
Abstract
Among 36 differentially-expressed genes during growth in longissimus muscle (LM) of Angus steers, Yin Yang 1 (YY1) had the most relationships with other genes including some associated with adipocyte differentiation. The objective of this study was to examine the effect of nutritional management on mRNA expression of YY1 along with its targets genes PPARG, GTF2B, KAT2B, IGFBP5 and STAT5B. Longissimus from Angus and Angus × Simmental steers (7 total/treatment) on early weaning plus high-starch (EWS), normal weaning plus starch creep feeding (NWS), or normal weaning without starch creep feeding (NWN) was biopsied at 0, 96, and 240 days on treatments. Results suggest that YY1 does not exert control of adipogenesis in LM, and its expression is not sensitive to weaning age. Among the YY1-related genes, EWS led to greater IGFBP5 during growing and finishing phases. Pro-adipogenic transcriptional regulation was detected in EWS due to greater PPARG and VDR at 96 and 240 d vs. 0 d. GTF2B and KAT2B expression was lower in response to NWS and EWS than NWN, and was most pronounced at 240 d. The increase in PPARG and GTF2B expression between 96 and 240 d underscored the existence of a molecular programming mechanism that was sensitive to age and dietary starch. Such response partly explains the greater carcass fat deposition observed in response to NWS.
Background
Recent transcriptomics and bioinformatics research by our group revealed that, among 36 transcription regulators differentially-expressed during growth in longissimus muscle (LM) of early-weaned Angus steers, Yin Yang 1 (YY1) had the most relationships with other differentially expressed genes (with a total of 5,616). 1 In non-ruminants, YY1 is a ubiquitously-expressed DNA-binding zinc-finger transcription factor implicated in the regulation of genes that are important for cell growth, development, differentiation, cell cycle, and programmed cell death. 2
Expression of YY1 in bovine tissue appears to be greater in fat cells of developing tissue as demonstrated by data from mammary fat pad vs. mammary epithelium. 3 Because the differentiation of 3T3-L1 preadipocytes to adipocytes is dependent on the suppression of the expression of CHOP-10 (a member of the C/EBP family) by YY1, 4 this transcription factor also could play a role in intramuscular fat (IMF) deposition of young beef cattle. Bioinformatics analysis using Ingenuity Pathway Analysis® (IPA; Ingenuity systems, CA, USA) revealed that YY1 interacts with insulin-like growth factor binding protein 5 (IGFBP5), signal transducer and activator of transcription 5B (STAT5B), general transcription factor 2B (GTF2B), and K(lysine) acetyltransferase 2B (KAT2B). The latter two play a role in chromatin remodeling (SWItch/Sucrose Non-Fermentable (SWI/SNF) and CBP/p300) and gene transcription. 5
Previous work has demonstrated that weaning beef calves earlier than the common 205 days of age and feeding high dietary starch, compared with high dietary fiber, leads to marked upregulation of adipogenic and lipogenic genes in LM. 6 Such nutritional management strategy often enhances carcass fat deposition.7,8 However, it remains unknown if expression of YY1 and its known interacting partners is responsive to nutritional management or whether it is associated with carcass fat deposition.
The general hypothesis of the present study was that longitudinal expression of YY1, its closely-related genes, and PPARG within LM would be altered by nutritional management in LM of growing steers. As an initial step in establishing a role for YY1 in LM, the specific objective was to profile gene expression of LM tissue from steers managed to consume different levels of dietary energy at an earlier or normal weaning age. Those data were combined with concentrations of hormones in blood as a way to better understand the system in a more holistic fashion.
Methods
Animal management
All experimental procedures involving steers were approved by the University of Illinois Institutional Animal Care and Use Committee under protocol no. 09143. Angus (A) and Angus × Simmental (SA) steer calves from the University of Illinois beef cattle herd were utilized. Prior to the start of the study, 12 A and 9 SA steers were randomly assigned to early weaning (129 ± 36 days of age) plus a high-starch diet (EWS, 4 A and 3 SA), normal weaning (225 ± 36 days of age) plus corn-based creep supplement (NWS, 4 A and 3 SA), and normal weaning with no creep feeding (NWN, 4 A and 3 S A). The latter reflects closely the current trends in management in the US industry. Steers in NWS had access to creep feeders in the pasture at time of early weaning. The EWS steers were fed ad libitum during their growing phase and, when NWS and NWN steers joined the feedlot with the EWS, all steers received the same finishing diet ad libitum. Table 1 reports the composition of the diet and creep supplement. The vitamin fraction of the dietary supplement contained 680,329 IU vitamin A/kg of dry matter, 68,039 IU vitamin D3/kg of dry matter, and 9,072 IU vitamin E/kg of dry matter.
Composition of the early wean high-starch diet (EWS), the corn-grain-based creep supplement fed to normal-wean steers (NWS), and the finishing diet offered to all steers the day after until harvest (finishing phase).

Composition of growing phase supplement (g/kg DM): Soybean Meal (90.2), Limestone (90), Trace Mineralized Salt (6.7), Copper Sulfate (0.03), Vitamin ADE (0.7);
composition of finishing phase supplement (g/kg DM): Ground Corn (844), Limestone (110), Urea (25), Trace Mineralized Salt (1), Thiamine (2.5), Copper Sulfate (0.5) and Vitamin ADE (1.0).
For the 96 d treatment period (ie, growing phase) EWS calves remained in the feedlot and NWS and NWN dams nursed their calves and grazed mixed pastures of endophyte-infected tall fescue (Festuca arundinacea), orchardgrass (Dactylis glomerata), bluegrass (Poa pratensis), red clover (Trifolium pratense), white clover (Trifolium repens), and alfalfa (Medicago sativa). Calves in NWS and NWN were weaned at 225 days of age and joined the early-weaned calves at the feedlot at which point both groups were fed a common finishing diet until slaughter (Table 1). Seven animals within EWS (n
Yin Yang 1 gene network expression
An investigation using the 126 genes linked to YY1 in LM from our previous microarray study 1 was used to assess which genes could be related to a biological process associated with myogenesis and adipogenesis. The relative level of expression of YY1 and its target genes in adipose tissue was the first parameter that we had to corroborate during the literature search; once the expression level of a gene and the possible role in adipose tissue was confirmed, it was chosen for qPCR in LM. The selection based on literature searches resulted in the identification of IGFBP5, STAT5B, GTF2B, and KAT2B. These 4 genes act as links between YY1 and RXR β (RXRB), PPARG, and VDR.
Primer design and evaluation
Primers were designed using Primer Express 3.0 with minimum amplicon size of 100 bp and limited 3′ G+C (Applied Biosystems, CA). When possible, primers were designed to fall across exon–exon junctions. Primers were aligned against publicly available databases using BLASTN at NCBI and UCSC's Bos taurus Genome Browser Gateway. Prior to qPCR, primers were tested in a 20 μL PCR reaction using the same protocol described for qPCR except for the final dissociation protocol. For primer testing we used a universal reference cDNA (RNA mixture from 5 different bovine tissues) to ensure identification of desired genes. Five μL of the PCR product were run in a 2% agarose gel stained with ethidium bromide (2 μL). The remaining 15 μL were cleaned using QIAquick® PCR Purification Kit (QIAGEN) and sequenced at the Core DNA Sequencing Facility of the Roy J. Carver Biotechnology Center at the University of Illinois, Urbana-Champaign. Only those primers that did not present primer-dimer, had a single band at the expected size in the gel, and had the right amplification product (verified by sequencing) were used for qPCR. The accuracy of a primer pair also was evaluated by the presence of a unique peak during the dissociation step at the end of qPCR. Additional information is available in Tables 2 and 3.
Gene ID, GenBank accession number, sequence and amplicon size of primers used to analyze gene expression by qPCR.

Primer direction (F–-forward; R–-reverse) and hybridization position on the sequence;
amplicon size in base pairs (bp).
Sequencing results of PCR products from primers of genes designed for this experiment. Best hits using BLASTN (http://www.ncbi.nlm.nih.gov) are shown.

Biopsy, RNA extraction and PCR
Details of all these procedures have been reported previously. 6 Briefly, a needle biopsy gun fitted with a 12 gauge biopsy needle was used to harvest ~0.5 g of tissue (Bard Magnum, C. R. Bard, Covington, GA, USA). Trizol extraction of RNA was exactly as reported previously. 11 qPCR data were calculated with the 7900 HT Sequence Detection Systems Software (version 2.2.1, Applied Biosystems, CA). The final data were normalized using the geometric mean of the genes UXT, MTG1 and RPS15A, which were previously identified as suitable internal control genes in LM 11. Additional details of qPCR performance are in the Table 4.
qPCR Performance.
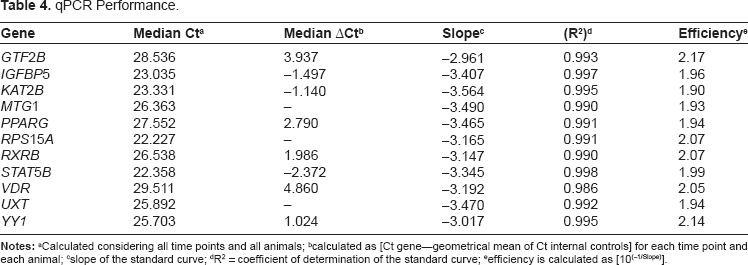
Calculated considering all time points and all animals;
calculated as [Ct gene–-geometrical mean of Ct internal controls] for each time point and each animal;
slope of the standard curve;
R 2 = coefficient of determination of the standard curve;
efficiency is calculated as [10(–1/Slope)].
Statistical analysis
Quantitative PCR data were analyzed using the MIXED procedure of SAS (SAS 9.1 Institute, Cary, NC, USA). Before statistical analysis, normalized qPCR data were transformed to fold-change relative to day 0 (ie, early weaning day). To estimate standard errors at day 0 and prevent biases in statistical analysis, normalized qPCR data were transformed to obtain a perfect mean of 1.0 at day 0, leaving the proportional difference between the biological replicate. The same proportional change was calculated at all other time points to obtain a fold-change relative to day 0. Fixed effects in the statistical model for each gene and blood parameters analyzed included treatment, breed, time, treatment × breed, treatment × time, and breed × treatment × time. Gene and blood data analysis of the three time points studied included a repeated-measures statement with an autoregressive covariate structure. Age at weaning was used as a covariate in the model. The random effect in all models was steer within treatment. The statistical model used was:
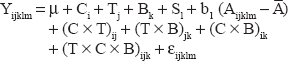
Moreover, partial Pearson correlation analysis among genes, adjusted for the fixed effects, was conducted using PROC CORR in SAS (Table 6).
Serum concentration (ng/mL) of leptin, growth hormone (GH), and IGF-1 of steers (n = 7/treatment) managed under three nutritional treatments during the growing phase (0 to 96 d) followed by feeding a common high-starch diet during the finishing phase (97 to 240 d).
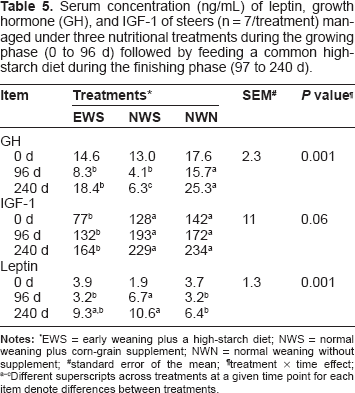
EWS = early weaning plus a high-starch diet; NWS = normal weaning plus corn-grain supplement; NWN = normal weaning without supplement;
standard error of the mean;
treatment × time effect;
Different superscripts across treatments at a given time point for each item denote differences between treatments.
Carcass quality parameters of Angus (A) and Angus × Simmental (SA) steers (n = 7/treatment) managed under three nutritional treatments during the growing phase (0 to 96 d) followed by feeding a common high-starch diet during the finishing phase (97 to 240 d).
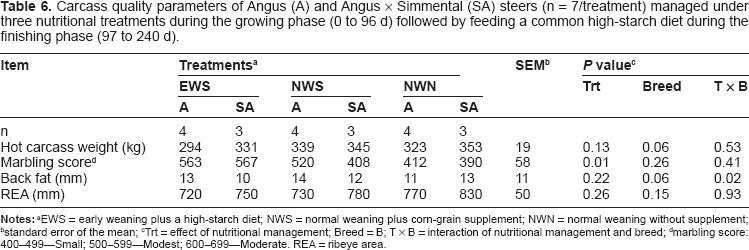
EWS = early weaning plus a high-starch diet; NWS = normal weaning plus corn-grain supplement; NWN = normal weaning without supplement;
standard error of the mean;
Trt = effect of nutritional management; Breed = B; T × B = interaction of nutritional management and breed;
marbling score: 400–-499–-Small; 500–-599–-Modest; 600–-699–-Moderate. REA = ribeye area.
Results
Blood metabolites
The concentration of GH was lower (interaction P < 0.05) in both EWS and NWS than NWN at 96 d (Table 5). However, IGF-1 concentrations were lower in EWS than in NWS and NWN steers at 96 days. Leptin concentrations were greater (interaction P < 0.06) in NWS than EWS and NWN at 96 d. At 240 d, concentrations of GH and leptin in EWS steers were intermediate to those in NWN (P < 0.05) and NWS (P < 0.05). However, EWS steers had lower IGF-1 at 240 d than in NWN and NWS. For these three metabolites there was no significant breed effect.
Carcass traits
There was no difference in HCW between EWS, NWS, and NWN steers. Even though carcass marbling score did not have a significant treatment x breed interaction (P = 0.41; Table 6), it had a significant treatment effect (P = 0.01), with greater values in EWS steers compared with NWS and NWN steers. This result led to a greater percentage of steers grading Choice or greater (data not shown). Back fat had a treatment × breed interaction (P = 0.02) with a breed effect (P = 0.06) due to greater values with A steers in EWS and NWS, but greater values with SA steers in NWN.
Gene expression
Results for YY1 indicated that there was no time x treatment effect (P = 0.34) but there was an overall effect of time (P < 0.05) on mRNA expression of all genes, ie, expression of YY1 increased throughout the study with highest expression at 240 d (Fig. 1). Among genes directly linked with YY1, there was a treatment x time interaction (P = 0.06) for RXRB due to greater expression for NWN compared with EWS at 240 days, and a strong negative correlation with YY1 and a positive correlation with VDR, IGFBP5 and STAT5B (Table 6).
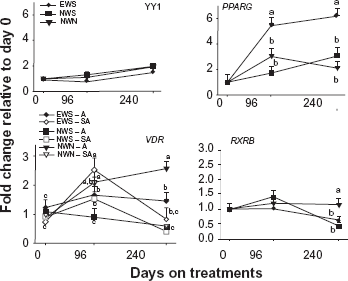
Patterns of mRNA expression of Ying Yang 1 (YY1), peroxisome proliferator-activated receptor γ (PPARG), vitamin D receptor (VDR), and retinoic X receptor β (RXRB).
The VDR gene was the only gene with a significant (P = 0.0006) breed × treatment × time interaction due to a gradual increase in expression during the growing phase with both EWS and NWN vs. NWS. Expression of VDR was lower during the finishing phase for both EWS and NWS but remained unchanged with NWN steers. Overall expression of VDR at 240 days was highest for NWN, intermediate with EWS, and lowest with NWS. There was a strong negative correlation between VDR, IGFBP5, RXRB, and STAT5B with YY1 in animals in NWS (P < 0.01), with RXRB in EWS (P = 0.02), and with STAT5B in NWN (P = 0.05); whereas, there was a positive correlation between YY1 with KAT2B (P = 0.001) and GTF2B (P = 0.001) in NWN, with IGFBP5 (P = 0.01) and STAT5B (P = 0.004) in NWS, and with GTF2B (P = 0.001) and KAT2B (P = 0.05) in EWS (Table 6).
There was a treatment × time interaction (P < 0.01) for the expression of PPARG due to a gradual increase between 0 and 96 d at which point expression was highest with EWS compared with NWS and NWN. During the finishing phase, expression of PPARG remained higher with EWS and also NWS, while it decreased due to NWN. Thus, at 240 days the expression of PPARG was markedly higher for EWS and not different for NWS and NWN (P < 0.01). In the EWS group, there was a positive correlation with IGFBP5 and KAT2B (Table 7). However, in both NWS and NWN there was a strong negative correlation between PPARG with STAT5B and in NWS only, between YY1, GTF2B and KAT2B (Table 7).
Pearson correlation coefficients for genes belong to YY1 network adjusted for the fixed effects.
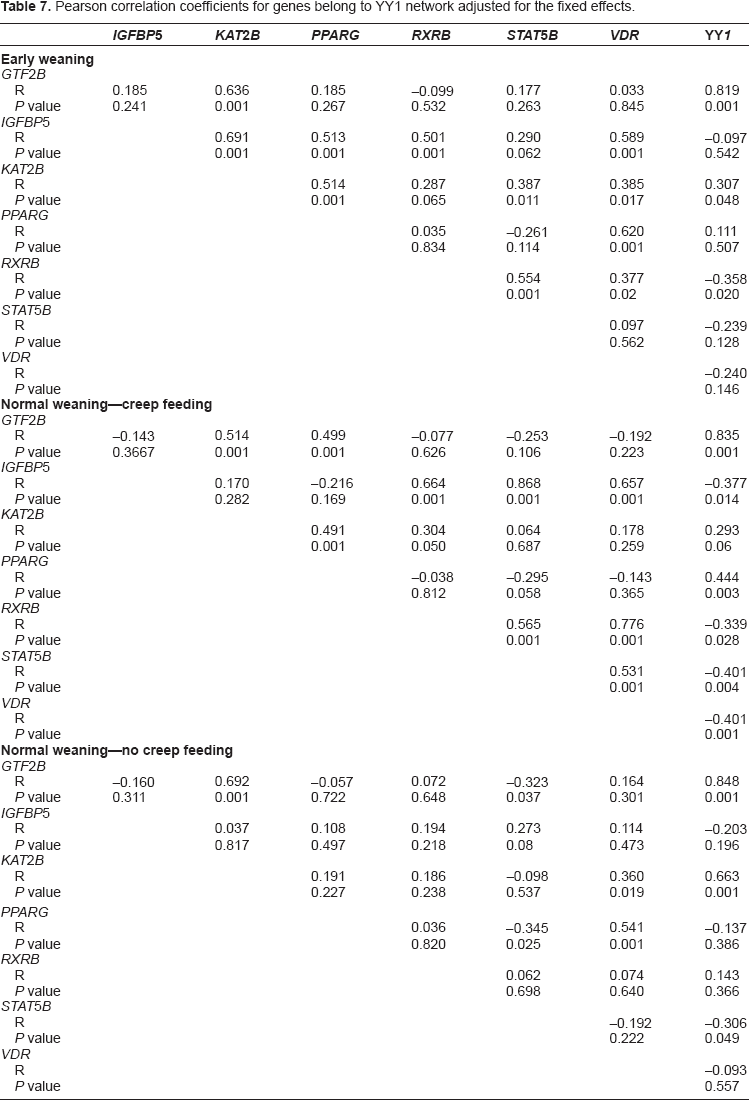
There was a time × treatment interaction (P < 0.01) and a breed × treatment interaction (P = 0.04) for IGFBP5. The former was due mainly to a longitudinal increase in expression with EWS but a decrease with both NWS and NWN leading to greater expression with EWS at 96 and 240 d (Fig. 2). In the NWS group, there was a negative correlation between YY1 and IGFBP5 and a strong positive correlation between YY1 and STAT5B. In the EWS group, there was a positive correlation between IGFBP5 with KAT2B and also between IGFBP5 and STAT5B (Table 7).
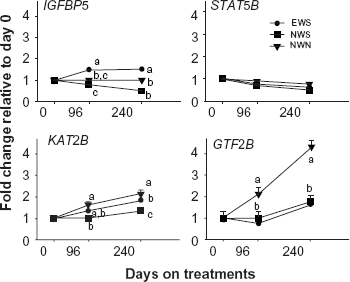
Patterns of mRNA expression of insulin-like growth factor binding protein 5 (IGFBP5), signal transducer and activator of transcription 5B (STAT5B), K(lysine) acetyltransferase 2B (KAT2B), and general transcription factor 2B (GTF2B).
In the case of GTF2B, there was a time x treatment interaction (P < 0.01) due to a gradual increase in expression primarily with NWN compared with EWS and NWS. Both of those treatments had the same response during the growing and the finishing periods resulting in greater expression at 96 and 240 days than at 0 d, however it was never higher than for NWN (Fig. 2). In all treatments there was a marked positive correlation between GTF2B with YY1 and between GTF2B and KAT2B. The correlation between GTF2B and PPARG was positive in the NWS group. However, a negative correlation was found between GTF2B and STAT5B in the NWN group (Table 7).
Expression of KAT2B had a time x treatment interaction (P = 0.02) due to a longitudinal increase in expression that was lower with NWS at 96 d, and with both NWS and EWS than NWN at 240 d. Moreover, KAT2B was the only gene from the ones related to YY1 with an effect of age at weaning (P < 0.01, data not shown). During the finishing period, all treatments had greater KAT2B expression relative to 0 and 96 d, however at 240 d expression was highest with NWN, intermediate with EWS, and lowest with NWS (Fig. 2). In the EWS group, there was a positive correlation between KAT2B expression and that of GTF2B, IGFBP5, PPARG, STAT5B, VDR, and YY1. In the NWS group there was a positive correlation between KAT2B expression and that of GTF2B, PPARG, and RXRB. In the NWN group there was a positive correlation between KAT2B with GTF2B, VDR, and YY1 (Table 7). Figure 3 depicts a summary of the relationships among genes and the overall response between treatments over time.
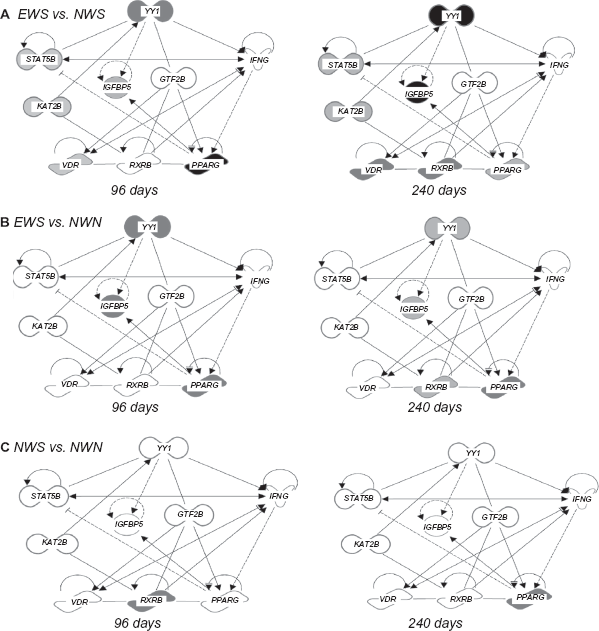
YY1 and target gene networks generated using ingenuity Pathway Analysis®.
Discussion
Nutritional management and carcass traits
Greater marbling scores in EWS steers, leading to a greater percentage of steers grading Choice or greater (data not shown), were likely due to the early feeding of starch. 6 This is expected to enhance ruminal propionate production with a resulting increase in serum insulin and glucose concentrations. 12 As propionate is the major glucogenic volatile fatty acid, more glucose is generated in the liver, resulting in more net energy available to the animal. A pro-insulinemic response would have led to greater glucose uptake by adipose. Despite the similar amount of time required for all steers to reach the point of harvest, EWS steers had longer access to a diet that is more typical of the finishing phase, ie, the molecular mechanisms driving marbling deposition were likely to have been activated earlier than normal, as we have previously seen. 6 These responses to high-starch feeding were accompanied by greater concentrations of leptin in serum. This response could partly explain the greater marbling in EWS steers during the finishing phase as serum leptin levels are positively correlated with percentage of body fat and fat mass. Steers in these groups likely had larger adipocytes, which actively secrete leptin. 13
Early studies led to the suggestion that calves must be creep-fed for ca. 80 days to express their maximum potential for marbling deposition. 8 However, despite NWS steers receiving creep feed for a period of 96 days, that in itself did not seem to affect the molecular mechanisms enhancing marbling relative to steers which received milk and pasture as their sole source of feed (NWN). Our results underscored the potential for precocious initiation of marbling in young steers when fed higher dietary starch.
Yin Yang 1 network gene expression
Although PPARG is not a direct target of YY1 (Fig. 3), YY1 binds and represses PPARD leading to inhibition of ligand-induced transcription activity. 14 Thus it is possible that YY1 in bovine skeletal muscle exerts some degree of inhibition over PPARG, which seems to be supported by the contrasting response in expression of PPARG and YY1 specifically in the EWS group (Figs. 2 and 3). Those animals had greater PPARG expression, supporting the existence of a more robust adipogenic/lipogenic activity.
The bioinformatics analysis using IPA of microarray data from LM in our previous study 1 revealed that the complex formed between VDR and RXR is affected by YY1. Transcription Factor 2 B (TFIIB) and CREB binding protein (CBP) bind to YY1 and regulate activation of transcription via 1,25-(OH)2D3 through an alleviation of the repressive effect induced upon YY1 binding to VDR. 15 YY1 also competes for the vitamin D response element (VDRE) with TFIIB through protein-protein interactions. 16 Hence it has been speculated that maximizing suppression of vitamin D induction when a target gene is not transcriptionally active may enhance affinity of YY1 for VDRE. 17 The outcome of this suppression would be the maintenance of basal transcription and inhibition of vitamin D sensitivity.
In NIH3T3 adipocytes, there is a stimulation of YY1 expression, which is dependent on serum containing IGF-1. Moreover, after prolonged serum deprivation, NIH3T3 cells lost YY1 expression. The rapid change in YY1 expression due to growth factor deprivation suggested the possibility that YY1 mediates some of the intracellular responses to IGF-1. 18 This fact is important in the context of our study as it was previously shown that serum IGF-1 concentration in Angus steers had a negative correlation with greater marbling scores, quality grades, fat thickness, and yield grades. 19 Most IGF-1 in the circulation is bound to IGFBP, which can either inhibit or facilitate IGF-1 binding to the IGF-1 receptor. IGFBP5 is a YY1 target gene in mouse embryonic fibroblasts (MEF) and it is expressed in white adipose tissue, kidney, and heart. 20 The pro-adipogenic role of IGFBP5 in LM tissue is likely indirect (eg, Fig. 3) and via several VDRE and PPARG response elements (PPRE) previously discovered in IGFBP5 promoter regions. 21
IGF-1 is produced primarily by liver as an endocrine hormone stimulated by GH, and its effect can be delayed by undernutrition, GH insensitivity, lack of GH receptors, or failure of the GH receptor downstream signaling pathway that employs STAT5B. 22 Work in non-ruminants showed that YY1 is a component of the GH-inducible nuclear factor complex (GHINF), which exerts close regulation of STAT5B transcription. 23 Since the distribution and function of each STAT (STAT 1, 2, 3, 4, 5A, 5B and 6) is unique, the regulation of tissue specific genes may reflect a physiological role for these proteins. 24 In adipose tissue, STAT5B activation is highly-specific and, upon activation, only GH was able to induce STAT5B to translocate to the nucleus. 25 Interestingly, YY1 associates with STAT5B during the response to GH in adipose tissue. 23 GH-activated STAT5B can inhibit PPAR-regulated transcription. This inhibitory cross-talk is mutual, ie, GH-induced-STAT5B can be inhibited by ligand-activated PPARG. 26
In the current study, the antagonistic transcriptional role of GH on PPARG has to be discarded as a central regulatory mechanism associated with marbling mainly because of a lack of treatment differences on STAT5B expression. Although it has been hypothesized that STATs may have a regulatory function in adipocyte gene expression, their particular functions during adipogenesis or in the mature adipocyte remain mostly unknown. Results showed a modest but consistent downregulation of STAT5B expression during development, which may be partly explained by the upregulation (ie, stimulation) of PPARG. Therefore, the lack of treatment effect on STAT5B expression argues against a possible GH-mediated inhibition of PPARG activation by STAT5B which was reported previously in rodents. 26 Such a mechanism was most evident in the EWS group.
GTF2B/TFIIB (General transcription factor IIB) is one of the ubiquitous transcription factors required for transcription initiation [ie, formation of the pre-initiation complex (PIC)] by RNA polymerase II (RNAP II). The rate of RNAP II-directed transcription is affected by VDR. 27 YY1 represses VDR-mediated 25-Hydroxy-vitaminD3-24-hydroxylase transcription by sequestering TFIIB/CBP. In addition, the N-terminal region of cAMP response element-binding protein (CBP) that interacts with YY1 can inhibit YY1 from binding to TFIIB. Thus, CBP may diminish YY1-mediated repression by preventing YY1 from binding to TFIIB, which is required for VDR-mediated transcription. 15 Besides its role as lipogenic substrate, acetate availability in intramuscular adipocytes within LM of growing steers also could alter the function of GTF2B, which is an acetyltransferase with a key role in promoting gene transcription. 28
One likely explanation for the greater GTF2B expression due to NWN is the higher availability of acetate (and acetyl-CoA) as substrate for the acetyltransferase reaction. That is, the pasture intake enhanced the production of ruminal acetate and could have enhanced transacetylation, thus, facilitating the interaction of GTF2B with other members of the PIC complex and SWI/SNF complex. 29 In that context, our data underscored that lipogenic substrate availability per se (ie, greater acetate) is insufficient to promote adipogenesis. In NWN steers, it is also possible that although the PIC complex was fully formed it remained “inactive” perhaps due to lower glucose availability (from gluconeogenesis) for ATP generation compared with EWS and NWS. The end result of such adaptations would have been a “repression” of the transcription machinery.
The transcriptional repressor activity of YY1 is regulated through histone acetylation by the transcription coactivators p300 (KAT3B) and PCAF (KAT2B). 30 PCAF-mediated acetylation supports SWI/SNF recruitment to the PPARG promoter 31 and functions as a nuclear receptor coactivator for retinoic acid receptor (RAR). Both KAT2B and GTF2B, encoding histone transacetylase, had a similar expression pattern in NWN steers likely due to greater availability of acetate and acetyl-CoA. However, as indicated above this response was insufficient to promote a proadipogenic response.
Summary Model
Figure 4 depicts a model based on data from the present study and data from published literature cited in the manuscript. The YY1-related network is represented by the genes encoding IGFBP5, STAT5B, GTF2B, and KAT2B. The transcription factor YY1 is part of the GHINF complex in which the major DNA binding component is STAT5B. Signaling via GH induces tyrosine phosphorylation, nuclear localization, and DNA binding of STAT5B. The GH-induced STAT5B signaling in primary preadipocytes represses transcription of genes required for terminal differentiation, partly by repressing PPARG. The GTF2B/TFIIB is one of the ubiquitous factors required for transcription initiation by RNA polymerase II. The interaction of the VDR with TFIIB represents a potential physical connection between the VDR-DNA complex and the transcription PIC. YY1 represses VDR-mediated 25-Hydroxy-vitaminD3-24-hydroxylase transcription by sequestering TFIIB/CBP.
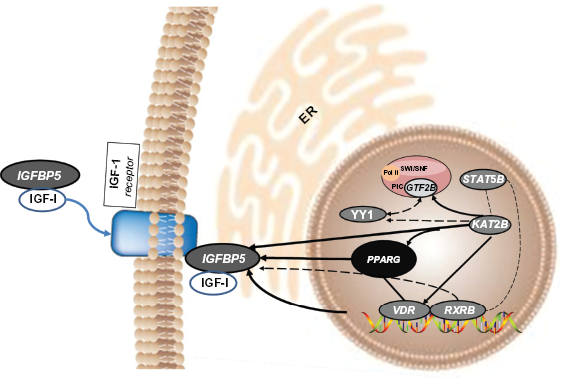
Summary of YY1 gene network expression in steers comparing early weaning (EWS) against normal weaning without creep feeding (NWN) at 240 days on treatment.
The transcriptional PIC interacts with transcriptional co-activators via protein-protein interactions. These co-activators are an ATP-dependent chromatin remodeling complex called SWI/SNF that leads to local chromatin de-condensation and gene expression. The SWI/SNF complex has a role as a bromodomain-containing transcriptional co-activator/co-repressor in adipogenesis. The KAT2B protein has a role in ensuring the maintenance of a stable association of the complex with chromatin, hence, maintaining the PIC on the PPARG promoter to support transcription. YY1 corepressor activity is regulated through histone acetylation by PCAF (KAT2B). Acetylation of the central region of YY1 is required for a full transcriptional repressor activity, while acetylation of the C-terminal zinc finger domain decreased the DNA-binding activity of YY1.
Conclusions
Based on upregulation of PPARG, early exposure to a diet with a high-starch level appeared to induce precocious intramuscular adipocyte differentiation that led to greater marbling score in the carcass. As the response did not seem to involve YY1, its role in this process is likely minor. During the growing phase, diets that increase ruminal acetate to propionate ratio (NWN) seemed to induce formation of a transcriptional complex, suggesting that transacetylation of GTF2B is responsive to substrate availability. The availability of adequate propionate for gluconeogenesis and subsequent provision of glucose to LM also seems important in order to activate the transcriptional machinery.
Funding
Author(s) disclose no funding sources.
Author Contributions
Conceived and designed the experiments: JJL, DWS, DBF. Conceived and performed the analyses: SJM, WTM, DK. Wrote the manuscript: SJM, JJL. Agree with manuscript results and conclusions: SJM, DWS, WTM, DK, DBF, JJL. Jointly developed the structure and arguments for the paper: SJM, JJL. Made critical revisions and approved final version: SJM, DWS, JJL. All authors reviewed and approved of the final manuscript.
Competing Interests
Author(s) disclose no potential conflict of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was sub.ect to blind, independent, expert peer review. The reviewers reported no competing interests.
Footnotes
Acknowledgements
We gratefully acknowledge the help from T. G. Nash and J. Dahlquist as well as the rest of the staff at the University of Illinois Beef Cattle Unit for animal handling and care. S. Moisa, on leave from Facultad de Agronomia, Universidad de Buenos Aires, was supported by a PhD fellowship from Agencia Nacional de Ciencia y Tecnologia and FAUBA of Argentina. There are no financial or other contractual agreements that might cause conflict of interests.