
Abstract
Several agents are known to improve sleep induction and/or maintenance in patients with insomnia disorder. These include the benzodiazepine (BZD) and non-BZD receptor allosteric modulators, the melatonin receptor agonist ramelteon, low-dose doxepin, and suvorexant. One of the drawbacks of the BZDs is their known reduction in both N3 sleep [also known as slow wave sleep or delta sleep and characterized by the occurrence of slow high amplitude delta (0.5–2 Hz) waves] and rapid eye movement (REM) sleep. Low-dose doxepin has shown similar association with decrease in REM sleep. By contrast, suvorexant increases REM sleep. The available evidence tends to indicate that irrespective of their mechanisms of action, the selective serotonin 5-HT2A receptor antagonists and inverse agonists, including volinanserin, pruvanserin, and nelotanserin, when given in isolated administration, increases slow wave sleep in laboratory animals. Wakefulness and REM sleep were decreased in some studies. Moreover, subjects with normal sleep showed significant increase in N3 sleep following the administration of eplivanserin, nelotanerin, and pimavanserin. Nelotanserin has also been shown to augment N3 sleep in patients with chronic insomnia disorder. N2 sleep tended to decrease in most of these studies, while REM sleep showed no significant changes. Taken together, these evidences suggest that the coadministration of a selective 5-HT2A receptor antagonist or inverse agonist with a hypnotic drug could be a valid clinical strategy for normalizing sleep induction and maintenance and for promoting N3 sleep in patients with insomnia disorder. Additionally, the 5-HT2A receptor agents may have a potential value for improving the cognition and memory deficits in patients with a chronic insomnia disorder as well as elderly patients who show reductions in N3 sleep.
Normal Sleep
The scoring manual of American Academy of Sleep Medicine 1 provides a classification system of sleep stages that comprises the following three different states: wakefulness (W), non-rapid eye movement (NREM) sleep, and rapid eye movement (REM) sleep. Relaxed W is characterized by an electroencephalogram (EEG) with alpha activity (8–12 Hz) intermixed with lower amplitude irregular beta waves (13–35 Hz), slow or rapid electrooculogram, eyelid blinks, a relatively high tonic electromyogram (EMG), and movement artifacts. NREM sleep is characterized by the following: stage N1 sleep represents the transition from W to sleep and is characterized by the presence of relatively low-voltage, mixed frequency waves (4–7 Hz range) with a prominence of activity in the theta range. It is viewed as a shallow sleep during which the subject can be easily aroused. Stage N2 sleep is the first unequivocal stage of sleep, which is characterized by the presence of two types of EEG signals, namely the sleep spindles (12–14 Hz) and the K-complexes. During this stage, a more intense stimulus is needed to arouse the subject. Stage N3 sleep is characterized by the occurrence of slow high-amplitude delta waves (frequency of ~0.5–2 Hz). It is also known as slow wave sleep (SWS) or delta sleep. REM sleep is characterized by the presence of low-voltage mixed frequency EEG activity, which closely resembles that of stage N1 sleep. Often observed against this background are sawtooth waves (theta activity, 4–7 Hz) in conjunction with bursts of REMs. In contrast, muscles are completely relaxed (with the exception of the extraocular muscles and the diaphragm). However, the flat EMG tracing can be periodically interrupted by muscle twitches. Of note is the fact that typically the first half of the night is dominated by stage N3 sleep, whereas REM sleep predominates in the second half of the night.
Age and gender variables have been shown to have associations with the duration of sleep stages. Young adults spend 20%–28% of a night's sleep (seven to eight hours) in REM sleep, 4%–5% in stage N1 sleep, 46%–50% in stage N2 sleep, and 20%–24% in stage N3 sleep. 2 The amount of time spent in each sleep stage does not differ significantly between males and females who are in the same age range. A shortening of sleep duration, which occurs in association with advancing age, is mainly related to the reduction in N3 sleep and REM sleep. 2
The electrographic activity of the rat, a species commonly used in preclinical studies of sleep, has been subdivided into various categories based on the predominant wave forms that are found in each: W [low-voltage fast waves in frontal cortex,a mixed theta rhythm (4–7 Hz) in occipital cortex and relatively high EMG activity], light sleep (high-voltage slow cortical waves interrupted by low-voltage fast EEG activity), SWS (continuous high amplitude slow frontal and occipital waves combined with a reduced EMG), and REM sleep (low-voltage fast frontal waves, a regular theta rhythm in the occipital cortex, and an EMG profile, which is mainly uneventful except for occasional myoclonic twitching). 3
The exact contribution of sleep stages to physiological processes remains unclear to date. Notwithstanding this, there is converging evidence that SWS contributes to memory consolidation processes. 4 In this respect, the active systems’ consolidation hypothesis proposes that during stage N3 memory representations are transferred into neocortical regions where they become integrated into preexisting knowledge networks. 4 On the other hand, the synaptic homeostasis hypothesis assumes that the synaptic plasticity of the cerebral cortex is highly correlated with the synchronized neuronal activity that occurs during N3 sleep. 5 However, these hypotheses are not necessarily mutually exclusive and may overlap. According to Backhaus et al. 6 , sleep-dependent declarative memory (capacity for conscious recollection of facts or events) is impaired in patients with insomnia disorder when compared to subjects with normal sleep. Moreover, in healthy subjects, performance change on memory tests is associated with the amount of N3 sleep, whereas in patients with insomnia whose N3 sleep values are significantly decreased, the memory task performance is closely related to the amount of REM sleep. These findings tend to suggest that the time spent in REM sleep may possibly compensate for the deleterious effects on the memory of N3 sleep loss. The finding of almost no impairment in either verbal or visual declarative memory consolidation in patients with insomnia whose N3 sleep values are similar to those of controls further supports the hypothesis that N3 sleep may play a consolidating role in sleep-dependent declarative memory.7,8 N3 sleep may also play a consolidating role in nondeclarative memory. Compared to healthy controls, patients with insomnia disorder and a marked decrease in N3 sleep fail to show a significant overnight skill improvement on nondeclarative memory tasks (heterogeneous nonconscious learning capacities expressed through behavior and that do not afford the awareness of any memory content) to which they have been exposed prior to go into sleep.8,9 These findings, thus, point to the importance of stage 3 sleep in normalizing the restorative functions of sleep and possibly in promoting the synaptic plasticity. They further support the inference that stage 3 sleep may have a causal primacy in obviating the deficits of memory impairment and performance, which are commonly seen in patients with a chronic insomnia disorder and in elderly patients. While further studies are needed to verify this view, a positive confirmation would clearly have implications for clinical treatment. If verified, the findings of the studies described earlier would suggest that clinical strategies should prioritize the prevention of stage N3 sleep reduction in the affected patients. Such strategies could emphasize nonpharmacological interventions and pharmacological agents, which have been shown to increase SWS. The former include, among others, acoustic stimulation, 10 while the latter involve the administration of drugs that promote SWS.
This article focuses on the effect of selective serotonin 5-HT2A receptor antagonists and inverse agonists on sleep variables in laboratory animals and humans, with special emphasis on N3 sleep. In addition, evidence is considered that these agents may play a role in the improvement of cognition and memory deficits in patients with a chronic insomnia disorder and in elderly patients.
Chronic Insomnia Disorder
According to the International Classification of Sleep Disorders, third edition, 11 a chronic insomnia disorder is characterized by one or more of the following: difficulty in initiating sleep [sleep-onset latency (SOL) of more than 30 minutes], difficulty in maintaining sleep (total sleep time of less than 5.5 to 6 hours), and waking up earlier than desired with inability to resume sleep. Common daytime complaints include the impairments of attention, concentration and memory, fatigue, impaired social and occupational performance, irritability, daytime sleepiness, and reduced motivation. In addition, patients with a chronic insomnia disorder are at risk for injury, drowsiness while driving, and illness. The sleep disturbance and associated daytime symptoms must have occurred at least three times per week and must have been present for at least three months.
Furthermore, compared to healthy subjects, a great number of patients with a chronic insomnia disorder have found to show a significantly reduced SWS and REM sleep and an increased wake time after sleep onset (WASO). 12
The International Classification of Sleep Disorders, second edition, 13 describes several comorbid insomnias arising from causative conditions. Accordingly, a chronic insomnia disorder can be precipitated or aggravated by a psychiatric condition (major depressive disorder, bipolar disorder, anxiety disorder, and schizophrenia) or neurological disease (Alzheimer's disease, Parkinson's disease, epilepsy, ischemic stroke, and headache), a general medical condition (cardiovascular, respiratory, gastrointestinal, renal, endocrine, neoplastic, and rheumatic diseases), a disturbance of circadian rhythm, or the direct effects of a medication or a substance of abuse. Evidence from several studies suggests that gender differences may play a role in the development of a chronic insomnia disorder. It has been shown for instance that sleep complaints are common during the menstrual cycle in young healthy women, as well as in pregnant and postmenopausal women. 11 Compared to their prepregnancy period, pregnant women have reported having more complaints of insomnia during their first trimester, as well as an increased number of awakenings and nonrestorative sleep. Although their sleep tends to normalize during the second trimester, pregnant women's difficulties in falling asleep and staying asleep worsen during the third trimester.14,15 A survey of postmenopausal women found that insomnia was one of the somatic symptoms that they experienced, in addition to difficulty in falling asleep, an increased nocturnal restlessness and awakenings. In addition, they reported feeling tired in the morning and during the daytime. 16 The prevalence of a chronic insomnia disorder has been shown to vary between 10% and 15% of the adult population. It is greater in older adults, and among this group, women show a significantly greater prevalence of the condition. Moreover, depression or anxiety disorders have been diagnosed in 40% of patients with a chronic insomnia disorder. 13
Drugs Approved for the Treatment of a Chronic Insomnia Disorder
Treatment of a chronic insomnia disorder comprises both behavioral and pharmacological therapies. Several classes of medications have been approved by international agencies for the treatment of a chronic insomnia disorder. 17 These include the benzodiazepine (BZD) receptor allosteric modulators (either BZD [triazolam, temazepam, and flurazepam] or non-BZD [zolpidem, eszopiclone, and zaleplon] agents), the melatonin receptor agonist ramelteon, and low-dose doxepin (a tricyclic antidepressant). Concerning the sleep promoting orexinergic inhibitors, the dual orexin receptor antagonist suvorexant has been recently approved for the treatment of an insomnia disorder. 18
The sleep induced by BZD hypnotics, including temazepam and flurazepam, in patients with a chronic insomnia disorder is characterized by a shortened SOL, a decreased number of nocturnal awakenings, the reductions in the time spent awake, an increased amount of stage N2 sleep, a consistent reduction in stage N3 sleep, a dose-dependent decrease in REM sleep, and the improvement of the subjective quality of sleep when compared to no treatment (Table 1).2,19 With respect to the non-BZD agents, immediate-release zolpidem is capable of reducing SOL and WASO in patients with a chronic insomnia disorder. In relation to sleep architecture, immediate-release zolpidem tends to increase N2 sleep, whereas N3 sleep and REM sleep are not significantly modified in most patients. 20 In contrast, zolpidem extended-release is effective for the treatment of insomnia characterized by difficulties in sleep onset and/or sleep maintenance. Notwithstanding this, sleep architecture, in particular SWS and REM sleep, shows no changes, while the amount of time spent in stage 2 sleep is increased during its administration (Table 1).21,22
The effects of benzodiazepine and non-benzodiazepine receptor allosteric agonists on sleep variables in patients with a chronic insomnia disorder.
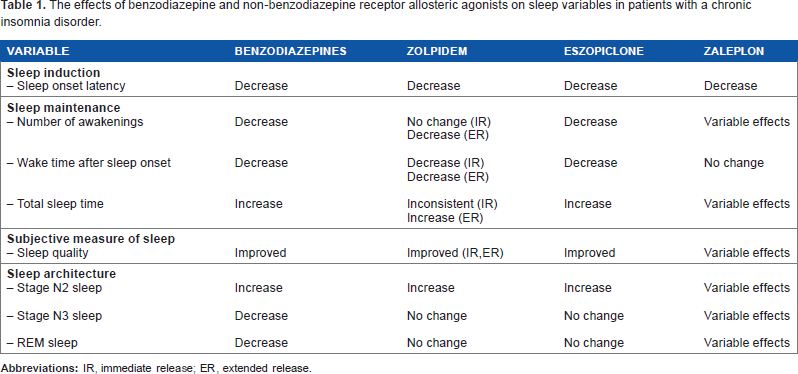
The effects of eszopiclone have been assessed also in patients with a chronic insomnia disorder. Eszopiclone administration significantly reduced SOL and WASO, whereas total sleep time (TST) and sleep efficiency (ratio of total sleep time to time in bed [SE]) showed significant improvements. N2 sleep was significantly augmented, whereas N3 sleep and REM sleep remained below normal levels (Table 1). 23
Polysomnographic data and subjective measures in adult patients with chronic insomnia disorder have shown that zaleplon significantly decreased SOL relative to placebo. However, no consistent effects on WASO, TST, or number of awakenings were seen with the hypnotic drug. Furthermore, zaleplon administration produced inconsistent changes in sleep architecture, including N2 sleep, N3 sleep, and REM sleep (Table 1). 24
The effects of ramelteon have also been evaluated in adult patients with chronic insomnia. Compared to the placebo group, patients taking the melatonin receptor agonist showed reductions in SOL. Their TST was increased, but the effect was restricted to the first week of treatment. The number of awakenings, WASO, and sleep architecture in the ramelteon group were not significantly modified (Table 2). 25
The effects of ramelteon, low-dose doxepin and suvorexant on sleep variables in patients with a chronic insomnia disorder.
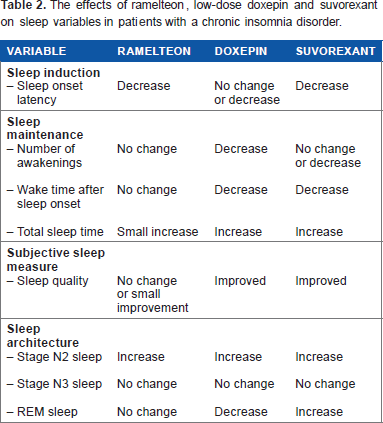
In adult patients with a chronic insomnia disorder, low-dose doxepin significantly improved WASO, TST, and SE. There was a significant increase in N2 sleep, whereas REM sleep was reduced. Values corresponding to N3 sleep remained unchanged (Table 2). 26
The effects of suvorexant have been assessed in young- and middle-aged patients with chronic insomnia. The orexin antagonist reduced SOL and increased TST. The greater amounts of TST were related to an increase in REM sleep and N2 sleep (Table 2). 27
In conclusion, current therapeutic approaches for the treatment of a chronic insomnia disorder address SOL (immediate-release zopidem, zaleplon, and ramelteon) and/or sleep maintenance (temazepam, flurazepam, zolpidem extended-release, eszopiclone, and low-dose doxepin). However, during their administration, N3 sleep and REM sleep do not regain normal sleep and indeed may even be reduced in most patients. With respect to suvorexant, currently available evidence tends to indicate that the compound increases REM sleep and N2 sleep in patients with an insomnia disorder.
Role of Serotonin 5-HT2A Receptor in the Regulation of Behavioral States
The 5-HT2A receptor consists of three (5-HT2A/2B/2C) subtypes and is structurally related to the superfamily of G-proteincoupled receptors. The 5-HT2A receptor is located within postsynaptic structures, predominantly on proximal and distal dendritic shafts. 28 5-HT2A receptors are distributed in brain regions involved in the promotion of (1) the waking state (basal forebrain, ventral tegmental area, dorsal raphe nucleus, locus coeruleus, and tuberomammillary nucleus), (2) the NREM sleep state (medial and lateral preoptic area), and (3) the REM sleep state (laterodorsal and pedunculopontine tegmental nuclei and sublaterodorsal tegmental/precoeruleus nucleus). Additionally, 5-HT2A receptors have been characterized in the cerebral cortex, limbic system, and basal ganglia. 29
Selective 5-HT2A Receptor Antagonists and Inverse Agonists
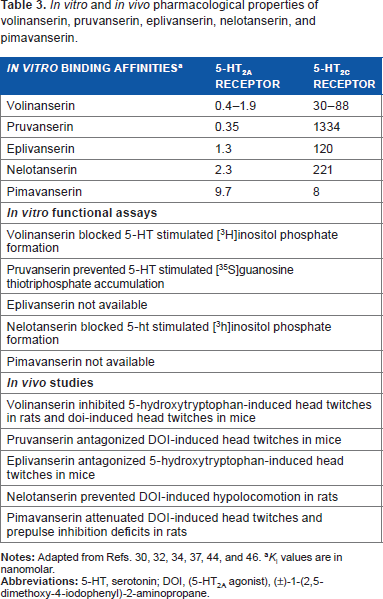
There are two types of drugs that can inhibit the activity of 5-HT2A receptor, namely the silent 5-HT2A receptor antagonists and the inverse agonists. The former block agonist-induced responses, while the latter attenuate the basal constitutive signaling activity of 5-HT2A receptors. Studies aimed at characterizing the effect of 5-HT2A receptor antagonists and inverse agonists on sleep variables in laboratory animals and/or humans have been limited to volinanserin, pruvanserin, eplivanserin, nelotanerin, and pimavanserin, respectively.30–33
Sleep Patterns in Laboratory Animals, Normal Subjects, and Patients with Insomnia Disorder Administered Selective 5-HT2A Receptor Antagonists and Inverse Agonists
Volinanserin
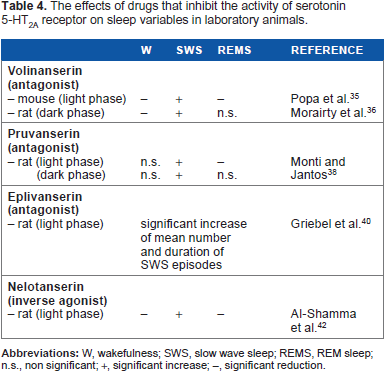
Kehne et al. 34 evaluated the intrinsic activity of the selective 5-HT2A receptor antagonist volinanserin, a chiral phenetyl piperidine. Volinanserin showed a high degree of selectivity for the 5-HT2A receptor, with more than 100-fold separation from the 5-HT2C receptor. Volinanserin in vitro antagonized the 5-hydroxytryptamine (5-HT)-stimulated inositol triphosphate accumulation in cells transfected with the rat 5-HT2A receptor. In in vivo, volinanserin reduced 5-meth-oxy-N,N-dimetyltryptamine and (±)-1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane (DOI)-induced head twitches in mice, and 5-hydroxytryptophan-induced head twitches in rats (Table 3). 34 Injecting volinanserin to male mice of two to three months of age three hours after the beginning of the light phase of the light–dark cycle produced a significant increase in SWS during the first three hours after administration. This compound also caused reduction in W and REM sleep during the same period (Table 4). 35 Administration of volinanserin to rats six hours after the beginning of the dark phase significantly reduced the latency to sleep onset and W values relative to vehicle. In addition, the 5-HT2A receptor ligand induced significant increase in SWS and EEG delta power. REM sleep was not affected by the treatment (Table 4). 36
In vitro and in vivo pharmacological properties of volinanserin, pruvanserin, eplivanserin, nelotanserin, and pimavanserin.
The effects of drugs that inhibit the activity of serotonin 5-HT2A receptor on sleep variables in laboratory animals.
Pruvanserin
Pruvanserin, a phenetyl piperazine, structurally related to volinanserin also shows 5-HT2A receptor-blocking properties. It binds to 5-HT2A receptor with high affinity (Ki = 0.35 nM). In contrast, the Ki values of >1000 nM were described for serotonin 5-HT2C receptor. Pruvanserin prevented 5-HT-stimulated [35S]guanosine 5‘-0–3-thiotriphosphate accumulation in hamster ovary cells transfected with the human 5-HT2A receptor. Additionally, it antagonized N-ethoxycarbonyl-2-ethoxy-1,2-di-hydroquinoline-induced decrease in [3H]ketanserin binding in frontal cortex of the rat, and DOI-induced head-twitch behavior in mice (Table 3). 37 The effects of pruvanserin on sleep and W have been determined in the rat during both phases of the light–dark cycle. Injection of pruvanserin two hours after the beginning of the light phase significantly increased SWS and reduced REM sleep. Administration of pruvanserin two hours after the beginning of the dark period gave rise to a significant increase in SWS. Values corresponding to W, light sleep, and REM sleep were not significantly modified (Table 4). 38
Eplivanserin
Eplivanserin is a propenone ether derivative with potent 5-HT2A receptor blocking properties and a relatively long duration of action. It has high affinity for 5-HT2A receptor, moderate affinity for 5-HT2C receptor, and low affinity for 5-HT2B receptor. In vivo [3H]eplivanserin was found to bind predominantly in mouse brain regions containing 5-HT2A receptor, and this binding was displaced by the nonselective 5-HT2A/2C receptor antagonists ritanserin and ketanserin. In addition, the agent antagonized 5-hydroxytryptophan-induced head twitches in mice (Table 3). 39
The effects of eplivanserin administered three hours after the beginning of the light phase on the sleep–wake cycle were studied in male rats prepared for chronic sleep recordings. 40 Eplivanserin significantly increased the mean number and duration of SWS episodes (Table 4).
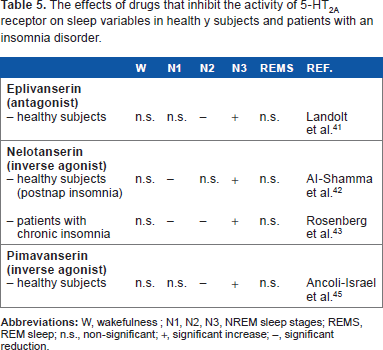
Landolt et al. 41 investigated the effects of eplivanserin (1 mg, p.o.) on sleep EEG and power spectra in young healthy men. Either eplivanserin or placebo was administered three hours prior to the beginning of the recording session according to a randomized, double-blind, crossover design. Eplivanserin induced a significant increase in N3 sleep, while N2 sleep was reduced. No significant differences between placebo and the 5-HT2A receptor antagonist were observed for SOL, REM sleep latency, TST, SE, and REM sleep in minutes (Table 5). NREM sleep power within 0.75–4.5 Hz was augmented, while that corresponding to spindle activity (12.5–15 Hz) was reduced. Subjective sleep quality was not affected by the compound.
The effects of drugs that inhibit the activity of 5-HT2A receptor on sleep variables in health y subjects and patients with an insomnia disorder.
Nelotanserin
Nelotanserin, a phenylpyrazole urea, is a potent inverse agonist of 5-HT2A receptor. Radioligand binding assays have shown that nelotanserin displays high affinity for human 5-HT2A receptor, while its affinity for 5-HT2C and 5-HT2B receptors is moderate and low, respectively. The Ki of the derivative for rat 5-HT2A receptor is sixfold greater than that of human 5-HT2A receptor. 42 In vitro experiments have shown that nelotanserin blocks 5-HT-induced inositol phosphate accumulation. During in vivo studies, the compound dose dependently prevented DOI-induced hypolocomotion in rats (Table 3). The acute effect of nelotanserin on sleep variables has been tested in rats, where the derivative was administered by oral route two hours before the beginning of the light phase. Nelotanserin induced a significant increase in SWS and EEG delta power, while W and REM sleep were reduced (Table 4). SOL remained unchanged. In addition, nelotanserin increased SWS consolidation as judged by the reduction in bouts number and increase in bouts duration. Subchronic administration of the 5-HT2A receptor inverse agonist two hours before lights on for five days induced an increase in SWS consolidation and in EEG delta power. Neither tolerance with repeated dosing nor rebound after dosing cessation was detected for any of the parameters quantified.30,42
Al-Shamma et al. 42 assessed the effects of nelotanserin (10, 20, and 40 mg, p.o.) in a postnap insomnia model in healthy subjects aged 18–45 years.
A significant increase in N3 sleep and reduction in N1 sleep was observed two to four hours after administration of the 40 mg dose (Table 5). SOL and TST were not affected by the treatment. The three doses of the drug significantly reduced the number of awakenings and of bouts of sleep, whereas the duration of bouts of sleep was augmented. The effects of nelotanserin have been evaluated also in patients with a diagnosis of chronic insomnia. The double-blind, placebo-controlled, three-way crossover study included patients with a mean age of 45 ± 11.8 years who received nelotanserin (10 and 40 mg, p.o.) for seven days. Both doses of nelotanserin induced a statistically significant increase in N3 sleep, whereas values corresponding to N1 and N2 sleep were diminished (Table 5). In addition, TST and SE were significantly increased at the beginning of the treatment following administration of the 40 mg dose. On the other hand, the number of awakenings and WASO were reduced by both doses of the derivative at both timepoints. 43
Pimavanserin
Pimavanserin is a 5-HT2A receptor inverse agonist that utilizes a urea core to bridge hydrophobic and quaternary amine moieties. Receptor selection and amplification technology have permitted a determination that pimavanserin is a potent inverse agonist at human 5-HT2A receptor. The inverse agonist potency of the compound is significantly less for 5-HT2C receptor and absent for 5-HT2B receptor. 44 Concerning the in vivo activity of pimavanserin, it has been shown to attenuate DOI-induced head twitches and to restore DOI-disrupted prepulse inhibition of acoustic startle response in rats (Table 3). 44 Ancoli-Israel et al. 45 evaluated the effects of pimavanserin in healthy adult volunteers in a randomized, placebo-controlled, double-blind study that included patients with a mean age of 51.8 ± 6.9 years. Pimavanserin (1, 2.5, 5, or 20 mg, p.o.) was administered in the morning for 13 consecutive days. The 20 mg doses of the derivative significantly increased N3 sleep and reduced N2 sleep along the administration period. The number of awakenings was significantly decreased on day 1 only. In contrast, SOL, WASO, TST, and REM sleep were not affected by pimavanserin (Table 5). With respect to spectral power density parameters, pimavanserin significantly increased slow delta, fast delta, and theta activities and reduced spindle frequency during NREM sleep. In addition, beta activity was diminished during REM sleep.
Summary of the Effect of the Inhibition of 5-HT2A Receptor Activity on Sleep
As presented in Table 4, SWS was consistently increased in laboratory animals irrespective of the inhibition of 5-HT2A receptor activity during the light period (when sleep predominates) or the dark period (when W predominates) of the light– dark cycle. In contrast, changes in W and REM sleep were inconsistent, which could be partly related to the amount of drug administered to the rodents.
With respect to the studies described in Table 5, it should be mentioned that irrespective of the inclusion of subjects with normal sleep, healthy subjects during a postnap insomnia, or patients with a chronic insomnia disorder, either the serotonin 5-HT2A receptor antagonist eplivanserin or the 5-HT2A receptor inverse agonists nelotanserin and pimavanserin significantly increased N3 sleep. Taking into account the high affinity and potency of these compounds for 5-HT2A receptor, it can be proposed that the increase in N3 sleep was dependent upon 5-HT2A receptor selective inhibition. In other words, the much lower affinity and potency of the 5-HT2A receptor antagonists and inverse agonists for 5-HT2C receptor tends to indicate that its contribution to the increase in N3 sleep was irrelevant. Our proposal applies also to the studies that included laboratory animals.
Effects of Coadministration of Eplivanserin and Zolpidem on Sleep Variables in Rats
Griebel et al. 40 characterized, in a preliminary study, the effects of eplivanserrin combined with zolpidem on sleep and W in rats. The isolated administration of 3 mg/kg of zolpidem, p.o., or 3 and 10 mg/kg of eplivanserin, p.o., was ineffective in this respect. In contrast, the combined administration of 3 mg/kg of eplivanserin, p.o., and 3 mg/kg of zolpidem, p.o., was followed by a significant decrease in W, while SWS and the mean duration of SWS bouts were augmented. It was concluded that the administration of subeffective doses of a 5-HT2A receptor antagonist with a short-acting non-BZD hypnotic induced a significant increase in SWS and a reduction in W in the rat.
Conclusion
This review focuses on chronic insomnia disorders, as characterized by difficulty in initiating sleep and/or difficulty in maintaining sleep. Compared to subjects with normal sleep, a considerable number of patients with an insomnia disorder show a significant reduction in SWS and REM sleep. Drugs approved for the treatment of a chronic insomnia disorder, including BZD receptor allosteric modulators (either BZD or non-BZD), ramelteon, low-dose doxepin, and suvorexant, are effective in treating the sleep initiation difficulties and/or in supporting the sleep maintenance. However, following treatment with these agents, N3 sleep does not revert to normal levels and indeed can be even further suppressed in more number of patients. This is an important issue, because N3 sleep reduction, particularly in patients with a chronic insomnia disorder or in elderly patients, could contribute to the deterioration of their day-to-day waking performance due to cognition and memory deficits, a phenomenon that is often exclusively attributed to independent neural or developmental factors. During the search for compounds that can increase N3 sleep, our attention was directed to two groups of drugs, namely the 5-HT2A receptor selective antagonists and inverse agonists. Currently available evidence tends to indicate that irrespective of their mechanism of action, isolated administration of eplivanserin and pimavanserin to subjects with normal sleep and nelotanserin to healthy volunteers during a postnap-induced insomnia significantly increased the duration of N3 sleep. A similar outcome was described when nelotanserin was given to patients with a chronic insomnia disorder. Interestingly, a preclinical study tends to indicate that the coadministration of subminimal doses of eplivanserin and zolpidem results in a significant increase in SWS and reduction in W in the rat. Thus, the association of a compound that inhibits the activity of 5-HT2A receptor with a hypnotic drug could be a valid alternative to improve the sleep induction and maintenance and, in addition, to normalize the N3 sleep values in patients with an insomnia disorder. It should be mentioned that up to the present time, none of the presently available selective 5-HT2A receptor antagonists and inverse agonists has been approved for clinical use. Safety concerns, including the incidence of side-effects, contraindications, and interactions, could have led to their failure to gain regulatory drug approvals.43,45 It is expected that new derivatives with a safer profile will become available in the near future.
Author Contributions
Conceived and designed the experiments: JMM. Analyzed the data: JMM. Wrote the first draft of the manuscript: JMM. Contributed to the writing of the manuscript: PT, SRP. Agree with manuscript results and conclusions: JMM, PT, SRP. Jointly developed the structure and arguments for the paper: JMM, PT, SRP. Made critical revisions and approved final version: JMM, PT, SRP. All authors reviewed and approved of the final manuscript.