author thanks Mr. L. Skilling for technical assistance, Mr. M. van Geldermalsen and his staff at Appleby Research Orchard, Nelson, for orchard management, and Dr. H. R. Thompson (Applied Mathematics Division, Department of Scientific and Industrial Research) for providing the χ2 homogeneity test used for Tables 5 and ![]() . I thank three anonymous reviewers for their constructive criticism of an earlier version of this article.
. I thank three anonymous reviewers for their constructive criticism of an earlier version of this article.