
Abstract
Background:
Junín virus (JUNV) and several other clade B New World arenaviruses cause human disease ranging from mild febrile illness to severe viral haemorrhagic fever. These viruses pose a significant threat to national security and safe and effective therapies are limited except in Argentina, where immune plasma is the standard of care for treating JUNV infection in cases of Argentine haemorrhagic fever.
Methods:
An in vitro screen of the Chemtura library identified several compounds with activity against Tacaribe virus (TCRV), a clade B arenavirus closely related to JUNV. Of these compounds, D746, a phenolic dibenzylsulfide, was further pursued for additional in vitro studies and evaluated in the AG129 mouse TCRV infection model.
Results:
D746 was found to act during an early to intermediate stage of the TCRV replication cycle and μM range activity was confirmed by virus yield reduction assays with both TCRV and JUNV. Although intraperitoneal twice daily treatment regimens were found to be highly effective when started 2 h prior to TCRV challenge in AG129 mice, post-exposure treatment initiated 3 days after infection was not efficacious. Interestingly, despite the pre-exposure treatment success, D746 did not reduce serum or tissue virus titres during the acute infection. Moreover, D746 elicited ascites fluid accumulation in mice during, as well as independent of, infection.
Conclusions:
Our findings suggest that D746 may be altering the host response to TCRV infection in AG129 mice in a way that limits pathogenesis and thereby protects mice from otherwise lethal infection in the absence of measurable reductions in viral burden.
Introduction
Several clade B New World arenaviruses cause disease ranging from mild febrile illness to fatal viral haemorrhagic fever (HF) in humans. These arenaviruses are transmitted to humans typically through contact with rodent reservoirs or inhalation of aerosolized virus passed in their excreta. Junín virus (JUNV) is the aetiologic agent of Argentine HF, a disease that can evolve into severe haemorrhagic and neurological manifestations in some patients with a 15–30% fatality rate in cases requiring medical attention [1]. The National Institute of Allergy and Infectious Diseases has classified JUNV and other pathogenic clade B viruses as category A priority pathogens because they pose a significant risk to national security and public health [2].
The non-specific antiviral, ribavirin, is the only licensed drug available to treat arenaviral HF under emergency provisions [3]. However, there is very limited data to support the use of ribavirin to treat severe infections with New World HF arenaviruses [1,4]. In addition, ribavirin is also associated with toxicity in the form of a reversible haemolytic anaemia in humans [5,6], and teratogenic effects observed in rodents [7]. The threat of intentional release of highly pathogenic arenaviruses by terrorist groups combined with the risk of naturally occurring outbreaks in certain regions of the world underlines the need for the development of safe and more effective therapies.
Tacaribe virus (TCRV) is a clade B arenavirus that is closely related to JUNV and other pathogenic clade B arenaviruses [8], but is not considered a frank human pathogen [9,10]. Consequently, early-stage drug discovery research is greatly facilitated as TCRV can be handled in biosafety level 2 laboratories rather than in stringent biosafety level 4 maximum containment required for JUNV and other HF arenaviruses. To this end, we and others have previously reported on the use of cell culture-based cytopathic TCRV infection assays to identify and evaluate promising anti-arenavirus compounds [11,12].
With the objective of discovering and developing novel anti-arenavirus compounds, we used TCRV infection assays as a screen for the Chemtura library, wherein we identified several compounds with antiviral activities against New World arenaviruses. Here we report on one compound, a phenolic dibenzylsulfide referred to as D746, which consistently demonstrated activity against TCRV and other New World arenaviruses in more rigorous virus yield reduction assays. Studies employing the TCRV AG129 mouse infection model, based on challenge of type I and II interferon receptor knockout mice [13], revealed D746 activity as a pre-exposure prophylaxis, but not as a post-exposure intervention. Remarkably, D746 pretreatment did not reduce viral titres, but did elicit accumulation of ascites fluid, suggesting that an immune-mediated alteration of the host response may confer the observed protective effect.
Materials and methods
Animals
AG129 interferon receptor-deficient mice were bred at Utah State University (Logan, UT, USA) under specific pathogen-free conditions. They were fed irradiated mouse chow and autoclaved water ad libitum. For the experiments, 5–10-week-old mice were used. All animal procedures used in this study complied with guidelines set by the US Department of Agriculture and Utah State University Animal Care and Use Committee.
Viruses
TCRV, strain TRVL 11573, was obtained from American Type Culture Collection (ATCC; Manassas, VA, USA). The virus stock used for in vitro experiments was derived from clarified cell lysates produced by three passages in Vero 76 African green monkey kidney cells. The virus stock used for mouse studies was from clarified liver homogenates from AG129 mice challenged with the in vitro stock. The JUNV Candid #1 strain was obtained from Robert Tesh (World Reference Center for Emerging Viruses and Arboviruses, University of Texas Medical Branch, Galveston, TX, USA). The Candid #1 virus used was passaged once in B-SC-1 epithelial African green monkey kidney cells and once in Vero cells. Pichindé virus (PICV), strain An 4763, was provided by David Gangemi (Clemson University, Clemson, SC, USA). The PICV used was passaged once through hamsters and once in Vero cells.
Test compounds
D746 was provided by Chemtura Corporation (Guelph, ON, Canada). For in vitro activity experiments, D746 was dissolved in DMSO, then further diluted in sterile minimal essential medium (MEM; Thermo Scientific Hyclone, Logan, UT, USA) to final DMSO concentrations of <0.5%. For efficacy studies in mice, D746 was dissolved in ethanol, and then further diluted in Cremaphor® (Sigma–Aldrich, St Louis, MO, USA) and saline (CES) at a ratio of 1:1:8. Ribavirin was supplied by ICN Pharmaceuticals, Inc. (Costa Mesa, CA, USA) and was prepared in sterile saline. All treatments were administered by intraperitoneal injection of 0.1 ml.
Cell culture antiviral assays
Vero cells were obtained from ATCC and maintained in MEM supplemented with 0.18% NaHCO3 and 10% fetal bovine serum (Thermo Scientific Hyclone). Viruses were titrated in MEM culture medium containing 2% fetal bovine serum to identify the optimal infectious dose that produced maximal cytopathic effect (CPE) in <9 days. Varying concentrations of D746 and ribavirin (positive control) were added to test wells containing 70–80% confluent Vero cells at the time of infection with viruses at a multiplicity of infection of approximately 0.001. Plates were incubated at 37°C, 5% CO2, until virus-infected control wells were observed to have maximal viral CPE (7–8 days), at which time the plates were processed to determine virus yield reduction (VYR) as previously described [11]. Concentrations of compound that reduced virus yield by 1 log10 (90% effective concentration [EC90]) were determined by regression analysis. For toxicity determinations done in parallel, compounds were added to uninfected Vero cells and the drug concentrations that reduced cell viability by 50% (50% cytotoxic concentration [CC50]) were also determined by regression analysis. Selectivity index values were calculated as CC50/EC90 for the VYR assays.
For time-of-addition assays, subconfluent Vero cell monolayers in 24-well plates were inoculated with TCRV at a multiplicity of infection of 0.02 and incubated at 37°C, 5% CO2 for 1 h. Triplicate wells were incubated with varying concentrations of D746 or MEM culture medium at −2–0, 0–3, 3–6 and 6–9 h relative to the time of infection. Following the 2–3 h treatment periods, the media was replaced with new growth media devoid of D746. Plates were frozen at −80°C 24 h post-infection and viral CPE was determined for calculation of 50% cell culture infectious dose (CCID50) as previously described [14].
In vivo D746 efficacy studies
In all experiments, AG129 mice were sorted into treatment groups of 10–22 mice to minimize gender and age differences and challenged intraperitoneally with 0.2 ml of TCRV prepared in MEM. D746, ribavirin and CES vehicle placebo treatments were administered intraperitoneally twice daily to achieve the stated mg/kg/day dose. The duration of treatment for all experiments was 7 days. For the prophylactic efficacy experiments, treatment was started 2 h prior to challenge with 150 CCID50 of TCRV. For the evaluation of post-exposure efficacy, D746 treatment was initiated 3 days after challenge with 450 CCID50 of TCRV. The increase to 450 CCID50 was made in order to achieve uniform lethality, which was not observed at the lower dose. In two of the experiments, groups of 4–5 animals were sacrificed on day 9 of infection for analysis of serum and tissue virus titres. The remaining animals were observed for 3–4 weeks for morbidity and mortality. Body weights were assessed every 3 days starting on day 0, relative to time of challenge.
Serum and tissue virus titres
Virus titres were assayed using an infectious end point dilution cell culture assay as previously described [11]. Briefly, a specific volume of tissue homogenate or serum was serially diluted and added to triplicate or quadruplicate wells of Vero 76 cell monolayers in 96-well microplates. The viral CPE was determined after a 10-day incubation, and the 50% end points were calculated as described [14]. The assay detection limits are designated by the lowest point on the y-axis of the virus titre graphs.
Statistical analyses
Two-way analysis of variance (ANOVA) with Bonferroni post-test was used to analyse the time-of-addition data. The Mantel–Cox log-rank test was used for analysis of Kaplan–Meier survival curves. One-way ANOVA with the Newman–Keuls post-test was performed to analyse virus titre differences. All statistical evaluations were done using Prism® (GraphPad Software, La Jolla, CA, USA).
Results
In vitro antiviral activity of D746
Following the initial ‘hit’ of the phenolic dibenzylsulfide, D746 (Figure 1), against TCRV infection during the screening of the Chemtura library, the inhibitory effect of the compound was confirmed using more rigorous secondary testing assays. As shown in Table 1, D746 was active in the μM range against TCRV, JUNV and PICV by VYR assays, with selectivity index values ranging from 5 to 21. Notably, the D746 TCRV inhibition dose–response curve was shallow compared to ribavirin (Figure 2A). The curve representing cell cytotoxicity of the compound relative to ribavirin is shown in Figure 2B. D746 was inactive against the MP-12 vaccine strain of Rift Valley fever virus, dengue virus (serotype 2), West Nile virus, yellow fever virus, Venezuelan equine encephalitis virus, SARS coronavirus, and several strains of influenza virus in primary testing (data not shown). These results demonstrate the specificity of D746 towards arenaviruses.
Summary of in vitro anti-arenavirus activity of D746 and ribavirin
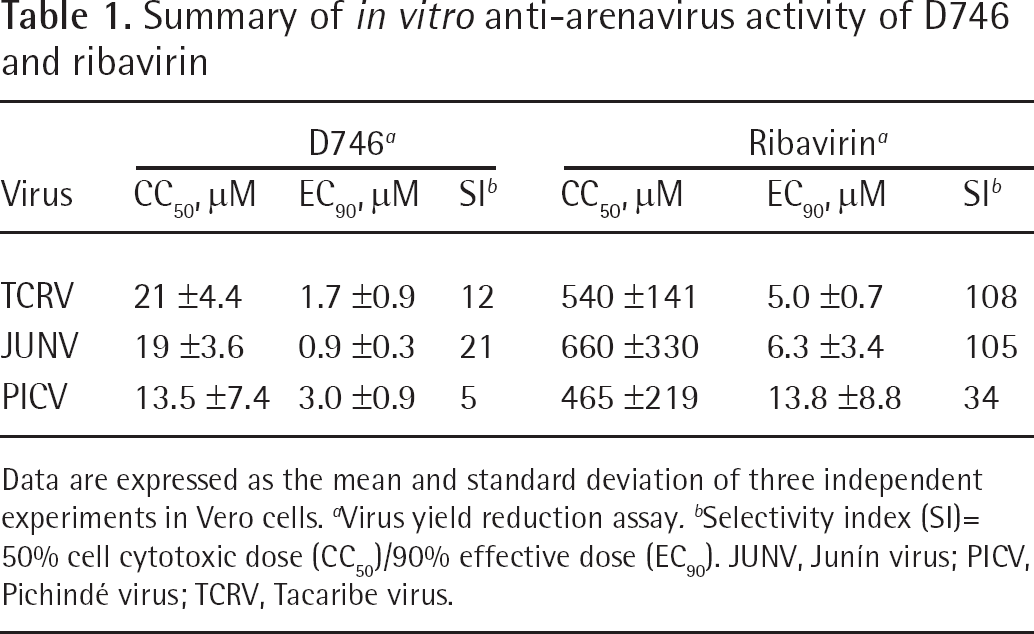
Data are expressed as the mean and standard deviation of three independent experiments in Vero cells.
Virus yield reduction assay.
Selectivity index (SI)= 50% cell cytotoxic dose (CC50)/90% effective dose (EC90). JUNV, Junín virus; PICV, Pichindé virus; TCRV, Tacaribe virus.
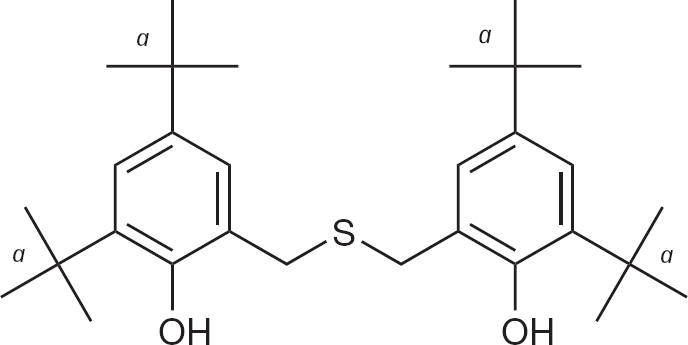
Chemical structure of D746, a phenolic dibenzylsulfide
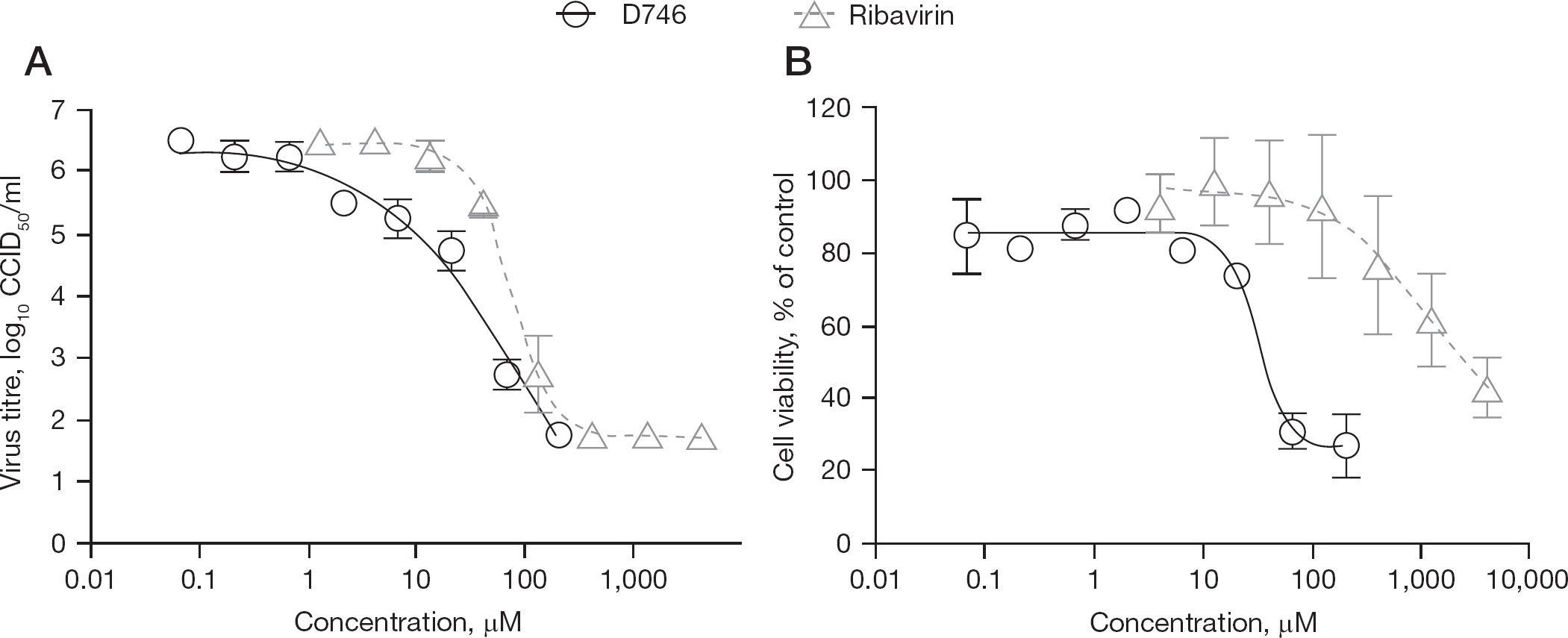
Effects of D746 and ribavirin on virus yield and cell viability
D746 time-of-addition effect on TCRV infection
Time-of-addition studies were conducted to provide insights into the mechanism by which D746 inhibits arenavirus infection. Vero cell monolayers were treated by the addition of D746 at various times prior to and during TCRV infection and the reduction in virus production relative to the untreated cultures was assessed at 24 h post-infection. The greatest inhibition of TCRV replication was observed when the cells were pretreated, with diminishing dose–responsive activity seen out to the 3–6 h treatment time (Figure 3). There was no significant inhibition seen when cells were treated 6 h after infection. We have previously reported that ≥14 h are required for the detection of infectious TCRV particles in Vero culture supernatants [15], suggesting that virus detected after 24 h in culture is from the initial infection cycle. Thus, our findings suggest that the effects of D746 target an early to intermediate stage of the virus life cycle, with inhibition of virus adsorption being likely.
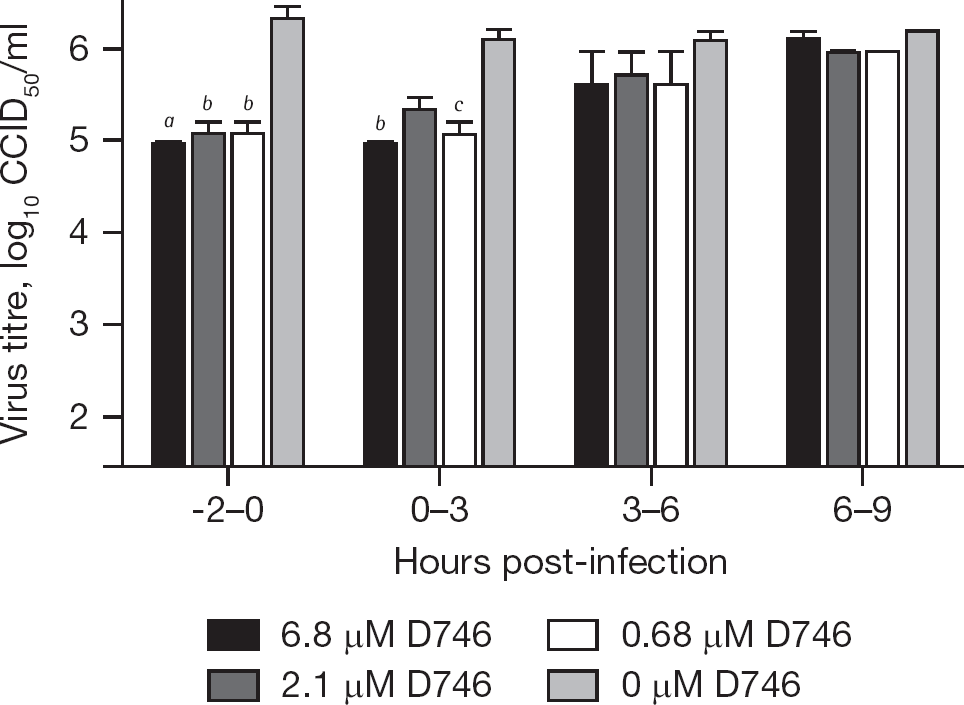
Inhibitory effects of D746 added during specified times relative to Tacaribe virus infection
Prophylactic efficacy of D746 against TCRV infection in mice
Considering the anti-arenavirus activity of D746 in cell culture, we next evaluated the compound in the TCRV AG129 mouse infection model. For the first efficacy study, we selected a 7-day treatment regimen consisting of daily doses of 100 and 10 mg/kg, administered by intraperitoneal injections every 12 h starting 2 h prior to TCRV challenge. These doses were well tolerated by uninfected AG129 mice in an initial mean tolerated dose study (data not shown). As seen in Figure 4, the 100 mg/kg/day dose was highly effective, protecting 6 of 8 (75%) infected AG129 mice from mortality compared to 4 of 22 (18%) survivors in the placebo group that received CES vehicle. There were no significant differences in survival curve analysis or mean day of death comparison between the groups treated with 10 mg/kg/day D746 versus placebo. Unexpectedly, ribavirin, routinely included as a positive control, failed to protect TCRV-challenged mice at a daily total dose (50 mg/kg/day) previously reported to be effective [13]. In the aforementioned studies, ribavirin was given as a once-daily (50 mg/kg) dose, whereas in the present study, it was divided into two 25 mg/kg doses every 12 h. It is conceivable that the larger bolus dose reached more effective concentrations in critical areas, thereby resulting in a measurable protective effect. Furthermore, the added stress due to twice-daily injections may have also contributed to the ineffectiveness observed in this initial experiment with D746.
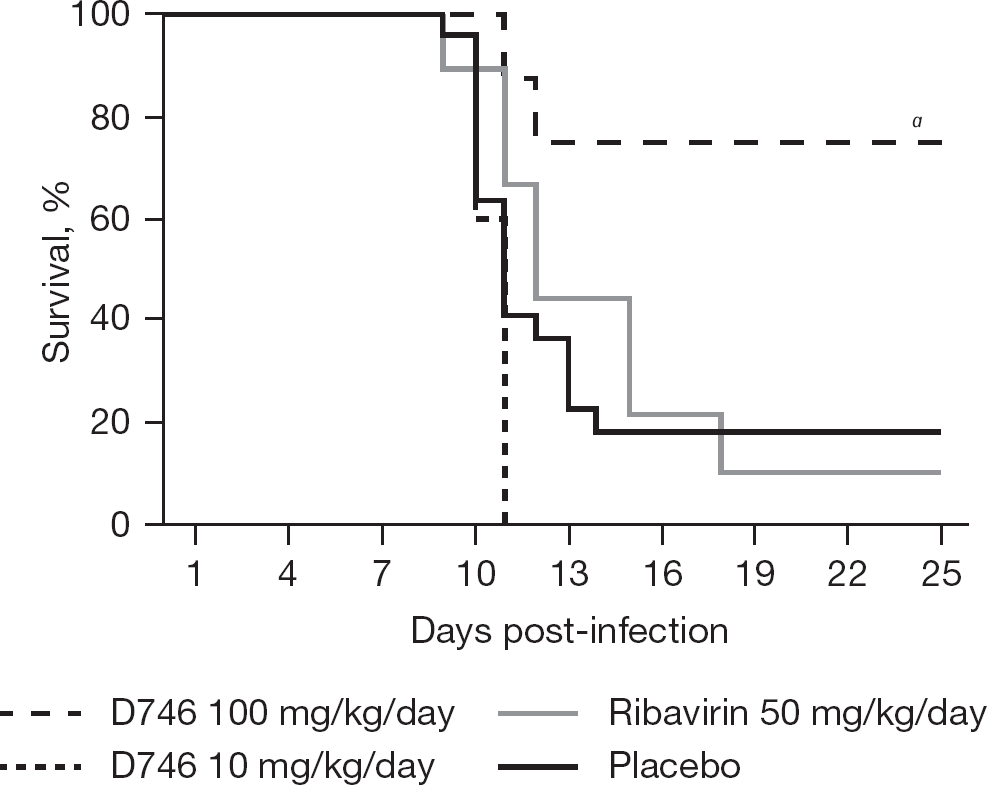
Prophylactic efficacy of D746 in the AG129 mouse Tacaribe virus infection model
Because significant protection was afforded by prophylactic D746 treatment in the first experiment, a second study that included higher doses of compound and additional animals for determining reductions in virus titre was conducted. All 10 mice treated with the highest dose of 200 mg/kg/day D746 survived the TCRV challenge, compared to 25% (3 of 12) in the placebo group (Figure 5A). In addition, significant protection (70% survival) was also observed at D746 doses of 100 and 50 mg/kg/day, confirming the results from the first experiment with the 100 mg/kg/day dose. Half of the mice that received the higher dose of ribavirin given for the second experiment survived the infection; however, due to the 25% survival rate in the placebo group, the difference was not significant. Doses of up to 200 mg/kg/day were also evaluated in parallel in uninfected animals with some weight loss observed, but all mice ultimately recovered following the cessation of treatment (data not shown).
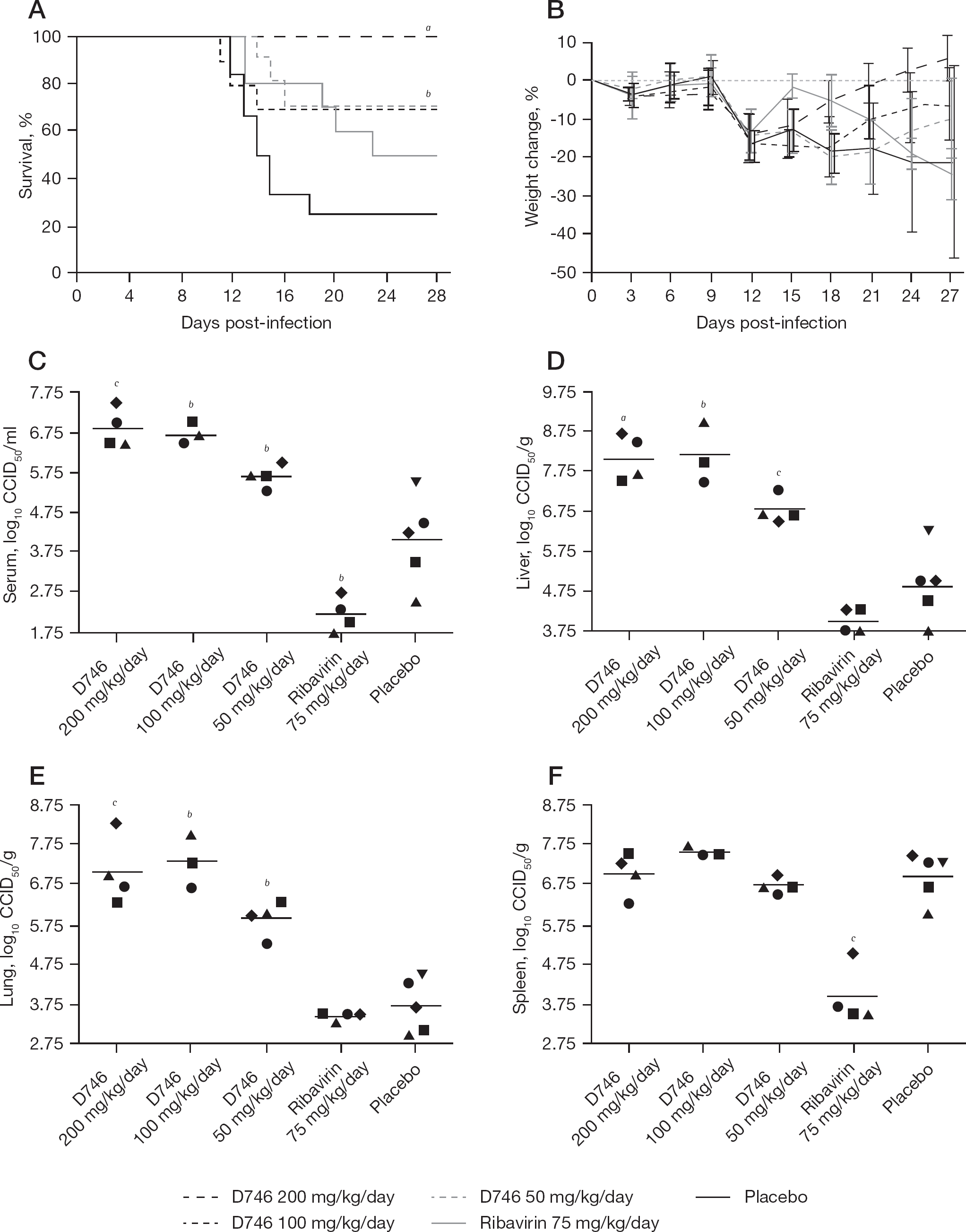
Effect of D746 pretreatment on disease outcome and virus titres in AG129 mice challenged with Tacaribe virus
In general, mean group mouse weights dropped dramatically from day 9 to day 12 (Figure 5B). The mice that received the 200 mg/kg/day dose of D746 started to recover by day 15. Mice treated with the 100 and 50 mg/kg/day doses began to show a collective increase in weight on day 21 and day 24, respectively. The placebo- and ribavirin-treated mice had the greatest mean weight loss by the end of the observation period, with the ribavirin group actually showing a brief improvement prior to a second decrease in animal weights. However, variability in weight change was most pronounced at the end of the experiment due to continued illness in some mice while others were in recovery.
Predetermined cohorts of infected and treated mice were sacrificed on day 9 for analysis of serum and tissue virus titres. Unexpectedly, significantly higher serum, liver and lung titres were present in the D746-treated mice when compared to the placebo-treated animals, and the effect appeared to be dose-dependent; however, there was no effect on spleen titres (Figure 5C–5F). Ribavirin reduced virus titres in the serum, liver and spleen. In general, virus titres in the placebo-treated animals were lower than expected, which was likely due to the disease developing more slowly in this particular experiment. Considering the survival data, the increased virus burden in the mice treated with D746 was unexpected.
Notably, during the sacrifice, we observed large volumes of ascitic fluid and fibrous material surrounding the liver. The effect was dose-dependent, with the mice in the 200 mg/kg/day D746 treatment group clearly having the most ascites fluid accumulation (data not shown). This phenomenon was restricted to the D746 groups as the ribavirin and placebo animals were unremarkable upon opening of the peritoneal cavity for tissue collection. This suggested that the intraperitoneal injections of D746 were causing this fluid accumulation.
Efficacy of D746 as a post-exposure treatment in TCRV-challenged mice
Having confirmed pre-exposure prophylactic efficacy, we next evaluated the ability of D746 to protect TCRV-challenged mice when administered starting 3 days post-exposure. We initiated treatment on day 3 based on previous data suggesting that day 4 is when virus is first observed in the spleen during TCRV infection [13]. This would allow for the evaluation of the compound prior to the appearance of substantial viral titres. Due to limiting amounts of compound, we evaluated D746 at a dose of 100 mg/kg/day. In contrast to the efficacy observed when given pre-exposure, D746 treatment did not prevent mortality when administered after infection (Figure 6A). Only ribavirin was effective post-exposure. Collectively, the ribavirin data from the three mouse experiments suggests that shifting the treatment schedule so that mice receive the drug through day 9 of infection (experiment 3), as opposed to day 6 (experiments 1 and 2), greatly improves survival. It is possible that allowing the immune system to respond to the natural progression of the virus for the initial 3 days resulted in better neutralization of the virus, which combined with ribavirin treatment extending further into the course of infection, protected 90% of the mice from a lethal TCRV challenge.
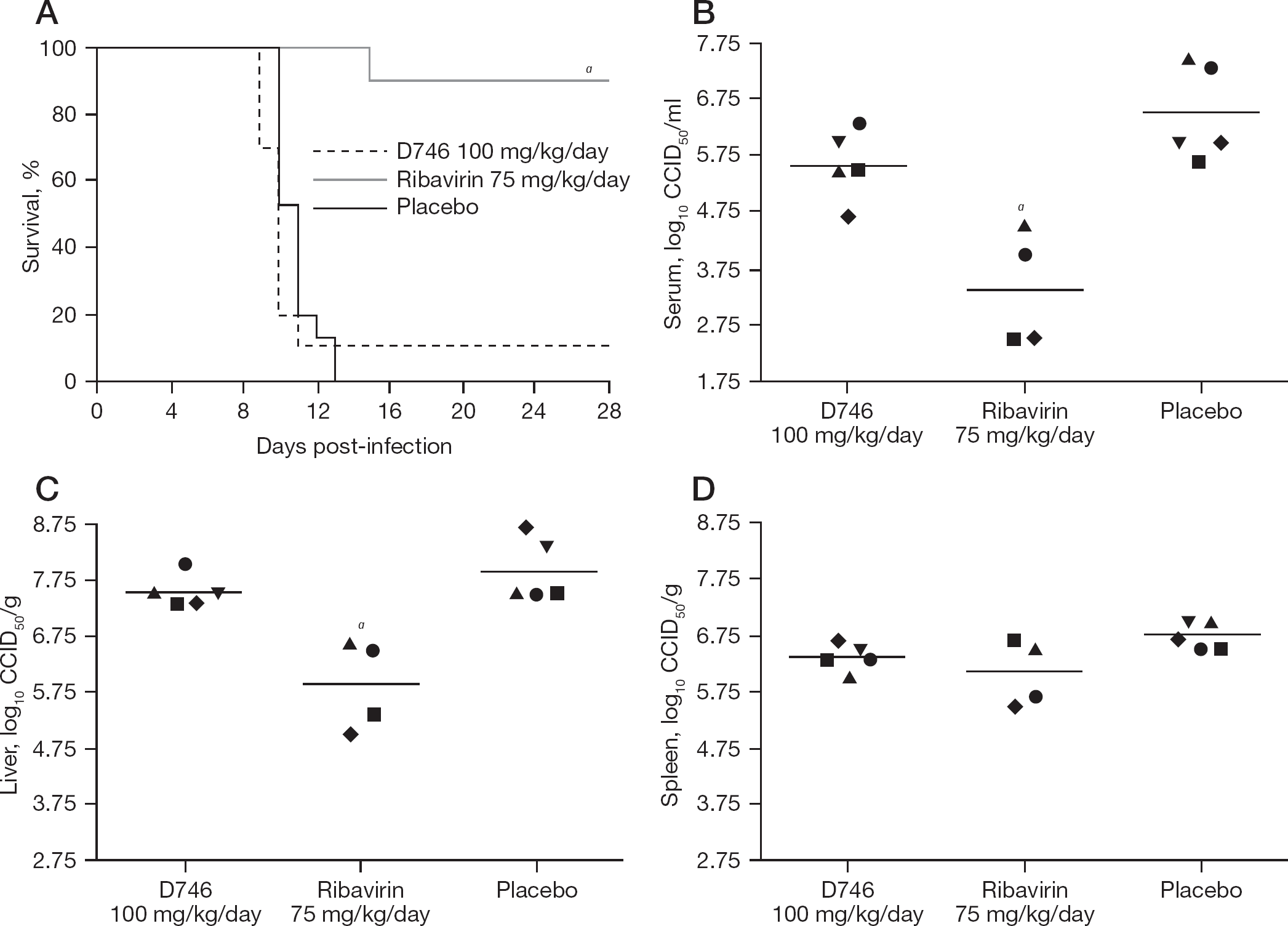
Effect of post-exposure treatment with D746 on survival outcome and virus titres in AG129 mice challenged with Tacaribe virus
The effect of the delayed treatments on reducing viral titres was also evaluated on day 9 in subsets of mice infected and treated in parallel to those observed for mortality. D746 was again ineffective at reducing viral titres; however, in contrast to the second experiment, viral loads in D746-treated groups were not significantly elevated compared to the placebo group (Figure 6B–6D). Consistent with the survival data (Figure 6A), the most dramatic reduction of virus titres was observed with ribavirin, with significant reductions in the serum and liver (Figure 6B and 6C). One death occurred in the ribavirin group on day 8 of infection, prior to the time of sacrifice, resulting in fewer animals for the analysis of serum and tissue virus titres. Because this occurred prior to when we would expect death due to the virus infection, this animal is believed to have died from other complications. Notably, the production of ascites fluid seen in the previous experiment in the D746-treated mice was confirmed during the harvesting of tissues from infected mice. Moreover, uninfected animals also had ascites fluid accumulation in the peritoneum, suggesting that the compound alone is inducing an inflammatory reaction at the site of treatment.
Discussion
The present study describes the identification of a phenolic dibenzylsulfide, D746, with in vitro activity against TCRV and JUNV, and prophylactic activity in a mouse model of TCRV infection. The apparent specificity for arenaviruses in cell culture may be due to inhibition of host cell functions that specifically support arenaviral replication. The precise mechanism by which D746 inhibits replication in cell culture is not clear, but time-of-addition studies indicate that an early to intermediate step in the virus life cycle is being disrupted. In particular, because treatment of cells prior to virus infection caused virus inhibition, this suggests that D746 may attach to cells and block subsequent virus adsorption. Such was the case with a previously reported dibenzamido molecule that inhibited cytomegalovirus adsorption to cells [16]. Further studies would need to be performed with D746 to validate this hypothesis.
Interestingly, despite highly reproducible inhibitory effects in cell culture experiments, the compound failed to reduce viraemia or tissue viral loads in mice. A similar disconnect has been reported previously with the aristeromycin derivative, MY-24, which was able to inhibit viral replication in cell-based arenavirus assays, but not in mice infected with TCRV [13]. Notably, however, MY-24 did not elicit ascites and was effective both as pretreatment and therapeutically when administered starting 5 days after virus challenge. These differences suggest that MY-24 acts in a distinct manner from D746.
In the case of D746, we hypothesize that the peritoneal cavity fluid elicited by the intraperitoneal treatment is indicative of an inflammatory response that may alter the disease process in a way that promotes survival of the mice when treatment begins prior to challenge, despite no reductions on systemic or tissue viral titres measured on day 9 of the infection. This beneficial effect was lost when treatment began after challenge, as there was still evidence of ascites fluid accumulation, but the mice were not protected from lethal TCRV infection. In the second pretreatment efficacy experiment, we actually found significantly higher serum, liver and lung titres in the D746-treated mice compared to the placebo group. We presume that when D746 was given prior to infection the changes resulting from the inflammatory response enhanced the production of virus, perhaps through the recruitment of more specific target cells that amplify the virus, while ameliorating host disease processes that likely contribute to the demise of the animals. When treatment was initiated after TCRV infection, the efficacy of the compound was lost and the increase in virus titres relative to the placebo treatment group was no longer observed. An alternative explanation of the higher titres in the D746-treated mice in the second pre-exposure treatment experiment could be that the placebo group may have had atypically lower viral titres than usual. Unfortunately, we did not have sufficient test compound available to validate whether D746 elevates viral titre under pretreatment conditions. But it is clear that D746 does not reduce viral titres in treated animals.
Presently, there are limited options for the treatment of severe arenaviral infections [1,17]. Despite active research towards identifying new treatments to counter arenaviral HF syndromes, only a limited number of therapies have been evaluated in animal models [18]. Although D746 was efficacious when it was administered prophylactically, it is questionable whether further development of the compound as an anti-arenavirus therapy is warranted since its activity in vitro was only modest, it was only active when administered prior to infection, treatment was not associated with reductions on viral titres, and excess fluid recruited into the peritoneal cavity was linked to intraperitoneal administration of the compound. Nevertheless, the findings reported describe a novel antiviral scaffold and underscore the challenges associated with translating in vitro results into animals.
Footnotes
Acknowledgements
This work was supported by contracts N01-AI-30048, N01-AI-30063 (awarded to Southern Research Institute, Birmingham, AL, USA), HHSN272201100019I and HHSN272201000039I from the Virology Branch, NIAID, NIH.
The authors declare no competing interests.