
Abstract
Malaria pathogenesis may be influenced by IgE responses and cytokine cross-regulation. Several mutations in the IL-4/STAT6 signaling pathway can alter cytokine cross-regulation and IgE responses during a Plasmodium falciparum malarial infection. This study investigated the relationship between a STAT6 intronic single-nucleotide polymorphism (rs3024974), total IgE, cytokines, and malaria severity in 238 Ghanaian children aged between 0.5 and 13 years. Total IgE and cytokine levels were measured by ELISA, while genotyping was done by polymerase chain reaction-restriction fragment length polymorphism (RFLP). Compared with healthy controls, heterozygosity protected against clinical malaria: uncomplicated malaria (odds ratios [OR] = 0.13, P < 0.001), severe malarial anemia (OR = 0.18, P < 0.001), and cerebral malaria (OR = 0.39, P = 0.022). Levels of total IgE significantly differed among malaria phenotypes (P = 0.044) and rs3024974 genotypes (P = 0.037). Neither cytokine levels nor IL-6/IL-10 ratios were associated with malaria phenotypes or rs3024974 genotypes. This study suggests a role for rs3024974 in malaria pathogenesis and offers further insights into an IL-4/STAT6 pathway mutation in malaria pathogenesis.
Introduction
Plasmodium falciparum infection results in clinical disease with varying degrees of severity. Although the majority of infected children may remain asymptomatic or develop uncomplicated malaria (UM), about 2% of children develop severe malaria (SM), which sometimes manifest as severe malarial anemia (SMA) or cerebral malaria (CM). 1 The pathogenesis of SM is only partly explained, and host factors as well as parasitic, climatic, and socioeconomic factors have been implicated.2–4 In spite of the insights gained from decades of studying the role of host immunogenetic factors on malaria pathogenesis and susceptibility,5–10 this area of research still has many gray areas. For instance, whereas the importance of imbalances in cross-regulatory cytokines in malaria pathogenesis is well described,11,12 very little is known about the underpinning pathway mutations that may account for such imbalances.
Pathway mutations can affect cytokine cross-regulation by influencing the production of cytokines, which may in turn throw the delicate balance among pro-inflammatory, anti-inflammatory, and regulatory cytokines overboard. In a typical P. falciparum malarial infection, equilibrium in cytokine cross-regulation is maintained through an early production of pro-inflammatory cytokines (such as TNF-α and IL-6), which are required to curb infection. This is followed by a timely release of anti-inflammatory cytokines (such as IL-4 and IL-13), which initiates the downregulation of pro-inflammatory cytokines to prevent immunopathology. Regulatory cytokines (such as IL-10) play a key role in this interplay by regulating the balance between pro- and anti-inflammatory cytokines.11,12 This balance in cross-regulatory cytokines in respect of time and quantity during infection is controlled by a network of signaling pathways including the IL-4/STAT6 pathway. This pathway has been shown to play essential roles in IgE synthesis, Th2 cytokine synthesis, and host of other processes. 13 A number of studies have investigated the effects of IL-4 mutations on IgE levels and malaria severity,5,14,15 but very few have considered signal transducer and activator of transcription 6 (STAT6) mutations and IgE levels in the context of malaria. 16
The STAT6 molecule is a ubiquitously expressed member of the STAT family of transcription factors. It is encoded by 19 kb gene with 23 exons and located on chromosome 12q13.3–q14.1; the same chromosomal region that encodes IFN-γ, neuronal nitric oxide synthase (NOSI), and other genes implicated in the pathogenesis of inflammatory diseases.18,19 Although several STAT6 single-nucleotide polymorphisms (SNPs) have been described, only a few have been studied in the context of infectious diseases. 19 One such STAT6 SNP was associated with urinary schistosomiasis in a Malian population,21,22 and another was associated with clinical malaria in Congolese children. 17 The STAT6 intronic (rs3024974) SNP caused by a C to T transition in intron 18 of the human STAT6 gene is another SNP that could be interesting in malaria pathogenesis. 19 The function of this SNP is not fully known, although studies in Caucasians have associated it with an increased production of total IgE in the context of asthma and atopy. 18 Current literature from both human and murine studies seem to underscore roles for STAT6 in immune-mediated diseases,18,23,24 but few have investigated the role of STAT6 SNPs in the pathogenesis of human malaria.
In the light of possible influence of STAT6 on cytokine cross-regulation and IgE production, a better understanding of the relationship(s) between STAT6 SNPs, cytokines, and IgE production in malaria pathogenesis has become interesting. Although there is some consensus on the role of cross-regulatory cytokines on malaria pathogenesis, the role of IgE has remained controversial.25–27 Although functional malariaspecific IgE is known to be associated with protection from malaria,27,28 total IgE is thought to have a pathogenic role.26,29 It is believed that total IgE in the form of immune complexes cross-link with low-affinity IgE receptors (CD23) and trigger local overproduction of TNF-α and NO, which are both associated with the development of SM. 26 Reconciling the purported role of STAT6 in regulating IgE with the fact that total IgE production is partly influenced by genetic factors,6,15,18,30 it is plausible that STAT6 mutations can also influence the levels of IgE production during a P. falciparum malarial infection.
In this study, we investigated the relationships between rs3024974 variants, total IgE levels, cytokine (TNF-α, IL-6, and IL-10) levels, and malaria severity in Ghanaian children. Although our data suggest that both rs3024974 and total IgE influence malaria pathogenesis, it appears they influence malaria pathogenesis via different mechanisms that are probably mutually exclusive.
Materials and Methods
Study participants
This study was conducted at the Department of Child Health, Korle-Bu Teaching Hospital, and the Noguchi Memorial Institute for Medical Research (NMIMR), both of the University of Ghana in Accra, Ghana. Total IgE measurements were done at the Immunology Department of the Wenner-Gren Institute, Stockholm University, Stockholm, Sweden. The Institutional Review Board of the NMIMR and the Scientific and Ethical Review Committee of the University of Ghana Medical School provided the ethical approval for this research. The research was conducted in accordance with the principles of the Declaration of Helsinki. Only children of parents or guardians who consented to the study by signing written informed consent forms were recruited into the study. Details of sample collection and classification of study participants have been previously described. 30 , 31 Children between 0.5 and 13 years who were brought to the hospital by their parents or guardians for antimalarial medical care were recruited as volunteers for this study. All the children were treated by a qualified medical doctor following the then Ghana Health Services guidelines. Five milliliters of venous blood samples were collected at enrollment into ethylenediaminetetraacetic acid (EDTA) tubes: plasma and buffy coat samples were separated and stored at – 80°C in the hospital. Healthy children without detectable parasites as examined by light microscopy were recruited from neighborhoods close to the hospital and used as a control group. Inclusion and exclusion criteria for patients were as follows: malaria was defined as fever (temperature > 37.5°C) measured within 24 hours of admission, asexual parasitemia of >2,500/μL of blood, and one other sign of malaria (vomiting, diarrhea, malaise, etc.). In addition, recruits had to be sickling and bacteremia negative and show no signs of other diseases. Malaria patients were categorized into UM, SMA, and CM groups. CM was defined as an unarousable coma (score of <3 on the Blantyre coma scale) and no other neurological manifestation; SMA was defined by a blood hemoglobin concentration of <5 g/dL, full consciousness, and no other cause of anemia; UM was defined as described for SMA, except that the hemoglobin concentrations were >8 g/dL. Children who did not strictly meet these criteria were excluded from the study.
Hematological and parasitological measurements
Hematological parameters including hemoglobin levels, total white blood cells (WBC) counts, total red blood cells (RBC) counts, and mean corpuscular volume, or mean cell volume (MCV) were measured with an auto hematological analyzer (Sysmex KX-21). Thick and thin blood films were Giemsa stained and observed under light microscope for parasites. Estimation of malaria parasites was done by counting parasites against 200 WBCs. Parasite densities were estimated by multiplying the total WBC counts by the parasites per 200 WBCs. The sickling status of potential volunteers was determined by sodium metabisulfite test, and sickling-positive patients were excluded from the study.
Measurement of total serum IgE
Enzyme-linked immunosorbent assay (ELISA) plates (Coster) were coated overnight with 50 μL/well of affinity purified goat anti-human IgE (Vector Laboratories) at 5 μg/mL in bicarbonate buffer. Plates were then blocked with 100 μL/well of 0.5% bovine serum albumin (BSA), incubated at 37°C for three hours and washed four times with phosphate buffered saline (PBS) + 0.05% Tween 20. Test sera (50 μL/well) diluted 1:1,000 were added to appropriate wells and incubated at 37°C for one hour. After incubation, 50 μL/well of biotinylated goat anti-human IgE (Vector Laboratories) diluted 1:10,000 with 0.5% BSA in bicarbonate buffer were added to the plates and incubated at 37°C for one hour. This was followed by the addition of 50 μL/well of alkaline phosphatase (ALP)-conjugated streptavidin (Mabtech) diluted 1:2,000 with 0.5% BSA in PBS + 0.05% Tween 20 and incubated at 37°C for one hour. Plates were finally developed with 50 μL/well of p-nitrophenyl phosphate (pNPP; Sigma), and the optical density values were read in a VMax Microplate Reader (Molecular Devices Corporation) at 405 nm. Total IgE concentrations were calculated from standard curves (human serum IgE from NIBSC and titrated threefold from 10 ng/mL over six duplicated wells) included on each plate.
Measurement of serum cytokine levels
Standardized protocols described elsewhere 30 , 31 were used to measure the levels of three cytokines (TNF-α, IL-6, and IL-10) in a subset of samples (60) that had enough sera for the assay. For the measurement of IL-6 and IL-10, 4 μg/mL of the respective purified rat anti-human antibodies obtained from BD Biosciences were coated to MaxiSorp microtiter plates (Nunc) at 50 μL/well and incubated overnight at a temperature of 4°C. This was followed by a washing step involving four washes with PBS-Tween 20 (0.05%) before blocking with 150 μL/well of 5% heat-inactivated fetal bovine serum (FBS) in PBS for an hour at room temperature. The plates were then washed twice with the same washing buffer as above before the addition of recombinant standards (threefold serial dilutions from 4,000 to 1.83 pg/mL; BD Biosciences) and undiluted serum samples (50 μL/well). The plates were then incubated for two hours at room temperature on a shaker. A washing step involving four washes was performed before the addition of 1 μg/mL biotinylated anti-human cytokine antibodies (BD Biosciences) in 5% heat-inactivated FBS in PBS at 50 μL/well. The plates were then incubated at room temperature for 45 minutes. The plates were washed five times before the addition of 0.5 μg/mL streptavidin–peroxidase (1:1,000 dilution of stock in 5% FBS in PBS; KPL) at 50 μL/well, and then incubated again for 30 minutes at room temperature. A final washing step involving eight washes was performed before the plates were developed with orthophenyldiamine substrate for 30 minutes. Optical densities were read at 492/620 nm within 30 minutes. TNF-α ELISA was similar to IL-6 and IL-10 with some variations. Immulon 2 microtiter plates (Thermo Labsystems) were used for the TNF-α ELISA. Purified rat anti-human antibody (2 μg/mL) obtained from BD Biosciences was used as the capture antibody, whereas recombinant human TNF-α (twofold serial dilutions from 2,000 to 15 pg/mL) from the same suppliers were used as the standards. Detection antibody was 1 μg/mL biotin-conjugated mouse anti-human TNF-α from the same supplier. Streptavidin–peroxidase from KPL were added at 0.125 μg/mL (1/4,000 dilution of the stock) before a final wash step involving 10 washes. Color development was done using orthophenyldiamine, and optical density reading was at 492/620 nm.
Genomic DNA extraction and genotyping of STAT6 SNP (rs3024974)
Genomic DNA was extracted from EDTA-preserved buffy coat samples using the QIAGEN DNeasy tissue kit (QIAGEN). The region of the STAT6 gene that contains the rs3024974 SNP within intron 18 was amplified by polymerase chain reaction (PCR) using the oligonucleotide 5′-GAACAGATGGGTAAGGATGG-3′ and 5′-TGACTGACCAAGGGTTGATG-3′ as forward and reverse primers, respectively. Amplification was performed in a 25 μL reaction mix containing 2 μg of genomic DNA, 200 μM deoxyribonucleotide triphosphates (dNTPs) (Sigma), 10 μM of each oligonucleotide primer (Eurogentec), and 1.5 units of Taq polymerase enzyme (Sigma). The reaction mix was centrifuged briefly and overlaid with mineral oil to avoid evaporation and refluxing during thermocycling. PCR was performed with PTC-100 thermal cycler (MJ Research Inc.) with the following cycling parameters: 95°C for 5 minutes (initial denaturation), followed by 45 cycles at 95°C for 45 seconds, 61°C for 45 seconds, 72°C for 45 seconds, and a final extension at 72°C for 10 minutes. A successful amplification gave a PCR product of 157 bp. PCR products were digested using PstI enzyme (New England Biolabs) in a reaction containing 12 μL of amplified product, 0.5 μL of 1 U Pst I, and 2 μL of 1 × NE Buffer 3 (New England Biolabs). Sterile double-distilled water was added to make up the final volume of 20 μL, and digestion was carried out at 37°C for five hours. The digested fragments were then analyzed on 4% agarose gel stained with ethidium bromide (Supplementary Fig. 1). PstI cleaves the C allele of rs3024974 into two fragments of 115 and 42 bp, while the T allele remains intact. The sizes of digested fragments were estimated by comparison with the mobility of a 100 bp molecular marker (Promega).
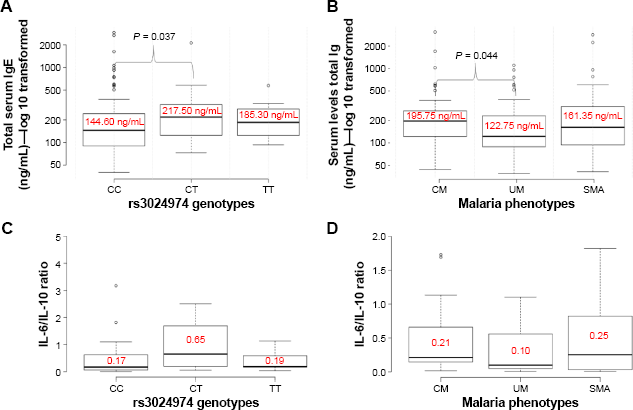
Comparisons of median values of IgE levels and IL-6/IL-10 ratios among malaria phenotypes and rs3024974 genotypes using a Kruskal–Wallis H test. (A) Comparison of total IgE levels among the rs3024974 genotypes, which found median IgE to be significantly higher in the CT group compared with the CC group. (B) Comparison of total IgE among malaria phenotypes, which found median IgE to be higher in the CM group compared with the UM group. Comparisons of IL-6/IL-10 ratios among rs3024974 genotypes (C) and malaria phenotypes (D) did not reveal any significant differences.
Statistical analysis
All statistical analyses were done using R (version 2.11.1, the R Foundation for Statistical Computing). Nonparametric methods for the analysis of variance were employed for data that failed the normality test. For comparisons involving more than two groups, Kruskal–Wallis H test and post hoc pairwise comparison with Bonferroni correction were employed. Conformity of allelic and genetic distribution to Hardy–Weinberg equilibrium was tested using chi square. Odds ratios (OR) were estimated for an association of STAT6 genotypes with the severity of the disease or the level of parasitemia using linear regression models. Pearson correlations were used to determine the association between two continuous variables. Box plots and scatter plots were generated with BoxplotR and Plotly, 32 respectively. Differences and associations were deemed significant at P < 0.05.
Results
General characteristics and malariometric indices among the patient groups
The general characteristics of the study participants are shown in Table 1. A total of 238 children aged between 0.5 and 13 years comprising 157 malaria cases and 81 controls participated in this study. Malaria cases comprised 66 UM patients (median age = 5.0 years), 45 SMA patients (median age = 2.0 years), and 46 CM patients (median age = 3.5 years). A Kruskal–Wallis H test and post hoc pairwise comparison with Bonferroni correction revealed that UM patients had significantly higher median age compared with CM (P = 0.042) and SMA (P < 0.0001) patients. The median ages of CM and UM were, however, not significantly different (P = 0.334). A similar test did not show a significant difference between the parasite densities in UM and CM patients (P = 0.651). However, parasite density was significantly lower in the SMA group compared with UM (P = 0.004) and higher in CM patients compared with SMA patients (P = 0.008).
General characteristics of study participants with clinical malaria.

P = 0.03,
P < 0.0001
P = 0.651
P = 0.004.
Distribution of rs3024974 SNP among malaria cases and controls
Allelic and genotypic distributions of rs3024974 in the study population are shown in Table 2. The genotype distribution of the rs3024974 was in Hardy–Weinberg equilibrium in the controls (P = 0.43) but not in the malaria cases (P = 0.00018).
Genotypic and allelic frequencies of rs3024974 among malaria phenotypes and controls.

The frequencies of the rs3024974 genotypes CC, CT, and TT within the study population were 57%, 25%, and 8%, respectively. The wild-type C allele and the mutant T allele had frequencies of 0.79 and 0.21, respectively, in the whole study population. The frequencies of the C allele in the UM, CM, and SMA disease categories, and controls were 0.9, 0.77, 0.64, and 0.64, respectively. Interestingly, none of the SMA patients had the TT genotype.
Genotypic frequencies of rs3024974 were compared among cases and controls to determine which genotypes were associated with disease. Comparing cases and controls, using the wild-type genotype (CC) as the reference genotype, children with the heterozygous genotype (CT) were less likely to have clinical malaria of any form: UM (OR = 0.13, 95% confidence interval [CI] = 0.05–0.32, P < 0.001), SMA (OR = 0.19, 95% CI = 0.07–0.48, P < 0.001), and CM (OR = 0.39, 95% CI = 0.16–0.88, P = 0.022; Table 3). Similarly using the CC genotype as reference, children with the TT genotype were less likely to suffer from UM (OR = 0.16, 95% CI = 0.04–0.59, P = 0.006) but not CM (OR = 0.49, 95% CI = 0.15–1.54, P = 0.22).
Association of rs3024974 with malaria phenotypes.

None of the SMA patients had the TT genotype.
Total IgE levels in malaria phenotypes and rs3024974 genotypes
To better understand the association between rs3024974 and total IgE production, we compared the median levels of total IgE among the various genotypes using a Kruskal–Wallis H test and post hoc pairwise comparison with Bonferroni correction (Fig. 1A). The rs3024974 CT genotype had significantly higher levels of total IgE than the CC genotype (P = 0.037). No such difference was observed when the TT and CC genotypes were compared (P = 0.404). A similar comparison of total IgE levels among rs3024974 genotypes within the various disease categories (UM, CM, and SMA) did not reveal any significant differences.
To gain further insight into the role of total IgE in malaria pathogenesis, we also compared the median levels of total IgE among malaria phenotypes using the same test (Fig. 1B). Total IgE levels were found to be significantly higher in CM (195.8 ng/mL) than in UM (122.8 ng/mL; P = 0.044) patients. However, there was no evidence of a significant difference in the total IgE levels between CM and SMA patients (P = 0.552) as well as between SMA and UM patients (P = 0.223).
Serum cytokines and parasite density among rs3024974 genotypes and malaria phenotypes
To better understand the relationship between rs3024974 genotypes and parasite density, we compared median parasite density among the genotypes. Parasite density was not significantly different among the rs3024974 genotypes (P = 0.4057, Kruskal–Wallis H test; Fig. 2). Variations in the levels of cytokines and the ratio of IL-6/IL-10 among the various genotypes were determined to gain further insight into the association of rs30224974 with cytokine cross-regulation. There was no significant difference in the cytokine levels among rs3024974 genotypes and malaria phenotypes (data not shown). There was no significant difference in IL-6/IL-10 ratio among the rs3024974 genotypes (P = 0.1629, Kruskal–Wallis H test; Fig. 1C) and malaria phenotypes (P = 0.6557, Kruskal–Wallis H test; Fig. 1D). None of the cytokines studied showed any association with parasite density except IL-6, which was positively correlated with parasite density in the UM group (r = 0.7, P = 0.003; Pearson correlation; Fig. 3D). Even when not statistically significant, IL-6 appeared to be positively correlated with parasite density in all malaria phenotypes except in the SMAs (r = –0.3, P = 0.282; Fig. 3C). None of the cytokines measured showed any relationship with the levels of total serum IgE.
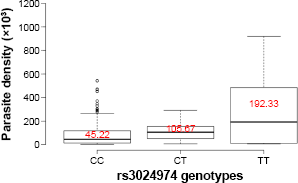
Comparison of the median levels of parasite density among the rs3024974 genotypes. A Kruskal–Wallis H test did not reveal any significant difference.
Discussion
This study investigated the relationship between a STAT6 intronic SNP, total IgE, and malaria severity in Ghanaian children. The genotypic distribution of the rs3024974 SNP in cases and controls suggests that this SNP may be associated with malaria in Ghanaian children. Most genetic studies use conformity to hardy-weinberg equilibrium (HWE) as an indicator of proper sampling and genotyping. 33 However, a deviation from HWE can also be an indicator of the existence of a gene–disease association. 34 To this end, screening for deviations from HWE in datasets of cases has become a reasonably robust method for detecting gene–disease associations. 34 In this study, conformity to HWE was seen in the control group only. Therefore, deviation from HWE in cases is consistent with our main findings, which suggest that this SNP could be associated with malaria in our study population.
The association of CM with heterozygosity but not with recessive homozygosity is intriguing but not inexplicable. This observation could be explained by the heterozygote advantage phenomenon. The heterozygous advantage is the situation where the heterozygous genotype has a higher relative fitness than either the homozygous dominant or the homozygous recessive genotypes. It is epitomized in the interaction between the sickle cell gene and malaria in endemic regions. 35 Regardless of its rare nature in the study population, it was interesting to observe that the TT genotype was absent in the SMA group. This is a noteworthy observation, although the absence of the T allele in the SMA group made it impossible for us to do any statistical analysis involving this group.
In line with the previous studies that suggested a role for total IgE in the pathogenesis of CM,25,26,36,37 our study found higher titers of total IgE in CM patients compared with their UM counterparts. Although the role of total IgE in the pathogenesis of malaria is yet to be fully clarified,26,27 it is believed to be involved in the immunopathogenesis of SM.29,36,37 In its pathogenic role, total IgE is thought to contribute to CM pathogenesis by forming immune complexes that cross-link with leukocytes to induce local overproduction of TNF-α, which is a pathogenic factor in CM.37,38 The literature on the protective and/or pathogenic role of total IgE is so far equivocal,21,26,28,36,37 and deciphering the exact role of IgE in malaria pathogenesis will require further investigation. This has become particularly important in the light of the recent evidence that free serum IgE may not necessarily reflect the body's total IgE pool. 39 In fact, high levels of cell-bound IgE have been detected even when serum IgE levels are low. 39
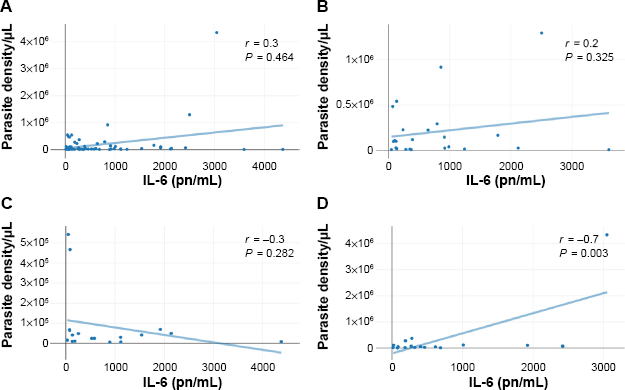
Correlations between plasma IL-6 levels and parasite density in the entire study population (A), CM patients (B), SMA patients (C), and UM patients (D). The P values were obtained after a Pearson product correlation test.
The association between rs3024974 variants with the total IgE levels observed in this study is consistent with an earlier report. 18 However, finding higher levels of total IgE in the heterozygous CT group of the rs3024974 variant does not support an IgE-mediated mechanism of protection against malaria. It is possible that rs3024974 SNP mediates malaria pathogenesis via other mechanisms besides IgE that were not considered in this study. Similarly, the variation in parasitemia among rs3024974 variants suggests that the rs3024974 may not offer protection from malaria via mechanism(s) that control parasitemia.
The inflammatory factors influencing the outcome of a P. falciparum malarial infection are complex. In mice, stat6 deficiency affects the T1/T2 balance by inducing a T2 environment, which eventually leads to the lack of IgE class switching. 40 Some of these studies have suggested roles for STAT6 in the induction of Th2 cytokines. 20 Although previous human studies have highlighted the role of various cytokines in malaria pathogenesis,41–43 this study failed to confirm well-known associations between cytokines (IL-6, TNF-α, and IL-10) and malaria severity, 11 probably due to sample size constraints. Our limitations notwithstanding the evidence from other studies have suggested that STAT6 can influence the production of TNF-α, IFN-γ, and IL-12. 20 Furthermore, a STAT6-dependent pathway for the pathogenesis of SMA in mice has been proposed by Thawani et al. 20 The study and its commentaries further suggested roles for IL-4 and IFN-γ in a STAT6-mediated erythropoietin suppression.21,22 The IL-6/IL-10 ratio, which is somewhat indicative of the prevailing inflammatory profile, was neither associated rs3024974 genotypes nor malaria phenotypes. The positive correlation of IL-6 with parasite density in the UM group alone suggests that IL-6 may play different roles in different malaria phenotypes.
Taken together, data from this study suggest that rs3024974 and total IgE may influence malaria pathogenesis differently and independently. Total IgE was associated with CM, whereas rs3024974 was associated with protection from clinical malaria in any form. It is possible that rs3024974 SNP influences malaria pathogenesis through other factors not considered in this study. The STAT6 signaling cascade involves several other molecules that could influence the pathogenesis of P. falciparum malaria. This current study provides fresh insights into the possible roles of a pathway mutation and total IgE levels in malaria pathogenesis. It also provides vital information for future studies that may want to consider pathway mutations in the IL-4/STAT6 signaling pathway in malaria pathogenesis and susceptibility.
Author Contributions
Conception and study design: DA-S and BAG. Experimentations: DA-S, KAK, and SA. Contribution toward reagents/materials/analysis tools: BAG, DD, MFO, BDA, BA, BQG, GOA, DO-Y, RHA, CB, and GF. Involvement in data analysis: DA-S, SA, and KAK. Initial draft of the manuscript: DA-S, SA, and KAK. Intellectual review of manuscript: DA-S, SA, KAK, DD, MFO, GOA, DEE, RHA, CB, BA, DO-Y, GF, SBA, MT-B, BDA, BQG, JA-M, and BAG. Collected clinical and demographic data: DA-S, BDA, BQG, GOA, and SA. All authors reviewed and approved of the final manuscript.
Supplementary Material
Footnotes
Acknowledgments
We thank the study participants and their parents/guardians for agreeing to partake in the study. We also thank the staff of Immunology Department of Noguchi Memorial Institute for Medical Research and the staff of the Department of Molecular Biosciences, the Wenner-Gren Institute, Stockholm University.