
Abstract
Although dental pain is a serious health issue with high incidence among the human population, its cellular and molecular mechanisms are still unclear. Transient receptor potential (TRP) channels are assumed to be involved in the generation of dental pain. However, most of the studies were conducted with molecular biological or histological methods. In vivo functional studies on the role of TRP channels in the mechanisms of dental pain are lacking. This study uses in vivo cellular electrophysiological and neuropharmacological method to directly disclose the effect of LaCl3, a broad spectrum TRP channel blocker, on the response properties of neurons in the mouse primary somatosensory cortex to low-temperature noxious stimulation of the dental pulp. It was found that LaCl3 suppresses the high-firing-rate responses of all nociceptive neurons to noxious low-temperature stimulation and also inhibits the spontaneous activities in some nonnociceptive neurons. The effect of LaCl3 is reversible. Furthermore, this effect is persistent and stable unless LaCl3 is washed out. Washout of LaCl3 quickly revitalized the responsiveness of neurons to low-temperature noxious stimulation. This study adds direct evidence for the hypothesis that TRP channels are involved in the generation of dental pain and sensation. Blockade of TRP channels may provide a novel therapeutic treatment for dental pain.
Keywords
Introduction
Dental pain, i.e., tooth pain, is a common symptom that is often caused by infection, tooth trauma, periodontal diseases, dentine hypersensitivity, and so on. Although dental pain is a serious health issue in daily life and its occurrence among the human population is very high,1,2 its cellular and molecular mechanisms are still unclear3–7 and the therapy for it is still limited nowadays. 1 Partly due to the unique structure of the tooth, dental pain is thought to be initiated via distinct mechanisms. 3 There are mainly three hypotheses proposed to elucidate the mechanism of tooth pain. The first hypothesis is called neural theory, which emphasizes the direct transduction of temperatures by nerve innervating the dentin and pulp.3,8 The second hypothesis postulates the sensory role of odontoblasts in the detection of thermal or mechanical stimuli.3,9 The third hypothesis, known as hydrodynamic theory, ascribes dental pain to the nociceptive responses resulted from the activation of nerve endings in the dentinal tubules by the movement of tissue fluid in it.3–5,10,11
Among these hypotheses, Brannstrom's classic hydrodynamic theory had been most widely accepted. However, some recent findings have challenged this theory. Of the most highlighted, transient receptor potential (TRP) channels, a group of nonselective cation channels, have been proposed to be involved in the mechanisms of dental pain.3,4,6,7 More recently, an “odontoblast receptor theory” was further proposed to explain the underlying mechanism of sensory transduction in dental sensation to noxious stimuli.6,12 In this theory, odontoblasts are deemed as sensory receptor cells through the activation of TRP channels.6,12 Shibukawa et al 12 investigated intercellular signal transduction between odontoblasts and trigeminal ganglion (TG) neurons, following the direct mechanical stimulation of odontoblasts. They found that mechanical stimulation of single odontoblasts increased the intracellular free calcium concentration by activating the mechanosensitive TRP channels. 12
TRP channel superfamily includes six subfamilies, namely canonical, vanilloid, melastatin, ankyrin, mucolipin, and polycystic TRP, 13 and contains totally 28 mammalian members, all of which are Ca2+-permeable, nonselective cation channels.14,15 TRP channels are polymodal channels that can be activated by voltage, temperature, mechanical, or chemical stimuli.16,17 Of these channels, a functional subset, including TRPV1, TRPV2, TRPV3, TRPV4, TRPA1, TRPM8, and TRPC5, is characterized by their temperature sensitivity,18,19 thus collectively referred to as thermo-TRPs, which cover a broad range of temperatures. For example, TRPV1, TRPM8, and TRPA1 are activated by >43°C, <25°C, and <17°C,3,15,18,20–22 respectively, and thus act as ideal primary transducers of noxious thermal stimuli. The recent progress of study on the roles of TRP channels in detecting noxious stimuli mainly includes two aspects, in vitro functionality and immunohistology or molecular biology. Using whole-cell patch clamp technique, Voets et al 18 showed that cooling up to 15°C or application of the cooling agent menthol activates a robust current in cells transiently transfected with TRPM8. Using single-channel recording technique, Moparthi et al 23 showed that cooling from 22°C to 10°C dramatically increases single-channel open probability of human TRPA1 channels, and the cold-induced activity is reversible and can be greatly inhibited by TRPA1 blocker. On the other hand, recent studies have also disclosed the expression of thermo-TRP channels, such as TRPV1, TRPV2, TRPM8, and TRPA1 in dental primary afferent neurons4,6,7,24–29 and TRPV1, TRPV2, TRPV3, TRPV4, and TRPM3 in odontoblasts,4,7,24,27,28 thus suggesting that TRP channels are responsible for dental pain elicited by noxious thermal stimuli.
However, a bottleneck in the research of the cellular mechanisms of dental pain is the lack of in vivo functional studies, possibly due to the difficulties of an in vivo electrophysiological technique, which is although a more direct approach to test in research animals whether a type of noxious stimuli is sensed or not and how intense the stimuli are sensed. In combination with neuropharmacological method, it will elucidate the roles that certain channels may play in the mechanisms of dental pain, and it can further provide evidence about whether a therapeutic intervention based on these particular channels is worth developing. In this study, I used in vivo cellular electrophysiological and neuropharmacological method to disclose the effect of LaCl3, a broad spectrum TRP channel blocker, on the response properties of neurons in the mouse primary somatosensory cortex to low-temperature noxious stimulation of the dental pulp.
Methods and Materials
Ethical Statement
The study was approved by the Animal Ethics Committee of Tianjin Medical University General Hospital. All the experiments performed in this study were consistent with International Association for the Study of Pain Guidelines for the Use of Animals in Research.
Animal Preparation
C57BL/6J male mice (body weight: 25–27 g) aged 8–10 weeks were used in this study. The animals were housed at room temperature with reversed 12-hour/12-hour light–dark cycles and provided with food and water ad libitum. The animals were allowed to habituate to the housing facilities for at least one week before the experiments.
The animals were initially anesthetized with urethane (1.2–1.5 g/kg in sterile saline solution, i.p.) and then sedated with chlorprothixene (10 mg/kg in water, i.m.). 30 Subcutaneous atropine (0.3 mg/kg) and dexamethasone (2 mg/kg) were also administered before the surgery to avoid respiratory secretions and brain edema, respectively. 30 The animals’ body temperature was maintained at 37.2 ± 2°C via a rectal probe and a homeothermic blanket. Oxygen was continuously supplied through a tube under the nose. Adequacy and depth of the anesthesia was monitored by the observation of the chest wall movement and the response to pinching a toe. Additional urethane (0.2–0.3 g/kg) was supplemented as needed during the experiment. Before the animals’ heads were fixed onto a stereotaxic apparatus, self-curing denture base resins were carefully placed surrounding the animals’ right lower incisors (Fig. 1A and B). A plastic drainage groove was embedded in the resins (Fig. 1B). The resins and the groove are used to prevent the cold-temperature solution from influencing other tissues during electrophysiology recordings. After the resins became hard enough, the animals’ right lower incisors were cut with a bone cutting forceps at about half the height of the teeth to expose the dental pulp (Fig. 1B). A successful acute tooth injury should be seen with red bleeding point at the top of the tooth, thus ensuring consistent pulp exposure and cavity depth. Thereafter, a saline-soaked cotton pellet covered the cavity until noxious thermal stimulation or chemicals were needed to be applied during electrophysiological recordings. The animals were then placed in a stereotaxic apparatus. The animals’ head were secured with ear bars and a nose clamp. A craniotomy was made above the left somatosensory cortex (~2 mm2) at ~2.8 mm lateral and ~1.5 mm anterior from the bregma point.31–33
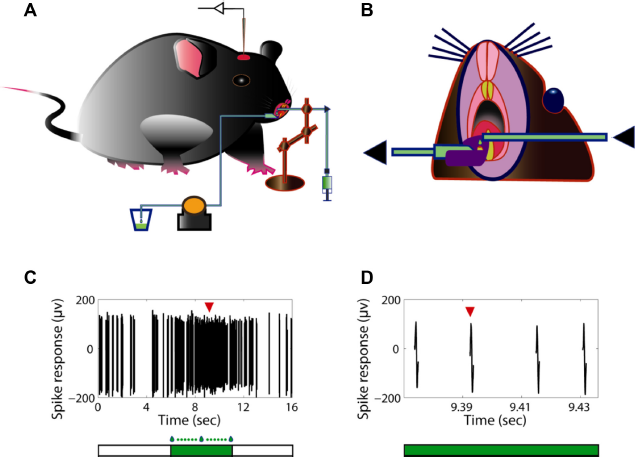
In vivo electrophysiological recording of mouse neurons in the S1 tooth area with noxious stimulation of the dental pulp.
In Vivo Electrophysiology
Extracellular recordings were performed with tungsten microelectrodes (5–7 Mō; FHC, Inc.). The electrode was vertically penetrated in increments of 1 µm using a pulse motor microdrive oil hydraulic manipulator (MO-81; Narishige). Recorded signals were preamplified through a differential headstage, amplified, and band-pass filtered (300 Hz–3 kHz) through a differential extracellular amplifier (EX4-400; Dagan). Single units were identified online using an oscilloscope and an audio monitor. Data were digitized at 25 kHz with a high-speed data acquisition interface (CED Power1401 mkII; Cambridge Electronic Design Ltd) and recorded on an Intel Core 2 Duo workstation computer using the Spike2 software (version 6; Cambridge Electronic Design Ltd) for further off-line analysis. The animals were killed at the end of recordings by an overdose of pentobarbital.
Noxious Thermal Stimulation and Application of LaCl3
In order to exert the noxious thermal stimulation (low temperature) or chemical substance over the exposed dental pulp, a piece of PE tubing (PE-160; Warner Instruments) was placed over the top of the injured tooth (Fig. 1A and B). One end of the tubing was securely held with a turnable metal arm, and the other end was connected with a three-way coupling that was further ready to be connected with a syringe prepared with cold physiological saline solution (0.9% NaCl) or cold LaCl3 (Sigma-Aldrich) solution (100 µM in saline solution). 34 The syringes prefilled with solution were stocked in an ice box in advance. During recordings, when noxious thermal stimulation or LaCl3 is needed to be administered, a syringe filled with the corresponding solution (with temperature lower than 10°C) was connected to the tubing coupling (Fig. 1A). A specified keyboard button will be pressed down to write an event marker in the recorded data file through the Spike2 software at the beginning and at the end of applying solution on the teeth. Generally, the duration of applying solution was controlled for about five seconds each time (Fig. 1C). The noxious thermal stimulation with or without LaCl3 was repeated for three to five times for each recorded cell. Washout of LaCl3 with 37°C physiological saline water was performed after the completion of LaCl3 effect observation. An additional test with cold physiological saline solution was then carried out to observe if the effect of LaCl3 is reversible.
Data Analysis
Off-line data analysis was performed using the Spike2 software (version 6; Cambridge Electronic Design) and Matlab (R2008a; MathWorks). Action potentials (Fig. 1D) were discriminated and sorted based on their waveforms in the Spike2 software. Data were analyzed and plotted in Matlab. For the calculation of firing rates, an observation time window was equally divided into many bins of 150 ms. The firing rate in each bin was calculated by dividing the spike count by the bin width. The average firing rate over the whole observation time window was then calculated. The length of the shortest time window for the observation of spontaneous activities was 5 minutes, while two continuous blocks of time window, i.e., 10 minutes, were executed before the administration of any stimulus. The length of each time window for the observation of evoked responses was five seconds, which is the same as the duration of applying stimulation for each time (Fig. 1C). All the statistical analyses were performed with Matlab statistics toolbox. Student's t-test was used to compare continuous variables, and the chi-square test was used for nonparametrical variables. Differences were considered to be significant if P < 0.05. All numerical values are expressed as mean ± SEM.
Results
Characteristics of Spontaneous Activities of Neurons in S1 Tooth Area
I gained single-unit recordings from 106 neurons in the primary somatosensory cortex of the contralateral lower incisor. The probability of obtaining recordings is higher at the deeper depths than that at the lower depths. A total of 70.8% (75/106) of neurons were recorded at the depths of 400-800 µm, and 29.2% (31/106) were recorded at the lower depths (<400 µm). The majority of cells (93/106, 87.7%) showed spontaneous firings in various degrees. The mean spontaneous firing rate was 7.4 ± 0.36 Hz (n = 93) in a 10-minute recording duration for each cell. The neurons in the deeper depths showed higher firing rates than those in the lower depths (8.8 ± 0.38 Hz vs 3.9 ± 0.27 Hz, t = 7.90, P < 0.001). Furthermore, the recorded neurons showed different firing patterns during spontaneous activity recordings: 81.7% (76/93) cells showed burst firing patterns, 7.5% (7/93) cells showed regular firing patterns, and 10.8% (10/93) cells showed irregular firing patterns. The neurons in the deeper depths showed higher probability of burst firing than those in the lower depths (77.3% vs 58.1%, x = 4.01, P < 0.05).
Response Properties of Neurons in S1 Tooth Area to Low-Temperature Noxious Stimulation
I used intermittently exerted low-temperature noxious stimulation to test the response properties of the recorded neurons. In the end, 74 cells (69.8%) were verified as neurons with properties responding to noxious stimulation (nociceptive neurons, Fig. 2A–C), while others showed no response to noxious stimulation (Fig. 2D and Supplementary Fig. 1A and B). A total of 71.6% (53/74) of the nociceptive neurons were recorded at the depths deeper than 400 µm, and 28.4% (21/74) were recorded at the lower depths. The average firing rates to noxious stimulation of neurons at different depths showed no significant differences (23.4 ± 1.63 Hz for the deeper cells vs 20.3 ± 2.23 Hz for the lower cells, t = 1.06, P = 0.293). In 74 nociceptive neurons, 13 cells showed firings only when noxious stimulation was given (Fig. 2C and Supplementary Fig. 2A and B). The average firing rate to noxious stimulation in these cells was 3.5 ± 0.18 Hz (Fig. 2C). The rest of the nociceptive neurons showed different patterns of firing (burst, regular, or irregular firing) during spontaneous activity recordings. The firing rates of all these neurons were significantly higher when noxious stimulation was given than during spontaneous activity recordings (26.5 ± 1.03 Hz vs 7.5 ± 0.45 Hz, t = 16.9, P < 0.001, Fig. 2C).
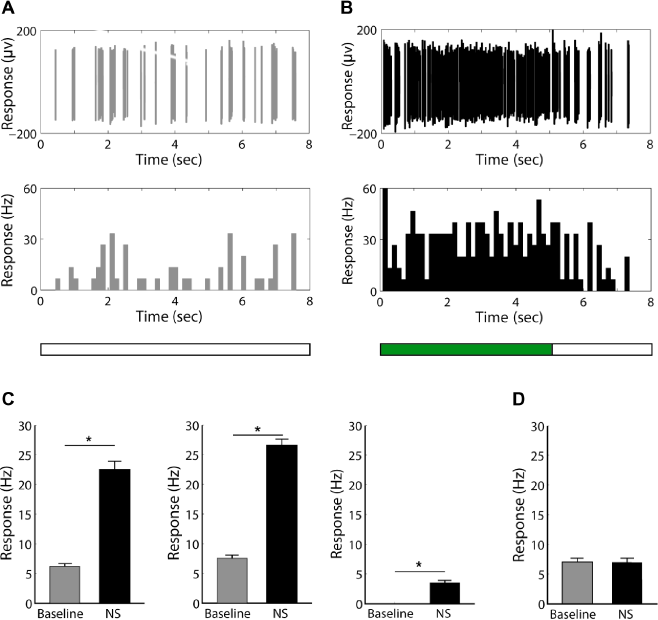
Responses of neurons in the S1 tooth area to low-temperature noxious stimulation.
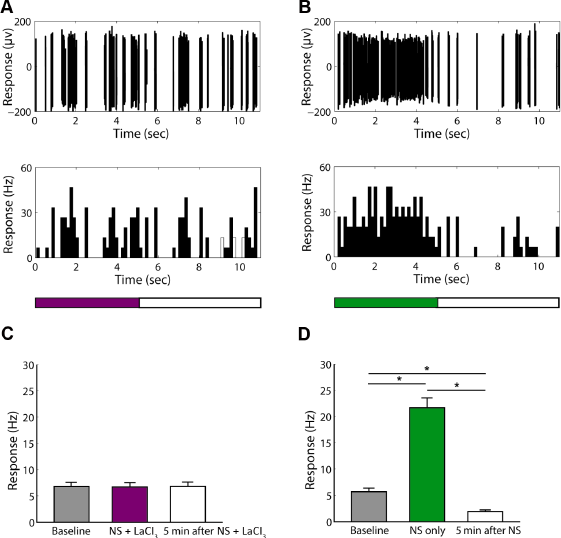
LaCl3 Reversibly suppressed the Hypersensitive Responses of Neurons in S1 Tooth Area to Low-Temperature Noxious Stimulation
Next, I investigated the effect of LaCl3, a broad spectrum TRP channel blocker, on the response characteristics of nociceptive neurons and nonnociceptive neurons in the S1 tooth area. I found that LaCl3 unexceptionally suppressed the high-firing-rate responses of all nociceptive neurons to noxious low-temperature stimulation (Fig. 3A–C and Supplementary Fig. 3A and B), while a few of the nonnociceptive neurons tested with LaCl3 also showed suppressed spontaneous activities (Fig. 4A–C). For the nociceptive neurons, the average firing rate during stimulation was significantly decreased from 23.4 ± 1.88 Hz to 6.7 ± 0.72 Hz (t = −8.27, P < 0.001, n = 36; Fig. 3C) and was not significantly different from the baseline activities (6.7 ± 0.72 Hz vs 6.8 ± 0.72 Hz, t = −0.09, P = 0.932, n = 36; Fig. 3C). For the nonnociceptive neurons, although most of the cells showed no change of responses to stimulation with LaCl3 (Supplementary Fig. 4A and B), 37.5% (6/16) cells showed decreased average firing rate (3.4 ± 0.57 Hz) during stimulation with LaCl3 compared with stimulation without LaCl3 (t = −2.58, P = 0.028) and the baseline activities (t = −2.63, P = 0.025), respectively (Fig. 4C).
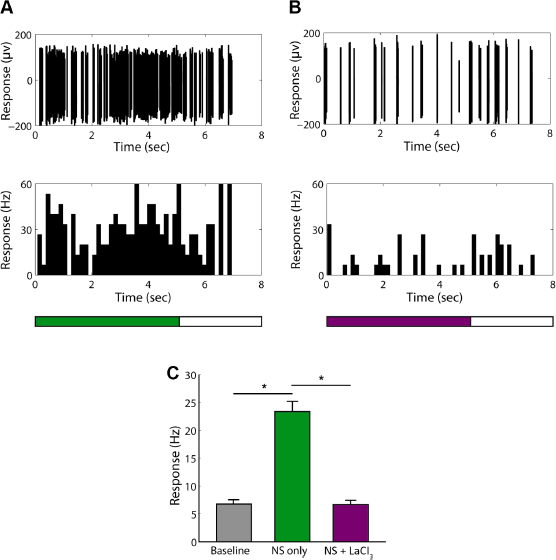
The effect of LaCl3 on the responses of nociceptive neurons to low-temperature noxious stimulation.
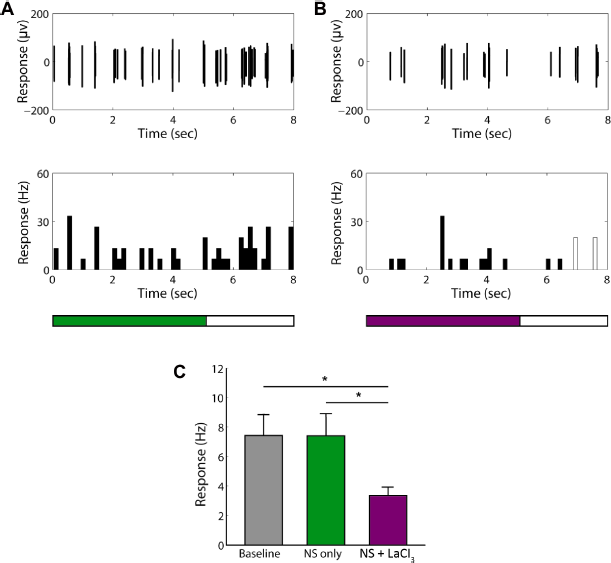
The effect of LaCl3 on the responses of some nonnociceptive neurons to low-temperature noxious stimulation.
Afterward, I observed the short-term and long-term aftereffects of LaCl3 on the spontaneous activities in the nociceptive neurons after stopping the application of cold saline solution with LaCl3. I defined the aftereffects as the change of spontaneous activities during a short (five minutes) or long (~60 minutes) period of time after stopping the application of cold solution compared with the baselines or the evoked responses, when the low-temperature stimulation has been ceased, but LaCl3 is still kept in the pulp tissue. For comparison, 74 cells were randomly assigned into groups with or without application of LaCl3 during low-temperature noxious stimulation (NS + LaCl3 group and NS group). I found that the spontaneous firing rates in the first five minutes after stopping stimulation in the NS + LaCl3 group did not show a significant change comparing with that of the corresponding baselines (6.5 ± 0.70 Hz vs 6.8 ± 0.72 Hz, t = −0.28, P = 0.813, n = 36; Fig. 5A and C), while in the NS group, the spontaneous firing rates decreased to lower than the corresponding baselines (1.9 ± 0.23 Hz vs 5.7 ± 0.70 Hz, t = −5.21, P < 0.001, n = 38; Fig. 5B and D). There was a significant difference of spontaneous firing rates in the first five minutes in these two groups (6.5 ± 0.70 Hz vs 1.9 ± 0.23 Hz, t = 6.42, P < 0.001). I also found that in the NS group, some nociceptive neurons showed gradually increased spontaneous activities from 10 to 60 minutes comparing with their baselines, even up to the level as that during the period the noxious cold-temperature stimulation was given (Fig. 6A and B), while others only showed temporarily increased spontaneous activities at ~10-20 minutes (Fig. 6A) and then recovered to the baseline levels in the subsequent observation time (Fig. 6A and B). However, in the NS + LaCl3 group, no cell showed significant increased spontaneous activities in a long period of time up to 60 minutes (Fig. 6A and B).
Short-term aftereffects of low-temperature noxious stimulation with or without LaCl3 in nociceptive neurons in the S1 tooth area.
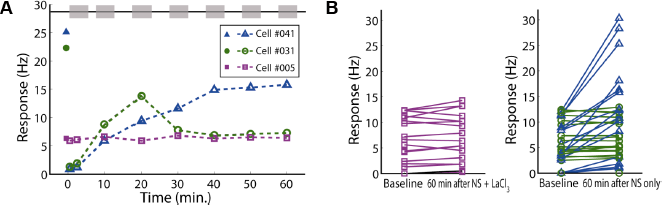
Long-term aftereffects of low-temperature noxious stimulation with or without LaCl3 in nociceptive neurons in the S1 tooth area.
Finally, I investigated whether the effect of LaCl3 on the responses of neurons in the S1 tooth area is reversible after washout (Fig. 7A). Nineteen cells in which washout experiments were performed immediately after short-term aftereffect observation (five minutes) showed quickly recovered high-firing-rate responses to the noxious cold-temperature stimulation (Fig. 7B), and 17 cells in which washout experiments were performed after long-term aftereffect observation (60 minutes) also showed quickly recovered high-firing-rate responses to the noxious cold-temperature stimulation (Fig. 7C). The firing rates responding to the noxious cold-temperature stimulation were not significantly different from the levels before the application of LaCl3 (24.0 ± 1.95 Hz vs 23.4 ± 1.88 Hz, t = −0.23, P = 0.817, n = 36).
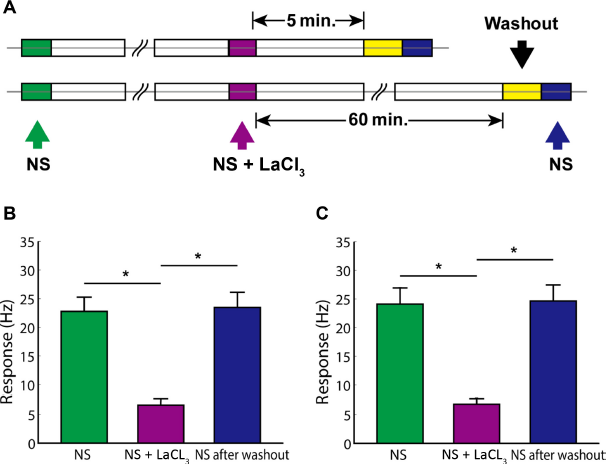
The effect of LaCl3 on the responses of neurons in the S1 tooth area is reversible after washout.
Discussion
Role of TRP Channels in Dental Pain
The classic hydrodynamic theory had been most widely accepted. However, in the past decade, accumulating evidence implies that TRP channels may be the most possible molecular mechanisms in the generation of dental pain, although it may take an effect through different hypothesized mechanisms. In the neural theory, to play a sensory role, the nociceptive TG neurons that innervate dental pulp must express receptors that take part in the transduction of a specific stimulus to electrical impulses. 3 It has been revealed that several temperature-sensitive TRP channels, such as TRPV1, TRPV2, TRPM8, and TRPA1, express in TG neurons and, more specifically, in dental primary afferent neurons. 4 In the “hydrodynamic theory,” it describes the cause of dental pain in terms of mechanical forces generated by the movement of dentinal fluid. 3 Lin et al suggested that the inward and outward movement of dentinal fluid contributes differentially to dental pain. 5 They demonstrated that intradental mechanosensitive nociceptors are not “equally sensitive” to inward (into the pulp) and outward (away from the pulp) fluid flows, thus providing mechanistic insights into the difference between hot and cold dental pain. 5 However, a key issue is that a molecular transducer that accounts for the mechanical detection of dentinal fluid movement has to be discovered.3,4 Some mechanosensitive TRP channels, such as TRPV1, TRPV2, TRPA1, TRPV4, and TRPM3, have shown expression in dental afferent neurons or TG neurons. 4 Finally, in the odontoblast transducer theory, it is suggested that odontoblasts play a role as sensory transducers and then transmit the signal to dental afferent neurons. A series of TRP channels have been confirmed their expression in odontoblasts. Furthermore, it has been shown that the cooperation of TRP channels with mechanosensitive K+ channels participates in the perception of temperature.35,36 The mechanism of signaling between odontoblasts and dental afferent neurons has recently also obtained a progress. It has been suggested that adenosine triphosphate (ATP) may mediate painful signaling between odontoblasts and neurons. 37 Thus, TRP channels should play a key role in the generation of tooth pain and sensation. However, most of the studies used molecular biological and morphological methods to investigate the expression of TRP channels and then deduce its role in the mechanisms of dental pain and sensation. No study has directly shown the link between TRP channels and the generation of dental pain and sensation in vivo.
In this study, a direct functional method, in vivo cellular electrophysiological and neuropharmacological method, was used to investigate if TRP channels may take a role in the mechanisms of dental pain and sensation. I examined the effect of LaCl3 on the response properties of neurons in the S1 tooth area to low-temperature noxious stimulation of the dental pulp. LaCl3 is a broad spectrum TRP channel blocker. It has been confirmed that trivalent cations, such as La3+ and Gd3+, can block nearly all TRP channels.38,39 Specifically, LaCl3 at low concentration (100 µM) has shown its inhibitory effect in blocking thermo-TRP channels.34,40,41 Furthermore, it has been established that long-term low dose of lanthanum has no neurotoxicity in rats. 42 In this study, I found that LaCl3 unexceptionally and almost completely suppressed the high-firing-rate responses of all nociceptive neurons to noxious low-temperature stimulation and partially inhibited the spontaneous activities in some nonnociceptive neurons. The results also suggest that the effect of LaCl3 on the neurons’ responses to low-temperature noxious stimulation is reversible, but this effect is persistent and stable unless LaCl3 is washed out. Washout of LaCl3 at any time can quickly revitalize the responsiveness of neurons to low-temperature noxious stimulation. At the same time, this study also showed that there was inhibition of spontaneous firing immediately after the noxious stimulation, which suggests a homeostatic mechanism in the primary somatosensory cortex to balance the superexcitation by noxious stimulation. Interestingly, some nociceptive neurons showed gradually and persistently increased spontaneous firings with time. This implies that noxious stimulation may hypersensitize the pain feeling to the stimuli. Another effect of LaCl3 is that it may alleviate the hypersensitization effect of the noxious stimulation. Thus, the results of this study provide additional evidence for the hypothesis that TRP channels are involved in the generation of dental pain and sensation.
Clinical Significance
Dental pain is a common dental health problem, which not only seriously affects the quality of life of millions of people worldwide but also leads to increased economic burden on families and society costs. In this study, a direct functional method, in vivo cellular electrophysiological and neuropharmacological method, was used to investigate if TRP channels may take a role in the mechanisms of dental pain and sensation. The results of this study not only verify that TRP channels definitely participate in the generation of dental pain and sensation but also provide the basic information that will be helpful for further translational medical research. According to the animal research in this study, TRP channel blockers can quickly act on the exposed dental pulp to decrease the responses of nociceptive cells in the somatosensory cortex. This effect is persistent and stable and can be quickly reversed after washout. Thus, the data of this study directly showed that TRP channel blockers are promising to be used in the treatment of dental pain and provide the baseline materials for the research and development of new drugs targeting specific TRP channels in the future.
Conclusions
In summary, this study has examined the effect of LaCl3, a broad spectrum blocker of TRP channels, on the response properties of neurons in the S1 tooth area to low-temperature noxious stimulation of the dental pulp and showed its inhibitory effect on the activities of neurons in the S1 tooth area. For the first time, this study provides the in vivo physiological data on the functions of TRP channels in the mechanisms of dental pain and sensation. This will guide future investigations to fill the gap between histology and physiology on the cellular and molecular mechanisms of dental pain and promote the research on the development of new medicines targeting TRP channels.
Author Contributions
Conceived and designed the experiments: YJ. Analyzed the data: YJ. Wrote the first draft of the manuscript: YJ. Made critical revisions: YJ. The author reviewed and approved of the final manuscript.
Footnotes
Acknowledgments
The author would like to thank Xuefeng Shi (Tianjin Eye Hospital, Tianjin, China) for his advice on the experimental design and his help in analyzing the data. The author is grateful to Xiaojun Zhang (University of South California, Los Angeles, CA, USA) for the constructive comments and editing and also would like to thank the reviewers for their helpful suggestions in the previous version of this manuscript.