
Abstract
Objective
The aim of this study was to assess the activity of cortisol-metabolizing enzymes in women with polycystic ovary syndrome (PCOS), using a fully quantitative gas chromatography/mass spectrometry (GCMS) method.
Design
We investigated the glucocorticoid degradation pathways that include llβ-hydroxysteroid dehydrogenase (llβ-HSD) type 1, 5α-reductase (5α-R) and 5β-reductase (5β-R), 3α-hydroxysteroid dehydrogenase, and 20α- and 20β-hydroxysteroid dehydrogenase (20α-HSD and 20β-HSD, respectively) in young nonobese women with PCOS, using a fully quantitative GCMS method.
Setting
This study was conducted in a tertiary referral hospital in Israel.
Patients
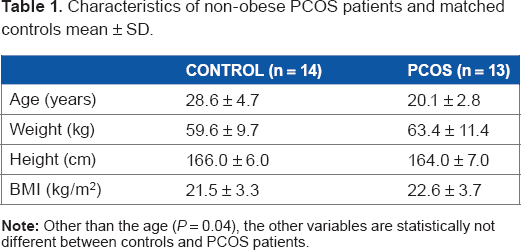
This study group consisted of 13 young women, aged 20.1 ±2.8 years (mean ± SD), with the body mass index (BMI) of 22.6 ± 3.7 kg/m2, diagnosed with PCOS according to the Rotterdam criteria. The control group consisted of 14 healthy young women matched for weight, height, and BMI.
Interventions
Urine samples were analyzed using GCMS. We measured urinary steroid metabolites that represent the products and substrates of the study enzymes and calculated the product/substrate ratios to represent enzyme activity.
Main Outcome Measures
The calculation of enzymatic activity, based on glucocorticoid degradation metabolites, was done by GCMS in PCOS vs. controls.
Results
All glucocorticoid degradation metabolites were higher in the PCOS group than in controls. Of the adrenal enzymes, the activities of 21-hydroxylase and 17α-hydroxylase were reduced, whereas the activity of 17,20-lyase was enhanced in PCOS. Of the degradation enzymes, the activity of 11β-HSD type 1 was reduced in women with PCOS only when calculated from cortoles and cortolones ratios. The activities of 5α-R/5β-R were increased only when calculating the 11-hydroxy metabolites of androgens. The activity of 20α-HSD was elevated in the patients with PCOS and its relation with the substrate levels was lost.
Conclusions
We confirm PCOS association with low 21-hydroxylase activity. PCOS is associated with dysregulation in glucocorticoid degradation. The activity of 5α-R is enhanced only through the backdoor pathway. Marked increase in the activity of 20α-HSD suggests a hitherto unknown derangement in PCOS.
Introduction
The polycystic ovary syndrome (PCOS) has been the subject of extensive research for the past several decades and has been shown to involve a widespread dysregulation of steroid metabolism. Hyperandrogenism in both lean and obese women is mostly of ovarian origin, but 20-30% of patients with PCOS also have adrenal hyperandrogenism.1–4 moreover, patients with PCOS demonstrate a generalized hypersecretion of adrenocortical products, basally and in response to adrenocor-ticotrophic hormone (ACTH) stimulation.3,4
One proposed contributing mechanism to these changes is an altered prereceptor Cortisol metabolism. Impaired function of llβ-hydroxysteroid dehydrogenase (11β-HSD) type 1 has been recorded in women with PCOS5,6 and has been suggested to contribute to the hyperandrogenic phenotype and the metabolic syndrome that are common in this disorder. This could occur through a mechanism of decreased Cortisol negative feedback and an exaggerated ACTH response, causing adrenal hyperandrogenism.4–8
However, little attention has been devoted to the degradation of glucocorticoids as a possible mechanism of hypercortisolism. This involves three major steps mediated by distinct enzymes. First, 5α-reductase (5α-R) and 5β-reductase (5β-R) in the glucocorticoid context irreversibly reduce Cortisol and cortisone into their dihydro metabolites 5α/β dihydrocortisol (DHF) and 5α/β dihydrocortisone (DHE), respectively. The activity of 5α-R in its androgenic role has been reported to be enhanced in women with PCOS.9–12 Second, 3α-hydroxysteroid dehydrogenase (3α-HSD) further reduces DHF, α-dihydrocortisol (αDHF), and DHE into tetrahydrocortisol (THF), α-tetrahydrocortisol (αTHF), and tetrahydrocortisone (THE), respectively. This enzyme has been suggested to play a role in hirsutism.
13
The third and final step involves 20α- and 20β-hydroxysteroid dehydrogenases (20α-HSD and 20β-HSD, respectively), which reduce DHF, αDHF, THF, αTHF, and THE into 20α-DHF and 20β-DHF, and α cortol (α-C) and β cortol (β-C) into α cortolone (α-CL) and β cortolone (β-CL), respectively. The cortols are metabolites of THF after degradation by the enzymes 20α-HSD and 20β-HSD, and the cortolones are metabolites of THE after the action of the same enzymes, as clearly shown in Figure 1. A small increase in the activities of 20α-HSD and 20β-HSD and their expressions in subcutaneous adipose tissues were suggested in women with PCOS.
14
Diagram showing the degradation metabolites of Cortisol and cortisone, explaining the formation of cortols and cortolones.
The present study was designed to assess the activity of adrenal and cortisol-metabolizing enzymes in women with PCOS, using a fully quantitative gas chromatography/mass spectrometry (GCMS) method.
Subjects and Methods
Subjects
Characteristics of non-obese PCOS patients and matched controls mean ± SD.
Methods
To reflect peak diurnal levels, urine sample was taken on the second morning from each subject during the follicular phase of the menstrual cycle (days 3-7). In women with PCOS who had no ovulations or menstruation, the urine sample was taken within two weeks of an induced withdrawal bleeding. This study was approved by the institutional review board committee and conducted in accordance with the principles of the Declaration of Helsinki. Patients signed informed consent.
Gas chromatography/mass spectrometry
Urinary steroid profiles were determined from spot urine specimens, as reported by a previous study. 17 An aliquot of urine that contained 6-μmol creatinine was processed by solid phase extraction, hydrolysis, reextraction, and derivatization. It was then purified by gel chromatography. Gas chromatography was performed using an Optima-1 fused column, and helium was used as a carrier gas at a flow rate of 1 mL/minute. The gas chromatograph (Agilent 6890) was directly interfaced with a mass selective detector (Agilent 5972A), operated in selected ion monitoring mode. For a fully quantitative analysis, calibration curves were created for each of the 31 metabolites.
Statistical analyses
Results were presented as mean ± SD. The statistical significance of comparisons was assessed using the two-tailed Student's i-test, and correlations were tested by the Pearson correlation coefficients, with a significance defined as P < 0.05.
Results
Adrenal enzyme activity as calculated from the product/substrate ratios.

steroid metabolizing enzyme activity as calculated from the product/substrate ratios.

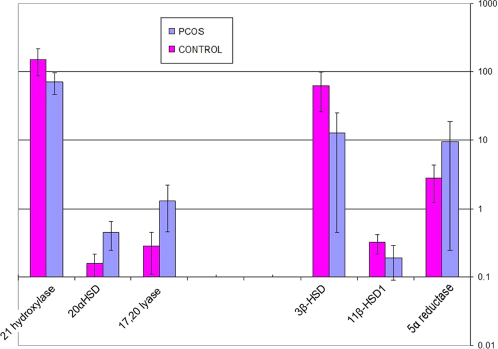
Enzyme activity–-PCOS patients vs. controls (mean ± SD).

urinary mineralocorticoids in PCOS patients and controls (mean ± sD).
Discussion
The present results confirm previous contention as to enhanced glucocorticoid levels in PCOS and low 21-hydroxylase activity, and add 17-hydroxylase deficiency and 17,20-lyase enhancement to the steroid abnormalities signature of the syndrome. Here, we add a set of deranged steroid degradation enzyme activities to the disease signature and show that nonobese PCOS subjects have enhanced activity of 20α-HSD, a forgotten enzyme that may have a function, which is yet unknown.
Previous reports demonstrated enhanced 5α-R activity,9–12 which we hereby confirm to a mild degree (P = 0.044) with regard to its backdoor pathway,18,19 expressed as BMI-dependent high ratio of 11OH-An/11-OH-Et, but not as measured for its mineralocorticoid degradation αTHB/ THB or its glucocorticoid degradation αTHF/THF. The backdoor pathway is an alternative route to dihydrotestoster-one (DHT) synthesis, which bypasses androstenedione and testosterone intermediates by the 5α-reduction of progesterone or 17OHP into the preandrogen metabolites. The activity of the backdoor pathway was assessed using the ratios of the urinary concentrations of pregnanediol to those of the metabolites of the classic 4 and 5 pathways. In contrast to etiocholanolone, which originates almost exclusively from the classic pathways, androsterone may be derived additionally from the backdoor pathway. Therefore, the androsterone-to-etiocholanolone ratio can be used as an indicator for the presence of the backdoor pathway.
These ratios are further catalyzed in androgen target tissues into DHT–-the principal virilizing hormone. 18 This pathway was originally described in tammar wallaby18,20 and in rodents. 21 Human beings have the complete enzymatic cascade for the backdoor pathway to DHT, 18 and its activity was shown to be enhanced in congenital adrenal hyperplasia (CAH) due to 21-hydroxylase deficiency. 19 This pathway, which catalyzes an abundance of androsterone, might explain how potent androgens are produced in PCOS, while the conventional androgen biosynthetic pathway remains unaffected.
The present results confirm the previously reported decrease in 11β-HSD type 1 activity in PCOS.5–8 When interpreting the results for urinary steroid metabolites, the intermediate tetrahydro metabolites are to be accounted for. When using the metabolite ratio (THF + 5α-THF)/THE, the difference in activity was insignificant. It became significant when calculating the 20α-hydroxy metabolites. Two studies have previously reported a weak increase in the activities of 20α-HSD and 20β-HSD in lean women with PCOS.6,22 When separately analyzing the activities of 20α-HSD and 20β-HSD, we found a highly significant increase in the former and a normal activity in the latter in the PCOS group. The 20α-aldo/keto reductase is a member of chromosome 10p15-p14 AKR1C1 gene superfamily, which consists of more than 40 known enzymes and proteins. These enzymes catalyze the conversion of various aldehydes and ketones to their corresponding alcohols by utilizing NADH and/or NADPH as cofactors. The enzymes display overlapping but distinct substrate specificity. The enzyme 20α-HSD has been investigated in rodents and human beings as an important ovarian enzyme in progesterone metabolism.23–25 It is expressed in human endometrium and decidua, catalyzes the conversion of progesterone to its inactive form 20α-dihydroprogesterone, and thus is thought to play a role in pregnancy and parturition. Mice deficient of 20α-HSD sustain high progesterone levels and display a delay of several days in parturition, suggesting that this enzyme regulates parturition in an essential and nonredundant manner. 24 It was suggested that the roles of human AKR1C are for the hepatic formation of 5β-tetrahydrosteroids of progesterone. 26 Urinary steroid metabolite analysis using GCMS is an established confirmatory diagnostic tool.12,19,27 The 20α-HSD excess has not been investigated, and therefore, attributing to it a role in the PCOS suppression of luteal progesterone generation and menstrual dysregulation remains speculative. The physiological role of 20α-HSD in glucocorticoid metabolism is unknown, but its expression in the mouse's X zone of the adrenal gland may suggest such a role. 23
Author Contributions
Conceived and designed the experiments: ZB, GK, NZL, ED, CK, ZH. Analyzed the data: ZB, GK, NZL, ED, CK, ZH. Wrote the first draft of the manuscript: ZB, GK, NZL, ED, CK, ZH. Contributed to the writing of the manuscript: ZB, GK, NZL, ED, CK, ZH. Agree with manuscript results and conclusions: ZB, GK, NZL, ED, CK, ZH. Jointly developed the structure and arguments for the paper: ZB, GK, NZL, ED, CK, ZH. Made critical revisions and approved final version: ZB, GK, NZL, ED, CK, ZH. All authors reviewed and approved of the final manuscript.