
Abstract
Background
In neonates, the increase in O2-delivery (DO2) by dopamine is offset by a greater increase in O2-consumption (VO2). This has been attributed to β3-adrenergic receptors in neonatal brown fat tissue. β3 receptors in the heart have negative inotropic properties. We evaluated the effects of SR59230A, a β3-antagonist, on the balance of systemic and myocardial O2-transport in newborn lambs treated with dopamine.
Methods
Lambs (2-5 days old, n = 12) were anesthetized and mechanically ventilated. Heart rate (HR) and rectal temperature were monitored. VO2 was measured by respiratory mass spectrometry and cardiac output (CO) by a pulmonary artery transonic flowmeter. Arterial, jugular bulb venous and coronary sinus blood gases and lactate were measured to calculate DO2, O2 extraction ratio (ERO2), myocardial O2 and lactate extraction ratios (mERO2, mERlac). After baseline measurements, lambs were randomized to receive SR59230A at 5 mg/kg iv (SRG) or placebo. Both groups received incremental doses of a dopamine infusion (0-5-10-15-20 mcg/kg/min) every 15 min. Measurements were repeated at the end of each dose.
Results
After SR59230A infusion, CO and HR trended to decrease (P = 0.06), but no significant changes occurred in other parameters. Over the incremental doses of dopamine, temperature increased in both groups (P < 0.0001) but to a lesser degree in SRG (P = 0.004). CO and HR increased (P = 0.005 and 0.04) and similarly in both groups (P > 0.1). DO2 trended to a small increase (P = 0.08). VO2 increased in both groups (P < 0.0001) but to a lesser degree in SRG (P < 0.0001). As a result, ERO2 increased in both groups (P < 0.0001), but to a lesser degree in SRG (P < 0.0001). mERO2 was lower in SRG (P = 0.01) with a faster increase (P < 0.0001). mERlac was higher in SRG (P = 0.06) with a faster decrease (P = 0.04).
Conclusion
Although SR59230A tends to induce an initial drop in CO, it significantly attenuates the rise in VO2 and hence the imbalance of systemic and myocardial O2 transport induced by dopamine at higher doses. Studies are warranted to examine the effect of SR59230A in cases of cardiac dysfunction and increased VO2, observed after cardiac surgery.
Introduction
Dopamine, a precursor in the endogenous synthesis of norepinephrine, is the most commonly used medication to provide cardiovascular support in critically ill patients with low cardiac output state, via the stimulation of β1 and β2 adrenergic receptors.1,2 Often, these patients have impaired balance of oxygen transport and poor tissue perfusion. Therefore, in addition to improved cardiovascular state, the major goal of therapy in these patients is to improve the balance of systemic oxygen transport and tissue oxygenation by increasing systemic O2 delivery (DO2) relative to changes in systemic O2 consumption (VO2). In adult humans and animal experiments, the increase in DO2 induced by catecholamines is about 10-fold of the increase in VO2.3–5
However, in neonates, in contrast to that observed in adults and older children, dopamine worsens the balance of systemic oxygen transport by a greater increase in VO2 than DO2 as shown after cardiopulmonary bypass surgery. 6 In neonatal lambs, dobutamine, a synthetic analog of dopamine, increases VO2 by 7-12 fold compared to older lambs, with a similar increase in DO2. The substantial increase in VO2 has been attributed to the abundant brown adipose tissue in neonates, which contains plentiful α, β1 and β3 adrenergic receptors.7,8 Stimulation of these receptors by catecholamines leads to non-shivering thermogenesis and a substantial increase in VO2.7,9,10 It has been shown that selective α1, β1 or β2-adrenoceptor blockade in newborn lambs does not affect the increases in VO2 or DO2 induced by dobutamine, but the combined adrenoceptor blockades markedly attenuated both VO2 and DO2. 3
β3-adrenergic receptor was discovered in 1970s and 1980s from pharmacological studies of rat adipocyte function, and is now known to be richly present in brown adipose tissue controlling of thermogenesis and energy balance.8,11,12 It has been shown that dopamine exerts a thermogenic effect on the brown adipose tissue and increases VO2 in rats. 10 Selective β3-adrenoceptor antagonist, SR59230A, 13 has been reported to down-regulate uncoupling protein-1 and norepinephrine induced cAMP accumulation,13,14 thus reduce thermogenesis in brown fat tissue in lambs. 15 In addition, it has been found that β3-adrenergic receptors in the heart have negative inotropic properties. This has led to research investigating potential therapeutic applications to enhance cardiac contractility by blocking β3-adrenergic receptors in heart failure. 16
Therefore, we hypothesized that SR59230A may attenuate the increase in VO2 induced by dopamine, and maintain the inotropic effect, thus cardiac output (CO) and DO2, thereby improving the balance of systemic and myocardial oxygen transport in neonatal lambs.
Methods and Materials
Animal Preparation
All experiments were conducted in accordance with guidelines and by approval of the Animal Care and Use Committee (Health Science), University of Alberta. Twelve mixed-breed newborn lambs 1 to 5 days of age weighing 2.5 to 5.0 kg were obtained from a local farm. Anesthesia was induced with isoflurane (2%-3%). After induction, isoflurane inhalation was ceased, and anesthesia was maintained with continuous i.v. infusion of fentanyl (5-10 μg/kg/h), midazolam (0.1-0.2 mg/kg/h) and pancuronium (0.05-0.1 mg/kg/h). Animals were intubated with a cuffed endotracheal tube to prevent any air leaks, and ventilated with FiO2 0.3 and volume-cycled ventilation (SERVO VENTILATOR 300, Siemens Medical Systems, Solna, Sweden), a tidal volume of 10 mL/kg, rates of 20-30 breaths/min, and PEEP of 4 cm H2O. Systemic arterial blood gases were measured (iStat, Abbott Laboratories. Abbott Park, IL USA) and ventilation was adjusted to maintain pH 7.35-7.45, PaO2 100-140 mmHg, and PaCO2 at 35-40 mmHg, while base deficits >2 mmol/L were corrected with 22 mEq/L NaHCO3. Blood that had been collected from an adult donor lamb was used for blood transfusion at a rate of 20 to 30 mL/h.
Left femoral arterial, superior vena cava, coronary sinus catheters were inserted to monitor pressures, blood gases and to administer fluids and medications. Heart rate was measured (Hewlett Packard, Palo Alto, CA). Maintenance fluids consisted of 5% dextrosesaline at 10 mL/kg/h. Temperature was monitored by a rectal thermometer, and maintained above 37 °C by a heating pad and warm blankets, which were kept constant during the experimental period. A sternotomy was performed and the ductus arteriosus was ligated. An 8-10 mm diameter ultrasonic perivascular flow probe (Transonics Systems, Ithaca, New York, USA) was placed around the main pulmonary artery to continuously measure CO.
VO2 was continuously measured using respiratory mass spectrometry (AMIS2000 Innovision A/S, Odense, Denmark). This is highly sensitive, accurate and rapid method to allow simultaneous measurements of multiple gas fractions, and has been described previously. 17 Stroke volume was calculated as CO/heart rate. DO2 was calculated as CO times arterial oxygen content (CaO2). Oxygen extraction ratio (ERO2) was calculated as VO2/DO2. All systemic hemodynamic and oxygen transport variables were indexed by weight as appropriate.
Myocardial oxygen and lactate extraction ratios (mERO2, mERlac, respectively) were calculated using standard equations:
mEO2 = (CaO2 - CcsO2)/CaO2
mERlac = (arterial lactate concentration - coronary sinus lactate concentration)/arterial lactate concentration
where CcsO2 indicates coronary sinus oxygen content.
experimental protocol
Lambs were randomized into two groups (n = 6 each) after a period of stabilization and baseline measurements of hemodynamics, oxygen transport and central temperature were obtained. Subsequently, the experimental (SR group) received an i.v. infusion of SR59230A at 5 mg/kg dissolved firstly in 0.5 mL DMSO, and then in 10 mL saline and an equal volume of the DMSO and saline, respectively, over 5 min. After 30 min, incremental dopamine i.v. infusions starting from 5 were increased to 10, 15, and 20 μg/kg/min sequentially every 15 minutes. Measurements were repeated before dopamine infusion and at the end of each dose of dopamine in both groups. At the end of the study, an over dose of phenobarbital was given for euthanasia.
Statistical Analysis
Data are expressed as mean ± SD. Mixed linear regression for repeated measures was used to compare the variables before and after SR59230A or saline infusion between the two groups. It was also used to compare the different levels of the variables and their trends of changes between groups over the incremental increases of dopamine, with analyses of the effects of group (Pgroup) and dose (Pdose) and the interaction between dose and group (Pggroup*dose). All data analysis was performed using SAS statistical software version 9.2 (SAS Institute Inc). A P value <0.05 indicates a statistical significance.
Results
There were no significant differences in age (mean 3 days old) and weight (4.0 ± 1.0 kg vs. 3.7 ± 0.6 kg) between the SR and Control groups, respectively.
Changes of the variables before and after SR 59230A/saline infusion in the two groups (Table 1 and Fig. 1)
All hemodynamic and oxygen transport variables were not significantly different at baseline between the two groups. Thirty minutes following SR59230A infusion, temperature and mean arterial pressure remained unchanged and were not different from those of Control group. Heart rate trended to decrease from 237 ± 33 beats/min to 202 ± 7 beats/min in SR group (P = 0.08). CO trended to decrease from 167 ± 31 ml/min/kg to 123 ± 8 mL/min/kg in SR group (P = 0.08). Stroke volume remained similar after SR59230A infusion within both groups. DO2, VO2, ERO2, mERO2 and mERlac did not change significantly in SR group. There were no significant changes in all the variables in Control group.
Mean ± SD values of temperature, hemodynamics, and oxygen transport during the study protocol.

P = 0.08 compared to Pre-SR values
Pdose < 0.05 for the change over incremental doses of dopamine
Pgroup*dose < 0.05 for the interaction between group and dose of dopamine, indicating the difference in the rate of change over incremental doses of dopamine between the two groups.
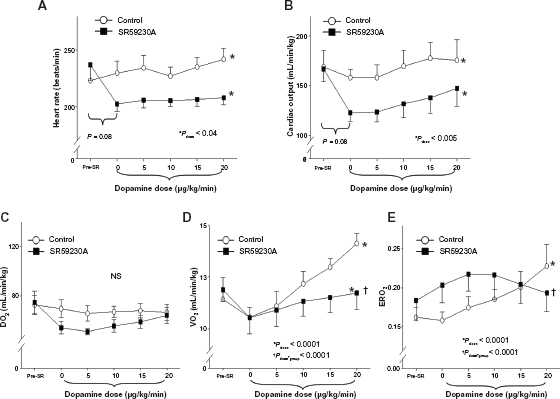
The changes of (
Changes of the variables over the incremental infusion of dopamine in the two groups (Table 1 and Fig. 1)
Over incremental doses of dopamine, temperature increased significantly in both groups (Pdose < 0.0001); the rate of temperature increase was significantly less in SR group compared to Control group (Pdose*group < 0.005). There was no significant change in mean arterial pressure in either SR or Control group. Heart rate significantly increased in both groups (Pdose < 0.05) to a similar degree (Pdose*group > 0.10). CO significantly increased in both groups (Pdose < 0.005) to a similar degree (Pdose*group > 0.10). There was no significant change in stroke volume in both groups. DO2 trended to a small increase (Pdose = 0.08) due to a small but significant decrease in hemoglobin in both groups (Pdose = 0.04). VO2 increased significantly in both groups (Pdose < 0.0001); however, the rate of VO2 increase was significantly less in SR group compared to Control group (Pdose*group < 0.0001). As a result, ERO2 increased significantly in both groups (Pdose < 0.0001). The rate of overall increase in ERO2 was significantly less in SR group (Pdose*group < 0.0001), and it started to decrease at the incremental dose of dopamine from 10 to 20 ug/kg/min. mERO2 did not increase significantly in both groups (P > 010). mERO2 was significantly lower in SR group (Pgroup = 0.01), with faster increase in SR groups compared to Control group (P < 0.0001). mERlac decreased significantly in both groups (P = 0.04). mERlac levels trended to be higher (P = 0.08) with faster decrease over the doses of dopamine (P = 0.04).
Discussion
This study demonstrates that in newborn lambs, SR59230A, a selective β3-adrenoreceptor antagonist, significantly attenuated the imbalance in systemic oxygen transport seen with incremental dopamine infusion. These increases in VO2 levels were significantly attenuated by SR59230A, whilst the increases in DO2 were not significantly different between the two groups. As a result, the balance of systemic and myocardial oxygen transport was improved, as indicated by the attenuated increase in ERO2 lower mERO2 and higher mERlac.
The goal of dopamine and other catecholamine therapy is to improve the balance of systemic oxygen transport by a greater increase in DO2 relative to changes in VO2. Both circulatory and metabolic stimulating effects by catecholamines share the same adrenergic signaling mechanisms, mainly, β1 and β2 and α-adrenergic receptors. If the circulatory responses to adrenergic stimulation are reduced, or the metabolic response enhanced, the beneficial effects of catecholamines might be abrogated. Such is the case in neonatal subjects particularly those with myocardial injury and increased VO2 as seen after cardiopulmonary bypass surgery. 6 It has been shown that α, β1, β2-adrenergic antagonists fail to improve the balance of oxygen transport, due to ineffective reduction of VO2 when each selectively used, or attenuation of both VO2 and DO2 to a similar degree when combined. 3
β3-adrenoreceptors also have both circulatory and metabolic effects, but distinctly different from other adrenergic receptors. In terms of metabolic effect, β3-adrenergic receptor are richly present in brown adipose tissue. 9 Brown adipose tissue generates heat following stimulation of α-, β1-, particularly β3-adrenoreceptors by the sympathetic nervous system. 7 It plays an essential role in non-shivering thermogenesis, hence VO2, in newborn humans and some in larger mammals including lambs. The ability to specifically activate or reduce energy expenditure via β3-adrenoreceptor manipulation is of much interest for the thermoregulation in newborns. Using the β3-adrenergic agonist Zeneca D7114 increased body temperature in moderately hypothermic Caesarean section-delivered lambs. 13 Selective and potent β3-adrenoreceptor antagonist SR59230A has been shown to down-regulate uncoupling protein-1 and norepinephrine induced cAMP accumulation, thus reducing thermogenesis in brown adipose tissue in animal experiments including sheep models.13–15 Our findings on the systemic effects of the attenuated rises in central body temperature and VO2 in newborn lambs treated with SR59230A prior to dopamine infusion are consistent with previous findings at tissue and molecular levels.13–15 The improved balance of systemic oxygen transport may, at least partly, contribute the improved myocardial oxygen transport as indicated by the lower mERO2 and higher mERlac in lambs treated with SR59230A be attributed, at least partly, to the improved balance of systemic oxygen transport.
Nonetheless, the specific cardiac effect of SR59230A should also be accounted for. Compared to brown adipose tissue, β3-adrenoreceptors in the heart have been less extensively studied, but have gained increasing interest in recent years. It has been reported that β3-adrenoreceptor stimulation exerts a profound dose-dependent negative inotropic effect in ventricles with decreased myocardial contractility in humans and other species.18–20 Hence, it is suggested that β3-adrenoreceptors serves as a “brake” during sympathetic overstimulation to antagonize β1-adrenoreceptors and β2-adrenoreceptors in the heart.20,21 In addition, β3-adrenoreceptor stimulation also exerts positive chronotropic effects, probably resulted from reflex mechanisms rather than from a direct stimulation of cardiac b3-adrenoceptors.22,23 It might be extrapolated that β3-adrenoreceptor antagonist may increase or maintain the inotropic effect of dopamine, thus myocardial contractility, but at the same time reduce the heart rate. As shown in our data, following SR59230A infusion, there was an immediate reduction in heart rate, but stroke volume was maintained. As a result, CO and DO2 decreased. Nonetheless, during incremental doses of dopamine infusion, the changes in heart rate, CO and stroke volume were not significantly different between the two groups. The lower heart rate and CO, thus reduced myocardial workload, might be attributable to the better balance of myocardial oxygen transport. It is important to note that, despite an initial reduction in heart rate and CO, the greater and continuous reduction in VO2 by SR59230A attenuated the imbalance of systemic oxygen transport at higher doses of dopamine infusion as indicated by the attenuated increase in ERO2.
Limitations
There are several limitations in our study. First, stroke volume was used to reflect cardiac contractility. The changes in myocardial contractility in the experiment may be more accurately assessed with direct measurement utilizing techniques such as tissue Doppler by echocardiography 24 or conductance catheter. 25 Secondly, myocardial lactate extraction ratio reflects the overall balance of the complex production and consumption of lactate by myocardium. A high lactate extraction may indicate less lactate production or more lactate consumption, or both. The exact mechanism can only be shown by direct examination of myocardial metabolism. Nonetheless, based on the lower myocardial oxygen extraction ratio in lambs treated with SR59230A, the less lactate production may be likely the case in our study. Thirdly, we did not directly assess the regional changes in oxygen consumption in brown fat tissue. Therefore, the relationship between VO2 and brown fat tissue remains speculative in our study. Forth, this is a pilot study in healthy newborn lambs using only one dose of SR59230A. A dose response study in newborn lambs with myocardial injury and increased VO2, such as newborn subjects following cardiopulmonary bypass may provide more detailed and relevant information about the effects on SR59230A on VO2 and myocardial contractility, thus CO and DO2.
Conclusion
Although β3-adrenoceptor antagonist SR59230A tends to induce an initial drop in heart rate and CO, it significantly attenuates the increase in VO2 and hence the imbalance of systemic and myocardial oxygen transport induced by dopamine at higher doses. Further studies are warranted to examine the dose response effect of SR59230A in subjects with myocardial injury and increased VO2 such as those after cardiopulmonary bypass surgery.
Authors' Contributions
RG, conducted the experiment and draft manuscript. PYC, designed the protocol, conducted the experiment and corrected manuscript. XY, MAA, JN, LGQ, YQL, JM, conducted the experiment, and corrected the manuscript. DBR and IMR, designed the protocol and advised on the animal surgery, and corrected the manuscript. JL, proposed the study, designed the protocol, conducted the experiment, analyzed the data, and corrected manuscript.
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.
Footnotes
List of Abbreviations
Acknowledgments
We thank Sally Cai, the Statistician in Data Centre, Congenital Heart Surgeons' Society, for her statistical advice.
The authors declare that we have no competing interests.