
Abstract
While ticks have been known to harbor and transmit pathogenic arboviruses for over 80 years, the application of high-throughput sequencing technologies has revealed that ticks also appear to harbor a diverse range of endogenous tick-only viruses belonging to many different families. Almost nothing is known about these viruses; indeed, it is unclear in most cases whether the identified viral sequences are derived from actual replication-competent viruses or from endogenous virus elements incorporated into the ticks' genomes. Tick cell lines play an important role in virus discovery and isolation through the identification of novel viruses chronically infecting such cell lines and by acting as host cells to aid in determining whether or not an entire replication-competent, infective virus is present in a sample. Here, we review recent progress in tick-borne virus discovery and comment on the actual and potential applications for tick cell lines in this emerging research area.
Ticks have been recognized as vectors of pathogenic protozoa and bacteria for over 120 years.1–3 The first confirmed tick-borne viruses were identified in the early 1930s causing disease in sheep: the flavivirus louping ill virus, transmitted by Ixodes ricinus in Scotland, 4 and the nairovirus Nairobi sheep disease virus, transmitted by Rhipicephalus appendiculatus in Kenya. 5 Since then, ticks have been incriminated as vectors and/or reservoirs of numerous viruses of medical and/or veterinary importance,6,7 including the emerging pathogens Alkhumra hemorrhagic fever virus, severe fever with thromocytopenia virus, Heartland virus, and Bourbon virus.8–11 In addition, many apparently nonpathogenic arboviruses have been isolated from ticks.6,7
Research on tick-borne viruses has benefited greatly from the availability of tick cell and tissue cultures. Almost as soon as techniques were developed for repeatedly producing primary tick cell cultures,12,13 they were put to use for experiments on arthropod-borne and other viruses, confirming the ability to replicate in tick cells and helping to clarify their host range.14–17 Continuous cell lines, first derived from ticks in the 1970s,18,19 have since been applied to many aspects of arbovirus research,11,20–25 including clarification of actual and potential vector capacity.11,14,15,21,22,26–28 Replication was demonstrated in tick cells of viruses carried by ixodid and argasid ticks, mosquitoes, sand flies and midges as well as some viruses with no known vector.15,20,29–31 Surprisingly, there are few reports of tick cells being used for the isolation of tick-borne viruses from field samples; primary tick cell cultures were found to be at least as sensitive as chick embryo cell cultures for the isolation of tick-borne encephalitis virus (TBEV) from vertebrate blood and brain suspension and tick suspension.32,33 An uncharacterized RNA virus, Cascade virus, was isolated from frozen triturated Dermacentor occidentalis ticks into a Dermacentor variabilis cell line (RML-15); Cascade virus also replicated well in amphibian (XTC-2) cells but poorly or not at all in mosquito cells, most vertebrate cells, and suckling mice. 34 The Rhipicephalus microplus cell line BME26 was used recently alongside Vero and BHK-21 cells for the isolation of a putative novel flavivirus, Mogiana tick virus from R. microplus ticks in Brazil. 35
Until recently, all the viruses detected in ticks had one characteristic in common-they could all replicate in vertebrate cells and indeed were initially isolated into vertebrate cells. However, it is now recognized that ticks themselves harbor a diverse microflora of vertically transmitted endosymbiotic bacteria and endogenous viruses36–40 that may not cycle between ticks and their vertebrate hosts. While endosymbiotic bacteria have been reported to occur in various different tick species for nearly a century,3,41–44 very little is known about endogenous viruses of ticks. 45 An electron microscope study revealed the presence of virus-like particles in the salivary glands of adult female R. microplus ticks feeding on cattle 46 ; the presence of large numbers of the particles was associated with possible abnormal tickfeeding behavior. A single apparently tick-only virus has been comprehensively described, sequenced, and characterized: the orbivirus St Croix River virus (SCRV) 47 was detected in, and replicates in, tick cell lines but has so far failed to infect or replicate in any of the vertebrate or insect cell lines tested. 43 Originally, SCRV was detected in the IDE2 cell line derived from the tick Ixodes scapularis,47,48 suggesting that this prostriate tick was its natural host; subsequent detection of SCRV in two cell lines (RA243 and RA257) derived from developing adult R. appendiculatus ticks established nearly 20 years previously in a laboratory on a different continent18,49 suggests that the virus may have originated from the latter metastriate tick species (Table 1). Electron microscopy revealed the presence of many more putative and potentially tick-only viruses in the majority of over 50 cell lines derived from 14 ixodid and two argasid tick species (Table 1).45,49,50 These tick cell lines represent an abundant, easily accessible resource for virus discovery, as well as contributing to studies on virus ancestry and evolution. Studies on the evolution of SCRV and other tickborne orbiviruses51–53 suggested that SCRV is the oldest orbivirus identified to date and provided evidence of coevolution of orbiviruses and their respective arthropod host/vector. More recently, the Amblyomma americanum cell line AAE12 54 was found to contain novel orbivirus sequences (H. Attoui, P. Alberdi, C. Sharp, and L. Bell-Sakyi, unpublished data, 2012) (Table 1); if these sequences belong to a replication-competent virus, it will be interesting to determine its relationship to SCRV and other orbiviruses.
Viruses and viral RNA sequences detected in tick cell lines. The tick species and instar from which each cell line was derived have been presented previously (49).
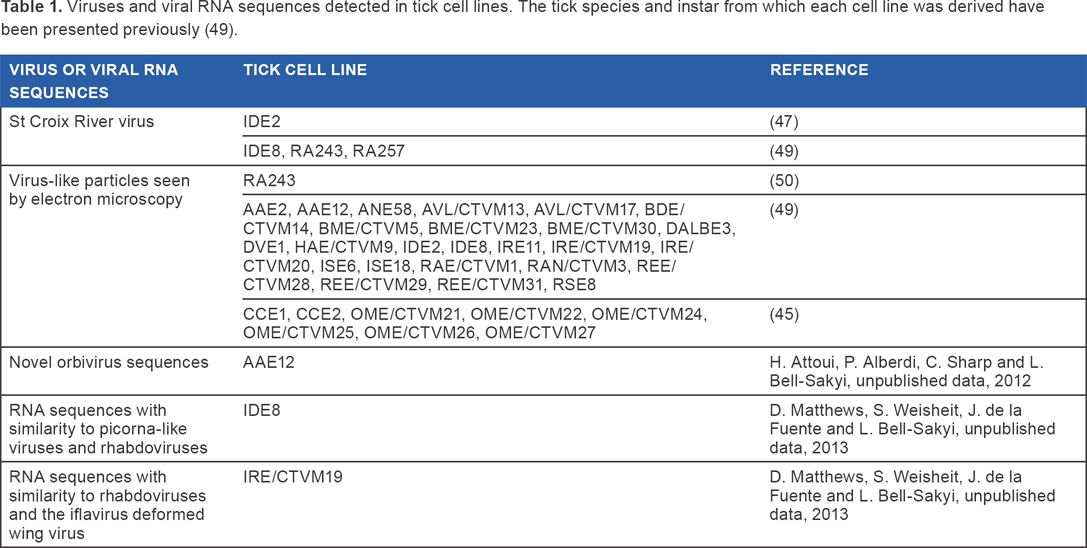
In the past few years, high-throughput molecular screening and deep sequencing approaches applied to various tick species36,38,40,55,56 have revealed tantalizing hints of many more potentially tick-only viruses belonging to diverse families, but for such putative viruses, if they cannot infect and replicate in vertebrate cells, tick cell lines will be needed to establish whether the sequences belong to intact, replication-competent viruses or to endogenous viral elements (partial or full-length DNA copies of the RNA genomes integrated into the genome of the vector/host; EVE) incorporated into the tick genomes. A proteomics informed by transcriptomics approach applied to RNAseq data from uninfected and TBEV-infected tick cells 24 revealed RNA sequences with similarity to picorna-like viruses and rhabdoviruses in the I. scapularis cell line IDE8 and rhabdoviruses and the iflavirus deformed wing virus in the I. ricinus cell line IRE/CTVM19 (D. Matthews, S. Weisheit, J. de la Fuente, and L. Bell-Sakyi, unpublished data, 2013) (Table 1). Rhabdovirus-like (weakly grouped with lyssaviruses), orthomyxovirus-like (related to quarjaviruses), and bunyavirus-like (related to Hazara and Uukuniemi viruses) endogeneous virus elements were identified in the genome of I. scapularis, 57 the only tick species with a completely sequenced genome (https://www.vectorbase.org/). 58 RNA sequences with similarity to genes of insect densoviruses, hepatitis E virus and the tick-borne pathogen African swine fever virus were detected in transcriptomes of R. microplus larvae and/or “tick gut” (unspecified life cycle stage) available on the CattleTickBase website (http://cattletickbase.ccgapps.com.au/). These data are not published, and it is unclear whether they represent authentic viral transcripts or EVE transcripts. It is noteworthy that reverse transcriptase activity, as well as retrotransposons, was detected in three of the I. scapularis embryo-derived cell lines (IDE2, IDE8, and ISE6) (H. Attoui and F. Mohd Jaafar, unpublished data, 2006). Reverse transcriptase activity may explain why DNA copies of RNA virus genomes occur in the I. scapularis genome. Interestingly, it has been shown that TBEV infection of Hep2 human cells resulted in the production of DNA copies of the viral genome and the stable integration of this DNA into the cellular genome backbone 59 and that TBEV DNA was found integrated into brain cells during progressive infection. 60 Whether or not a similar process occurs in TBEV-infected tick cells remains to be determined.
In mammalian genomes, endogenous retroviruses have been shown to exert essential regulatory gene expression functions, 61 when integrated in specific positions of the mammalian genome. A decade ago, we identified a DNA form of a virus related to cell fusing agent virus in the Aedes albopictus mosquito cell line C6/36, including a 1557 amino acid open reading frame containing the NS1 to NS4B. 62 Antibodies raised against recombinant NS1 and NS3 of this virus helped to identify expression of its NS3 in C6/36 cells (H. Attoui and F. Mohd Jaafar, unpublished data, 2006). The Dicer-2 of C6/36 cells is nonfunctional as a result of mutations, 63 making them highly permissive to a range of arboviruses; expression of viral protein such as NS3 (viral protease) from EVE may also enhance permissivity of cells to viral infections. Similar situations may exist in tick cells in which EVE are expressed.
In conclusion, the present portfolio of cell lines derived from different ixodid and argasid tick species, and new cell lines under development from additional species, constitutes a huge and hitherto underexplored and underutilized resource for the discovery, detection, isolation, and propagation of novel arthropod-borne viruses. It is likely that the application of molecular technologies such as those used in the studies mentioned previously,35,36,38,40,55,56 and novel pipelines for metagenomic analysis, 64 will very soon start to reveal the secrets of tickborne viruses and endogenous viral elements harbored by tick cell lines and, by inference, the ticks from which they were derived.
Author Contributions
Composed, reviewed, and approved the final article: LB-S and HA.
Footnotes
Acknowledgments
The authors would like to thank Ulrike Munderloh and Timothy Kurtti of the University of Minnesota for providing tick cell lines.