
Abstract
Cancer therapies based on single target molecules have proved to be ineffective both in terms of their desired action and associated undesired side effects. Combinatorial cancer therapies involve selection of different components with targeted effects, which can lead to a synergistic effect for anticancer therapy. Curcumin induces the expression of p53 and downregulates that of Mdm2, ultimately resulting in induction of apoptosis. Subsequently, there is an elevated expression of p53-induced genes, which activate reactive oxygen species (ROS) thereby establishing cellular communication and disposition of any aberrant cell by growth arrest or apoptotic cell death. As a whole, the triad of curcumin, p53, and ROS presents a unique and promising solution to the designing of modern and patient-specific cancer therapeutics.
Introduction
Curcumin (diferuloylmethane; molecular formula: C21H20O6, melting point: 183 °C) is a polyphenol substance derived from plants. Curcumin is a rhizome of the plant Cucuma longha and is one of the major components found in turmeric (vernacular name – turmeric/haldi; Fig. 1). The common solvents of curcumin include ethanol, dimethyl sulfoxide, and other organic solvents; it is mostly insoluble in water (Fig. 1). Although it has a varied day-to-day utilization as a food spice, its medicinal significance has motivated several scientific and clinical researches. 1 A few of the known attributes of curcumin highlight it as a novel candidate for treating diseases such as arthritis, inflammation, multiple sclerosis, Alzheimer, and cancer. 1 Previous documented studies and preclinical literatures have shown that curcumin can potentially inhibit the tumor formation in the animal model of carcinogenesis.1,2 However, identification of its specific and shared molecular targets still remains elusive.
Keto form of curcumin.
In addition, not much is known about what constitutes an effective patient/client response to the curcumin-based anticancer therapy. A possible reason that has surfaced over these years involves differential gene expression and modulation of the existing genomic and proteomic constitution of the patient.
With curcumin exhibiting a wide range of pharmacological activities, the mainstream focus of cancer therapeutics has witnessed a translation from search for single-agent treatment to that of potent multimodal anticancer therapy.
Apoptosis or programmed cell death is vital in limiting cell growth and regulating cell cycle in order to maintain homeostasis. There are two major pathways (ie, extrinsic and intrinsic) that the programmed cell death cumulates within a cell leading to the activation of the aspartate-specific cysteine protease (caspases). 3
The extrinsic pathway involves engagement of death receptors that belong to the tumor necrosis factor (TNF) family, in the formation of the death inducing signaling complex. 4 A conditional rise in the expression of these death receptors (ie, TNF receptors and FAS receptors) on specific cells and further stable conjugation with their complementary ligands (TNF-alpha ligand, FAS ligand) lead to activation of caspases. The caspase-based induction of cellular death deploys both initiator/activator caspases (caspase-8 and -9) and executioner caspases (caspase-3, -6, and -7). 4
The intrinsic pathway is triggered in response to DNA damage (strand breaks, base dimerization, etc) 5 and is associated with mitochondrial depolarization and release of cytochrome c from the mitochondrial intermembrane space into the cytoplasm. Cytochrome c, apoptotic protease-activating factor-1, and procaspase-9 are activated which in turn promote the activation of caspase-3. 6 The process of DNA damage at the intrinsic level is often understood as an outcome of the negative effect of reactive oxygen species (ROS).
ROS and free radicals, such as hydroxyl radical and hydrogen peroxide, are produced in the body as by-products of several metabolic pathways 7 and upon exposure to exogenous stress, such as ionizing radiations, air pollution, or external stressors. 6 Several life processes involve the integration of multistepped redox pathways where ROS are recruited as secondary messengers. 7 However, within the body, the systemic response to the level of these radical species is headed by a well-equipped antioxidant defense mechanism (involving enzymes) that counteract the ROS. 8 Therefore, any failure in the activation or enhancement of these antioxidant defendants can lead to an elevated oxidative stress and in turn higher cellular damage and death. Therefore, we can correlate the role of ROS in the transformation of a normal cell into a tumor cell and its metastatic progression into cancerous cells.
One of the most ubiquitous physiological aberrations associated with cancer pathology is dysregulation of cell cycle and an uncontrolled event of cell proliferation. Tumor suppressor proteins occupy a pivotal position in maintaining genomic integrity. Therefore, our search for molecular regulators ended up highlighting p53 (tumor suppressor gene) 8 and its corresponding negative regulator, ie, Mdm2 (Mouse double minute 2 homolog – an inhibitor protein) as two such unique candidates.
p53, a proapoptotic gene, 9 is known to dispose a cell for programmed cell death (apoptosis) via a cascade of signaling pathways. On the other hand, Mdm2 exhibits a putative regulation of cell growth and death by altering the transformation of normal cells into tumor cells. Thus, for therapeutic innovations targeting cancer cell growth, it will be wise to explore this keynote interplay of apoptosis and cell cycle in the presence of curcumin.
Although the effect of Mdm2 on regulation of cell cycle has been elaborately characterized in the form of its p53 targets, p53-independent targets have been gaining momentum and are in focus very recently. 8 This review will summarize our current understanding of Mdm2- and p53-based regulation 9 of cell death via differential signals involving ROS-mediated pathways. 9 In addition, most of the available experimental data and literatures have ignored the interaction of curcumin with each of these molecules (p53 and Mdm2), and critically, how the interplay of these three can be an effective anticancer therapy needs more discussion and understanding.
Multimodal Action of p53
Sensation, altercation, and autoregulation
p53, a tumor suppressor protein, plays a key role in the regulation of several cellular processes, including the cell cycle, apoptosis, DNA repair, angiogenesis, and antioxidant defense mechanism. 10 The apoptotic function of p53 is critical for tumor suppression and reconstitution of inactive p53. 10 It actively agitates physiological response to cellular stress, such as hypoxia, DNA damage, and oncogene activation, and has the ability to eliminate excess, damaged, or infected cells by inducing apoptosis. Therefore, p53 is critical for proper regulation of cell proliferation in multicellular organisms.
Although its cellular level is low in homeostatic conditions, there is a significant rise in the level of p53 after sensation of any physiological stress condition. The p53 protein induced in stressed cells shuttles into the nucleus where its action as a transcription factor induces the expression of several downstream genes such as Bax (Bcl-2 associated X protein), GADD45 (the growth arrest and DNA damage 45 protein), and p21, 11 which largely come under the group of apoptosis-inducing and tumor growth-inhibiting genes. 12 It would be convincing enough to hypothesize that such a regulator of cellular growth must have an autoregulatory feedback loop, to regulate its own cellular levels.
And given the case that in normal cellular conditions p53 is expressed at low level, it justifies that its negative regulator protein must be upregulated in similar conditions. Therefore, in normal conditions, expression of p53 induces the expression of Mdm2 oncogene, which acts as a negative regulator of cellular levels of p53.
MDM2 Regulation of p53
From transcriptional inactivation to proteosomal degradation
MDM2, an oncoprotein, is coded by Mdm2 gene (which is Hdm2 in human beings) and acts as an E3 ubiquitin ligase. It is known to target p53 and thus commits it to proteosomal degradation (short-lived protein) in normal conditions. This action of Mdm2 requires its shuttling out of the nucleus via activation of nuclear export signal 12 and thereby causing the rise in the cytosolic levels of Mdm2. 13 During this cytosolic localization, there is a rapid decline in the level of p53. One such interesting experiment performed by Freedman et al exhibited the role of NES activation in Mdm2 and subsequent degradation of p53 in vivo even leading to low and steady levels of p53. 14
Apart from regulating p53 protein levels, Mdm2 also exerts an active inhibition of transcriptional levels of p53 by binding to its transactivation domain. Several papers have confirmed this inhibitory role of Mdm2 both in in vitro and in vivo assays. Moreover, Mdm2 interacts with several tumor suppressor proteins, including retinoblastoma, p21, p19/14ARF, E2F1 (E2F transcription factor 1), p73, and Mtb (Mycobacterium tuberculosis).13,14 These proteins constitute for p53-independent targets of Mdm2, which can also prove to be instrumental in our search for a potent anticancerous property of curcumin via regulation of Mdm2.
Anticancer Effects of Curcumin
Evidences and preliminary insights
Cancer drug development over the past decade has consolidated our focus on the modulation of specific targets, mostly one at a time (genes or proteins leading to dysregulation of cell growth and proliferation pathways). Emergence of the new generation of combinatorial and patient-specific cancer drug designing has led to the development of effective and targeted cancer therapeutics, where multiple carcinogenic modalities are under focus.
Indeed, cutting-edge molecular biology-based research has strengthened our claim for curcumin's role in the disruption or restriction of specific cancer-causing molecular mechanisms (transformation, proliferation, and metastasis). Furthermore, even in the case of drug-resistant tumor cell lines, the response has been positive to suppress the growth of tumor. 14
Curcumin exerts highly cell-specific and context-dependent regulation of cellular reproduction. In already existing tumors, curcumin tightly regulates the molecular signaling involved in cell cycle. This helps to enhance the cellular population of healthy control and stops the uncontrolled proliferation of new tissues.
One of the major advantages of curcumin-based anticancer therapy is its minimal side effect. This can be summarized as negligible off-target effects of curcumin along with noninvasiveness toward neighboring healthy tissues. 15 It is also known to destroy tumor growth by making these tumors more susceptible to pharmacologic cell-killing treatments. 16 In addition, curcumin modulates tumor suppressor pathways by triggering mitochondrial-mediated cell death in tumor tissues and thereby increasing the death of cancer cells. 17 In its multimodal approach, curcumin results in the starvation of tumors of their vital blood supply by blocking angiogenesis and vasculogenesis. 18 It has proved to be effective in opposing many of the processes that permits the spread of metastatic cancer cells. These multitargeted actions are central to the capacity of curcumin to block multiple forms of cancer by targeting different stages of tumor growth and cellular plasticity.16,17
Curcumin inhibits interaction of MDM2 and p53
Curcumin binds to Mdm2 and leads to the loss of hold on p53. As a consequence, p53 that was otherwise held inactive by Mdm2 (ubiquitn ligase protein) gets reactivated and translocates into the nucleus (Fig. 2). 18 As a result, specific pathways such as cell cycle repair, antioxidant defense mechanism, and other apoptotic pathways are called into action (Fig. 2) and cellular homeostasis is restored. Several experiments of Lin et al over the past decade have proved the correlation of curcumin with cancer inhibition in different clinical models of cancer. One important contribution of the group has been their work on proposing curcumin as a dietary supplement for combating cancer and thus has been of immense promise to the scientific community. However, most of these effects have been characterized in clinical models of cancer and in in vitro experiments where the exact role play of molecular modulators have been largely ignored. 19 Taking this cue, it can be hypothesized that curcumin can have much better and finer chemopreventive and chemotherapeutic impressions than that earlier reported in several experiments conducted by Li et al (Fig. 2).
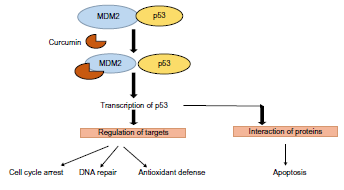
Inhibitory effect of curcumin on Mouse double minute 2 homolog (MDM2) and p53 interaction. Curcumin binds to MDM2 and results in the disassociation of p53 and downstream transcriptional regulation of target gene expression. The disassociated p53 also interacts with other proteins and heralds programmed cell death (apoptosis).
Literature review suggests that multiple experimental approaches and therapeutic paradigms have established curcumin as a chemotherapeutic agent. One such study involves the designing of a nanoparticle-based drug delivery system for curcumin at the site of specific cancer cells. In addition, several efforts are ongoing to improve the efficacy of curcumin by developing derivatives of curcumin, eg, Pculin02H. 20
However, much recently, attention has been shifted in understanding the molecular regulation of this process. Curcumin negatively regulates MDM2 protein resulting in destabilization of the interaction of Mdm2 with p53 (Fig. 3). 21 The then released p53 tries to repair DNA damage by activation of p21 and 14-3-3 and subsequent arrest of cell cycle at G1 and G2 22 check points, respectively (Fig. 3). 23 As a result, there is cellular induction of apoptosis. Interestingly, the exact mechanism driving this p53 guided cell cycle arrest or apoptosis is only partially understood and is a subject of further study.
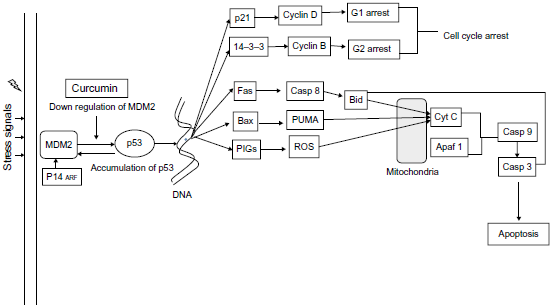
Curcumin downregulates MDM2 and thereby upregulates the transcriptional influence of p53 on several target genes. One line of action involves the transcriptional activation of p21 and 14-3-3 expression, which leads to cell cycle arrest. in addition, it also upregulates proapoptotic genes such as Fas, Bax, and p53-induced genes (PIGs), which lead to activation of caspases and ultimately end with cellular apoptosis.
Various factors that are known to influence this decision of cell cycle arrest or apoptosis include the level of p53 expression (Fig. 3), the nature and type of stress signal, the cell type, and the cellular state when under stress. 23
Curcumin-induced extrinsic apoptotic pathway is mediated by death receptors like Fas and TNF, which involves cleavage of BH3-interacting domain death agonist (BID) 24 by activated caspase-8 (initiator caspase; Fig. 3). This results in the release of cytochrome c 25 from the mitochondria, and subsequent activation of caspase-3 (executioner caspase) resulting in apoptosis. 25 However, the role of curcumin in mitochondrial regulation of antiapoptotic proteins such as B-cell lymphoma-2 (Bcl-2) or B-cell lymphoma-extra large (Bcl-xl) is still unresolved and needs more investigation (Fig. 3).
Curcumin and inflammation
Curcumin blunts cancer-causing inflammation and thereby reduces the level of inflammatory cytokines throughout. This is achieved by blocking the inflammatory master molecule nuclear factor-kappaB (NF-κb). 26 Curcumin inhibits a crucial inflammation protein called NF-κb, which ultimately reduces the site-specific onset of allergic reactions. 25
Most anti-inflammatory drugs try to prevent the onset of inflammatory reactions by inhibiting specific inflammatory enzymes. Curcumin is known to work effectively against cyclooxygenase-2 (COX-2) and lipoxygenase (LOX), which are molecular modulators of cellular inflammation and lead to DNA damage. 26 Regular intake of curcumin is known to significantly reduce the levels of inflammation-triggering enzymes. In human beings, a dosage of 200 mg/kg of body weight of curcumin can prevent hyperallergic inflammations. 27
Curcumin downregulates the production of dangerous and advanced glycation end products that could trigger inflammation otherwise leading to cancerous mutations. 27
The Complex triad: Curcumin, ROS, and p53
In response to cellular stress resulting from accumulated ROS, extensive DNA damage is a common outcome. Although the wild-type p53 orchestrates transcription of numerous genes and decides the fate of these stressed cells, the broad outcome of stressed cells includes arrest of cell cycle, senescence, and apoptosis. The role of curcumin in regulating biological redox system seems complex and paradoxical. Substantial literature based on in vivo and in vitro experiments have established curcumin both as an antioxidant and a prooxidant. 28 Ironically, curcumin scavenges ROS and counterbalances endogenous redox system, whereas it is also known to incite cellular ROS production (Fig. 4). 29
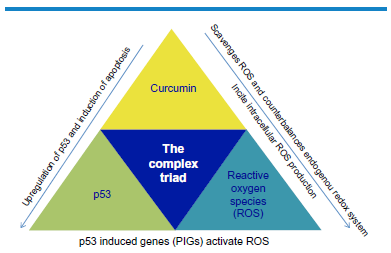
The complex triad of curcumin, p53, and ROS: exhibiting the complex interlinking relationship between the three key regulator molecules in the modulation of cancer cells and their microenvironment.
However, in noncancerous cells exposed to curcumin, with an increase in ROS (redox activation), 28 there is a concomitant upregulation of p53 and, ultimately, resulting in increased apoptosis. Similar association of ROS and curcumin-induced apoptosis in malignant cells was reported by Yoshino et al. 30 Therefore, in our search for an effective anticancer therapy, we have developed a broad understanding of curcumin, p53, and ROS interactions and have named these three interlinked components as “the complex triad (TCT)” (Fig. 4). The word complex is to emphasize the nondirect and mediator involved in the interaction of these molecules where it is still hazy to decipher, if each of these molecules exerts a feedback regulation on the other.
Oxidative stress has been correlated with different cell state directed cell rewards, such as cell cycle arrest, DNA repair, and apoptosis (Fig. 4). The excessive generation of ROS in mitochondria resulting from the treatment with chemotherapeutic agents lead to apoptosis, while oxidative stress in the nucleus directs cell to p53-dependent DNA repair pathway. 31
During several biological processes involving redox reactions, PIGs (Fig. 3) activate ROS, 31 which as a secondary messenger molecule moves into the mitochondria and interacts with the resident signaling molecules (Fig. 4). Such an interaction results in imbalanced mitochondrial membrane potential and thus cytochrome c gets released from the mitochondria. 32 Discrete response patterns advice that multiple biological pathways exist, which lead to an overall integration of p53 signaling (Fig. 5).
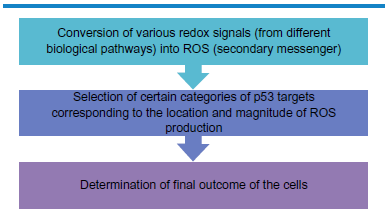
Discrete response of ROS and p53 on cells depends on several reasons. The cascade involves conversion of redox signals originating from different bioprocesses into ROS. Depending on the level of cellular ROS and its localization, specific targets for p53 transcriptional modulation are regulated (PIGs). This whole process determines the final outcome of the cells.
Annotation of Multiple Molecular targets of Curcumin
Not surprisingly, tremendous ongoing research on cancer over the past few years have established that carcinogenesis is a multistep process involving abnormal functioning or dysregulation of varied molecular regulators. These include that of the growth factors, growth factor receptors, transcription factors, cytokines, apoptosis, and proliferation genes. 33
Precisely, it is an overall loss of the cellular homeostasis influenced by both genetic and environmental makeup of a person. As a whole, this leads to the cellular onset of cancer growth and clinical manifestation in the form of metastasis.
Some specific molecules are described here for better understanding of potent with targets of curcumin and how they can be incorporated in our search for developing a curcumin based anticancer therapy. 34 This paper aims to reinforce our present understanding of the role of curcumin as a potent anticancer drug via these target molecules. 19
Of the several molecular targets of curcumin, Cyclin D, Akt, and NF-κb are major regulators of cell growth and differentiation, thereby determining the process of cell cycle and survivability.
The epithelial growth factor receptor and fibroblast growth factor-mediated signaling of phosphatidylinositol-3-kinase (PI3K)-Akt pathway is a major cell survival pathway. The dysregulation of this pathway has been implicated in several cases of tumorigenesis and significantly disturbed in metastatic cancers. Reports have linked the concentration and time of exposure-dependent interaction of curcumin with that of PI3K/Akt pathway, where the net outcome is the inhibition of Akt pathway.
At molecular levels, this inhibition is known to be achieved by limiting the phosphorylation of mammalian target of rapamycin (mTOR), Akt, and downstream substrates involved in the Akt pathway. An elaborate study by Yu et al highlighted a possible restoration of this negative effect of curcumin by upregulating Akt or siRNA-mediated gene silencing of tuberous sclerosis protein 1 (TSC1) and tuberous sclerosis protein 2 (TSC2; Fig. 6). 35 TSC1 and TSC2 are the major components of mTOR signaling pathway and are putative sites for modulation of cancer cells. 35
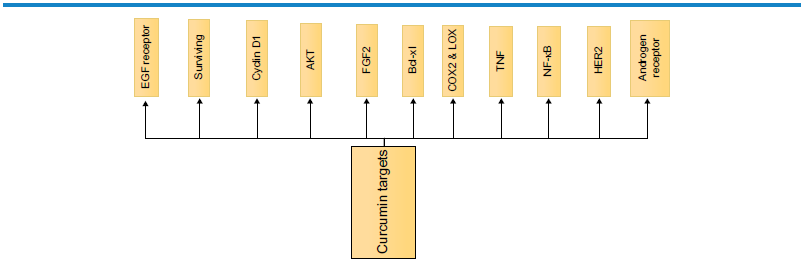
Common molecular targets regulated by curcumin directly or indirectly.
COX-2 and LOX are the major molecular regulators of cellular inflammatory reactions, and curcumin inhibits inflammation by downregulating the expression of these molecules (Fig. 6). These inflammatory reactions otherwise could have resulted in cellular damage and intrinsic damage of DNA leading to cancer progression. 19
TNF and Bcl (B-cell lymphoma) group of proteins constitute proapoptotic proteins, which activate caspases (activator and executioner caspases; Fig. 6) leading to onset of apoptosis. 35
Herceptin 2 and androgen receptors are commonly correlated with the onset of breast cancer and therefore are active targets of curcumin. This makes curcumin an effective therapeutic target for combating breast cancer (Fig. 6). Pculin02H, a curcumin derivative, has been effective in inhibiting proliferation than that of curcumin and thus highlights the development of curcumin derivatives, which can potentially target the growth of cancer cells. 20
Conclusion
Curcumin and p53 have long been known as anticancerous agents. MDM2 (primary cellular inhibitor of p53 activity) was found to hinder p53 through direct protein–protein interaction, ie, Mdm2–p53 and regulation during p53 transcription. We suggested that for this particular protein–protein interaction, it may be feasible to design a potent, nonpeptide, drug-like small-molecule inhibitor using curcumin as one of the key components to block the p53–MDM2 interaction. It is generally considered that the ROS may promote either cell proliferation or cell death activity depending on the intensity/ location of the oxidative collapse and the activity of the antioxidant system. The excess generation of ROS in mitochondria results from the treatment with chemotherapeutic agents giving rise to apoptosis, while oxidative stress in the nucleus directs cells to p53-dependent DNA repair.
However, limitation in understanding the molecular basis of this regulation has impacted on its transition from theory into practice. Other than traditional medicine in a few countries, the modern medicine has largely ignored this anticancerous drug in its mainstream cancer treatment strategies. In addition, the molecular interactions of curcumin, p53, and MDM2 have also opened up avenues for drug designing and development. Targeting this co-interaction model with significant inhibition on cancer will require further characterization of these therapies that can be developed into a vaccine or an adoptive cell transfer method to fight against cancer. Therefore, understanding the molecular basis of curcumin's action on cancerous cells will lead to newer studies as to how these can be incorporated to the conventional treatment and pharmacological formulations.
Author Contributions
Prepared the first draft of the paper: RKS. Developed the concepts, redrafted the paper and developed the concept of complex triad: SS. Helped in developing illustrations and figures, contributed to the discussion and conclusion section of the paper: CK, K. All authors reviewed and approved the final version of the manuscript.