
Abstract
Background:
Pathogenic hantaviruses geographically distributed in the Old World cause haemorrhagic fever with renal syndrome (HFRS), whereas New World hantaviruses are the aetiological agents of hantavirus cardiopulmonary syndrome (HCPS). Ribavirin, a drug associated with toxicities, is presently indicated for treatment of HFRS, whereas treatment of the more frequently lethal HCPS is limited to supportive care. Because of the need for safe and effective antivirals to treat severe hantaviral infections, we evaluated favipiravir (T-705) against Dobrava and Maporal viruses as representative Old World and New World hantaviruses, respectively. Dobrava virus causes HFRS in Europe. Maporal virus (MPRLV), recently isolated from western Venezuela, is phylogenetically similar to Andes virus, the principal cause of HCPS in Argentina.
Methods:
Hantavirus replication in the presence of various inhibitors was measured by focus-forming unit assays and quantitative reverse transcriptase PCR. Phylogenetic relationships were assessed by the neighbour-joining and bootstrap consensus methods.
Results:
Here, we show that infection of Vero E6 cells with MPRLV is dependent on β3 integrins, similar to that reported for pathogenic hantaviruses. Furthermore, by analysis of molecular determinants associated with the G1 glycoprotein cytoplasmic tail, we show the close genetic proximity of MPRLV to other HCPS-causing hantaviruses in these regions predictive of pathogenicity. We also demonstrate anti-hantavirus activity by favipiravir with inhibitory concentrations ranging from 65 to 93 μM and selectivity indices >50.
Conclusions:
Our data suggest that MPRLV may serve as a safer alternative to modelling infection caused by the highly lethal Andes virus and that hantaviruses are sensitive to the effects of favipiravir in cell culture.
Introduction
Hantaviruses (Bunyaviridae, Hantavirus) are enveloped RNA viruses with tripartite negative- or ambisense genomes that can cause severe and often fatal forms of disease in humans. Pathogenic hantaviruses geographically distributed in the Old World can cause haemorrhagic fever with renal syndrome (HFRS), whereas New World hantaviruses are the aetiological agents of hantavirus cardiopulmonary syndrome (HCPS) [1]. The latter was initially described through an outbreak that occurred in the four corners region of southwestern United States in 1993 [2]. There are at least a dozen hantaviruses known to cause HCPS in North, Central and South America [3–17]. The agents that have received the most attention are Sin Nombre and Andes hantaviruses, which have caused significant disease outbreaks in North and South America, respectively [18]. The high mortality (>40%) associated with this emerging infectious disease, the potential for intentional release of hantaviruses, and the lack of effective antiviral drugs or vaccines available for therapeutic use and prevention stress the importance of developing effective antivirals and treatment modalities.
For some time, the lack of a practical animal model of disease has hindered the development of therapeutics for the treatment and prevention of HCPS. In efforts to study the pathogenesis of HCPS, a hamster disease model based on Andes virus (ANDV) infection was initially established [18]. More recently, a hamster model of infection using a recent isolate, Maporal virus (MPRLV) was described [19]. The virus was isolated in 1997 from central Venezuela and is phylogenetically similar to hantaviruses known to cause HCPS in southern South America [20]. Milazzo and colleagues [19] demonstrated that the MPRLV infection and disease in hamsters parallels that of the ANDV model, which closely resembles human disease. MPRLV-infected animals showed signs of cardiac depression and diffuse pulmonary oedema with rapid onset comparable to findings in clinical cases of HCPS.
Currently, there is no evidence that MPRLV can infect humans and cause disease. Thus, unlike the ANDV hamster infection model that requires maximum containment facilities (biosafety level [BSL]-4) not readily available to most researchers, studies with MPRLV have been performed under less stringent (BSL-3+) containment. In light of the above, the MPRLV hamster infection model may be a more accessible and a suitable surrogate for the study of disease caused by HCPS-causing hantaviruses. Although further characterization of this model will be necessary (especially at the level of immune response and cardiac involvement), initial work by Milazzo and coworkers [19] suggest that it will be a valuable tool for the early stage evaluation of candidate therapeutics to counter HCPS.
The use of integrins as receptors that mediate cellular entry of hantaviruses has been extensively studied [21]. Considerable evidence indicates that pathogenic hantaviruses that cause HCPS and HFRS gain entry via β3 integrins, whereas non-pathogenic hantaviruses utilize β1 integrins [22,23]. β3 integrins participate in the control of vascular permeability [24,25]. Thus, interaction of pathogenic hantaviruses with β3 integrins may disrupt normal function and contribute to the alteration of vascular biology that often leads to severe disease.
Within the hantavirus genome, the M segment is translated into a polyprotein that is cleaved into N-terminal G1 and C-terminal G2 glycoproteins [26,27]. The G1 glycoprotein contains a long cytoplasmic tail that contains a conserved immunoreceptor tyrosine-based activation motif (ITAM) present in all HCPS-causing hantaviruses, but not HFRS or non-pathogenic hantaviruses [28]. Also in the same region of the conserved ITAM motifs, hantavirus G1 cytoplasmic tails can contain a C-terminal hydrophobic domain that directs proteasomal degradation. This so called ‘degron’ sequence directs ubiquitination and degradation of the cytoplasmic tail, which is linked to hantavirus virulence [27,29,30]. The hantavirus degron has been shown to consist of tyrosines that reside within the ITAM (Y619 and Y632), as well as an additional eight residues [28].
Although ribavirin has been shown to have some effectiveness in treating HFRS, it lacks specificity and has toxicity [31]. Recently, the antiviral favipiravir (T-705; 6-fluoro-3-hydroxy-2-pyrazinecarboxamide) has been reported to be active against a number of bunyaviruses both in vitro and in vivo in mouse and hamster models [32,33]. Notably, favipiravir has also been shown to be effective in treating severe influenza virus, arenavirus and flavivirus infections in animal models [34–39]. Because favipiravir is active against other members of the Bunyaviridae family, we hypothesized that it would also inhibit hantavirus replication. Thus, in the present study, we investigated MPRLV β integrin utilization and genetic determinants of pathogenicity, and evaluated hantavirus sensitivity to the effects of favipiravir in cell culture.
Methods
Cells and viruses
The monkey kidney cell line Vero E6 was purchased from American Type Culture Collection (ATCC; Manassas, VA, USA). Cells were maintained in minimal essential medium supplemented with 10% fetal bovine serum (FBS) and gentamicin (Sigma, St Louis, MO, USA). All media and serum were from HyClone Laboratories (Logan, UT, USA) unless otherwise stated. Prospect Hill virus (PHV; strain MP40 TVP6042), Dobrava virus (DOBV; strain Sotkamo) and MPRLV (strain HV9021050) were provided by Robert Tesh (World Reference Center for Emerging Viruses and Arboviruses, University of Texas Medical Branch, Galveston, TX, USA). All three viruses were propagated in Vero E6 cells and their identity confirmed by quantitative reverse transcriptase (RT)-PCR (KKB et al., data not shown). Infections were performed in Dulbecco's modified essential medium (DMEM) with 2% FBS and gentamicin. BSL-3 facilities were used for studies with DOBV and MPRLV, whereas BSL-2 containment was used for work with PHV.
Ligands, antibodies and staining reagents
Sin Nombre virus (SNV) hyperimmune mouse serum (kindly provided by Robert Tesh) was used as the primary anti-hantavirus antibody for focus-forming unit (FFU) detection. The secondary antibody used was a goat anti-mouse horseradish peroxidase conjugated antibody from Kirkegaard & Perry Laboratories (KPL; Gaithersburg, MD, USA). Both were used at a 1:500 dilution. Polyclonal function-blocking antibodies directed at β1 and β3 integrins for receptor usage determination studies were purchased from Millipore (Chemicon, Temecula, CA, USA) and titrated for use at 1 μg/ml. Fibronectin and vitronectin were purchased from Sigma and titrated for use at concentrations of 20 and 1 μg/ml, respectively. Acidin and biotinylated horseradish peroxidase complex (ABC) and 3,3′-diaminobenzidine (DAB) staining kits were purchased from Vector Laboratories (Burlingame, CA, USA).
Focus-forming unit assay
Vero E6 cells (approximately 80% confluent) in 96-well plates were pretreated with known β integrin ligands or function-blocking antibodies for 1 h at 37°C before viral infection. Ligands or antibodies were removed and 100 μl volumes containing 1,000 FFUs of MPRLV or 4,000 FFUs of DOBV or PHV were adsorbed onto the pretreated cells at 37°C, 5% CO2 for 90 min. After adsorption, virus inocula were removed, and the cells were incubated in DMEM with 2% FBS and gentamicin an additional 24 h for MPRLV, 3 days for DOBV and 5 days for PHV prior to staining. All cell cultures were confluent at the time of immunoperoxidase staining.
Immunoperoxidase staining of the nucleocapsid protein in infected cells has been previously described [23]. In brief, cell monolayers were fixed with 100% methanol at 4°C, for 10 min, permeabilized with 0.25% Triton X-100 in PBS for 10 min and blocked with 4% goat serum in 1% bovine serum albumin in PBS for 1 h. Monolayers were incubated with SNV hyperimmune mouse serum (1/500) for 1 h at 37°C and subsequently with goat anti-mouse horseradish peroxidase conjugated antibody (1/500). Infected cell foci were quantified after staining with ABC and working DAB solution as per the manufacturer's instructions.
Phylogenetic analysis
Amino acid sequences from the GenBank database were compared using Molecular Evolutionary Genetics Analysis (MEGA) software (Center for Evolutionary Medicine and Informatics, Tempe, AZ, USA). Analysis of phylogenetic relationships were made using the neighbour-joining and bootstrap consensus methods [40] analysing only the ITAM/degron 30 amino acid region [28,29] of the M segment of the genome. The evolutionary distances were computed using the Poisson correction method and are in the units of the number of amino acid substitutions per site [41]. All positions containing gaps and missing data were eliminated from the dataset (complete deletion option).
Hantavirus M segment GenBank sequence accession numbers were as follows: Andes (AAO86638.1), Bayou (AAA61690.1), Bermejo (AAB87911), Choclo (ABB90558.1), Dobrava (AAY27875.1), Hantaan (AAK27683.2), Laguna Negra (AAB87603), Maporal (AAR14889.1), New York-1 (AAC54560), Prospect Hill (CAA38922.1), Puumula (AB297666.2), Seoul (NP_942557.1), Sin Nombre (AAG03036.1) and Tula (NP_942586).
Drug efficacy studies
Favipiravir was provided by the Toyama Chemical Company (Toyama, Japan). Ribavirin was obtained from ICN Pharmaceuticals (Costa Mesa, CA, USA). The antiviral activity of favipiravir and ribavirin was determined as follows: Vero E6 cells (approximately 80% confluent) in 96-well plates were infected with hantavirus FFUs as specified above. MPRLV and DOBV inocula were adsorbed for 1 h and PHV was absorbed for 6 h. After infection, virus was removed and cells were treated with eight serial half-log10 dilutions of the drugs. Plates were incubated (37°C, 5% CO2, DMEM with 2% FBS and gentamicin) for 24 h for MPRLV, 3 days for DOBV and 5 days for PHV. To determine cell cytotoxicity, drugs were added to uninfected parallel cultures and cell viability was determined at 24 h, 3 days or 5 days (consistent with the various hantavirus assay time requirements) by neutral red dye uptake, as previously described [32]. Toxicity was presented as the 50% cell cytotoxic concentration (CC50), defined as the concentration of the drug that resulted in a 50% decrease in cell viability compared with the cell controls. Drug efficacy was determined by quantification of infected cells and determining the 50% inhibitory concentration (IC50), defined as the concentration of the drug required to reduce viral FFUs by 50%. The selectivity (or therapeutic) index (SI) is defined as the ratio of the CC50 to the IC50.
Quantitative (q)RT-PCR was also used to determine drug efficacy against MPRLV by measuring relative viral RNA concentrations after incubation of infected cells with the drugs. Triplicate wells of a 24-well plate containing approximately 80% confluent Vero E6 cultures were infected with 250 μl containing 300 FFU of MPRLV for 6 h, at which time the virus inocula were removed and the cells treated with serial dilutions of ribavirin or favipiravir. After 3 days in culture, RNA was collected using an RNeasy total RNA purification kit (Qiagen, Valencia, CA, USA) following the manufacturer's recommendations.
The MGB Eclipse probe system from Epoch Biosciences (Bothell, WA, USA) was used in combination with the SuperScript III Platinum One-Step Quantitative RT-PCR system from Invitrogen (Carlsbad, CA, USA), as recommended by the manufacturers for the qRT-PCR reactions. The MPRLV primers and probe combination were designed using the MGB Eclipse Online Design software and targeted the nucleocapsid coding sequence. The forward primer was 5′-GGA CAT TTC CAT AAC GCA GTG-3′ and the reverse primer was 5′-TGG CAG CTC AGA AAC TGG CTT CAA A-3′. The probe target sequence was (MGB EDQ)-5′-GTC ATC AGG TTC AAG C-3′-(FAM).
Viral RNA levels were assayed in duplex with, and normalized to, β2-microglobulin expression. All reactions were performed using the DNA Engine Opticon 2 Real-Time PCR detection system (Bio-Rad, South San Francisco, CA, USA). The IC50 was calculated as the dose of drug reducing the relative MPRLV RNA levels by 50% compared with infected untreated controls. At the lower concentrations of compounds tested, cytotoxicity was not observed in parallel cultures that were not infected with MPRLV (KKB et al., data not shown).
Results
β3 Integrin chain-specific antibodies and vitronectin block MPRLV infectivity
β3 Integrin usage is a characteristic feature shared by pathogenic hantaviruses. To assess MPRLV integrin usage, Vero E6 cell monolayers were treated with antibodies or integrin antagonists to block hantavirus infectivity. Following incubation, infected cells were quantified by FFU assay and experimental treatment groups were compared with untreated virus-infected controls. PHV infection was significantly inhibited by anti-β1-specific integrin antibodies, but not anti-β3 integrin antibodies (Figure 1A). Conversely, pretreatment of Vero E6 cells with polyclonal antibodies to β3 integrins significantly decreased MPRLV and DOBV infectivity.
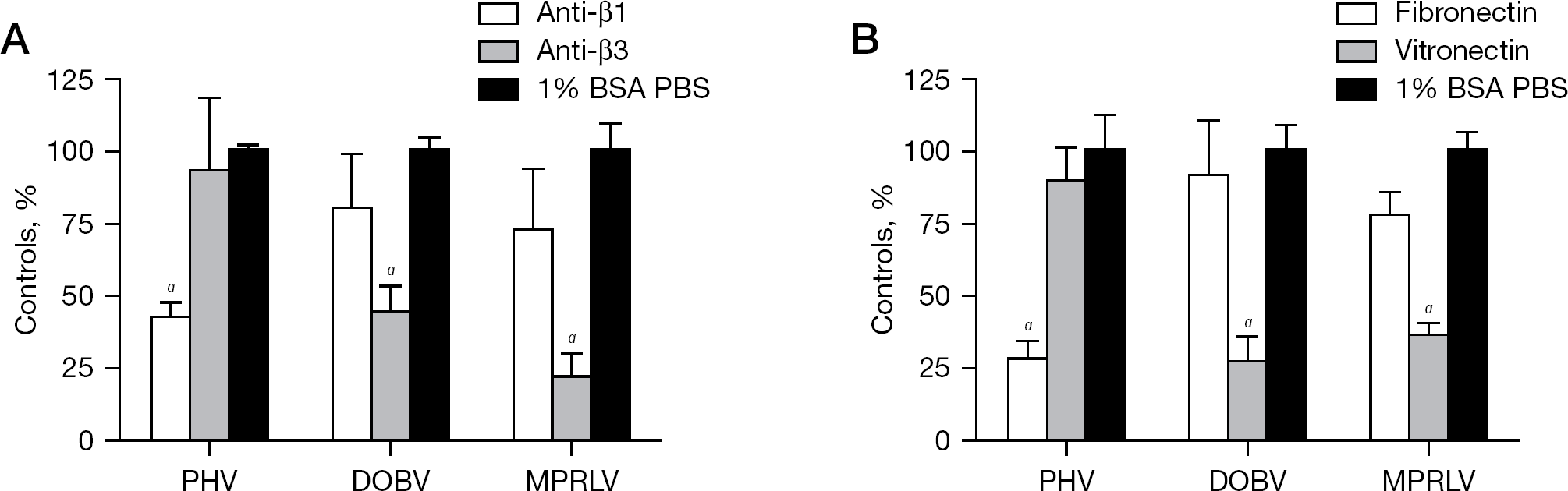
MPRLV infection is mediated by β3 integrins
We next assayed the ability of specific integrin ligands to inhibit MPRLV infectivity. Vitronectin and fibronectin are extracellular matrix (ECM) proteins with high affinity for αVβ3 and α5β1 integrins, respectively. Consistent with the antibody studies, pretreatment of Vero E6 cells with vitronectin significantly reduced MPRLV and DOBV infectivity. By contrast, PHV infection was significantly reduced with fibronectin (Figure 1B). Collectively, these data suggest that MPRLV host cell infection is facilitated principally by β3 integrins, similar to the known pathogen DOBV.
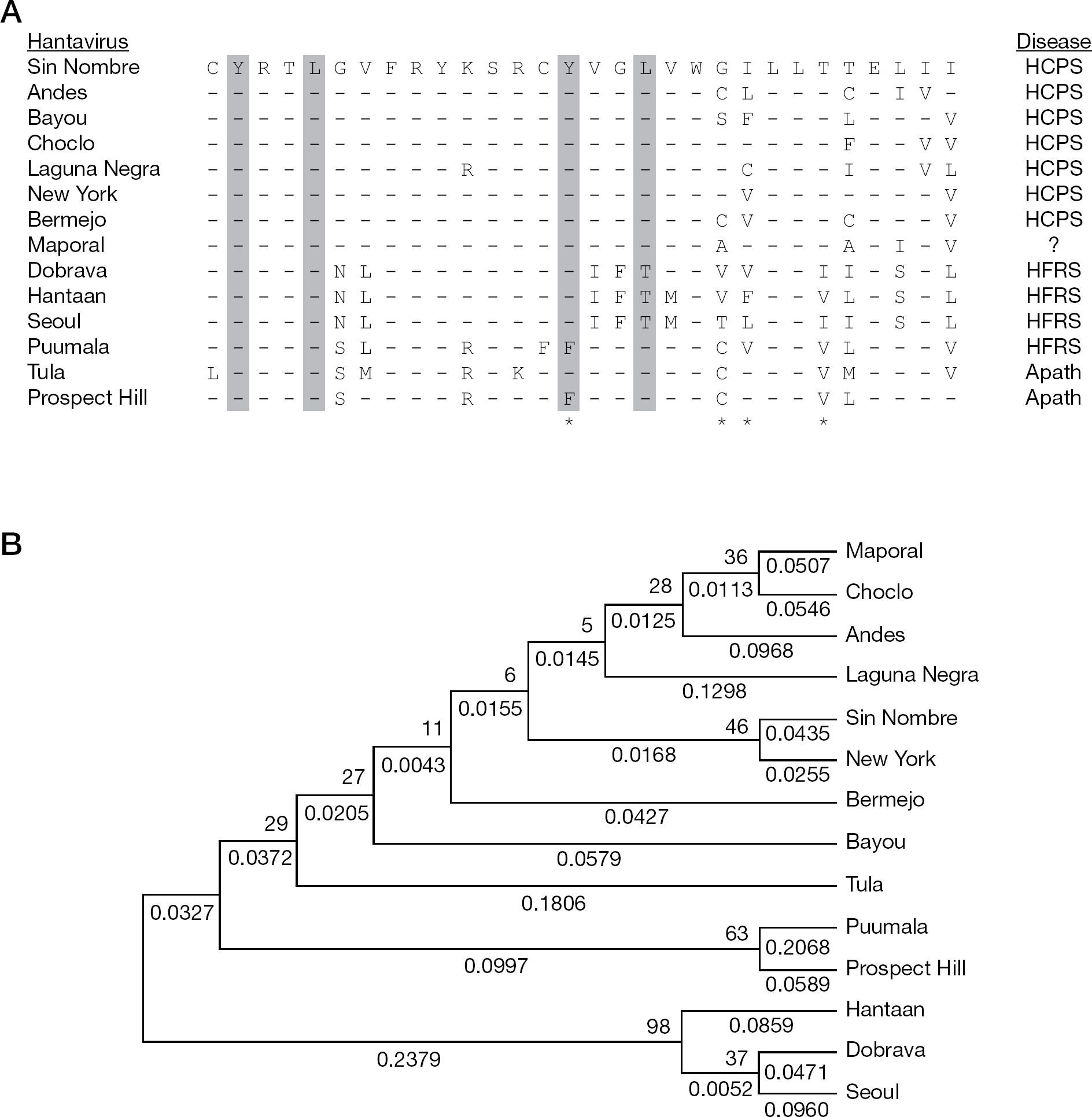
Amino acid comparison and phylogenetic relationships among the ITAM/degron sequences of 14 different hantaviruses
MPRLV shares ITAM and degron sequence homology with pathogenic hantaviruses
We performed a phylogenetic analysis to compare ITAM and degron protein sequences of 13 hantaviruses with that of MPRLV as a means to obtain insights into potential pathogenicity of MPRLV. An alignment of hantavirus sequences obtained from GenBank that contain the overlapping ITAM and degron regions is shown in Figure 2A. This ClustalW alignment indicated that the G1 cytoplasmic tail of MPRLV contained the characteristic ITAM sequence found in all HCPS-causing hantaviruses. The comparison also indicated that MPRLV does not contain the conserved residues that stabilize the G1 tail in non-pathogenic hantaviruses, but does contain residues consistent with pathogenic hantaviruses (Figure 2A). Phylogentic analysis restricted to the 30 amino acid sequence encompassing the ITAM/degron region shows that MPRLV is more closely related to the HCPS-causing hantaviruses (Figure 2B). Notably, similarity of this region with ANDV was striking.
Efficacy of favipiravir and ribavirin against MPRLV
The inhibitory activity of favipiravir and ribavirin against hantaviruses was determined by infectious FFU assay (Table 1). Ribavirin, known to be active against HFRS- and HCPS-causing hantaviruses in vitro [42–44], inhibited MPRLV infectivity in a dose-dependant manner, with an IC50 of 47 μM and therapeutic index of 22. As expected, ribavirin was also active against DOBV and PHV. Favipiravir, reported to be effective at inhibiting replication of related bunyaviruses [32], was also found to be similarly active against MPRLV with an IC50 of 65 μM and therapeutic index of 74. It was also effective against DOBV and PHV.
In vitro inhibitory effects of favipiravir and ribavirin against hantaviruses by FFU assay a
Data are the mean and standard deviations from 3–4 separate experiments in Vero E6 cells.
50% cytotoxic concentration (CC50) and 50% virus inhibitory concentration (IC50) values are in μM.
Selectivity index (SI) = ratio of CC50 to IC50. DOBV, Dobrava virus; FFU, focus-forming units; MPRLV, Maporal virus; PHV, Prospect Hill virus.
A qRT-PCR assay was also employed to evaluate the anti-MPRLV activities of favipiravir and ribavirin. As shown in Figure 3, favipiravir activity was verified by the reduction of MPRLV RNA (IC50 of 33 μM). Ribavirin also reduced MPRLV RNA levels in a dose-dependent manner with an IC50 of 22 μM (Figure 3). As observed with the FFU assay, the IC50 of ribavirin was slightly less compared with the IC50 of favipiravir by the qRT-PCR method. Notably, the qRT-PCR assay generally resulted in lower IC50 μM ranges. Presumably, this difference resulted from differences in assay formats (96-well versus 24-well plates), the duration of the assays and the lower multiplicity of infection.
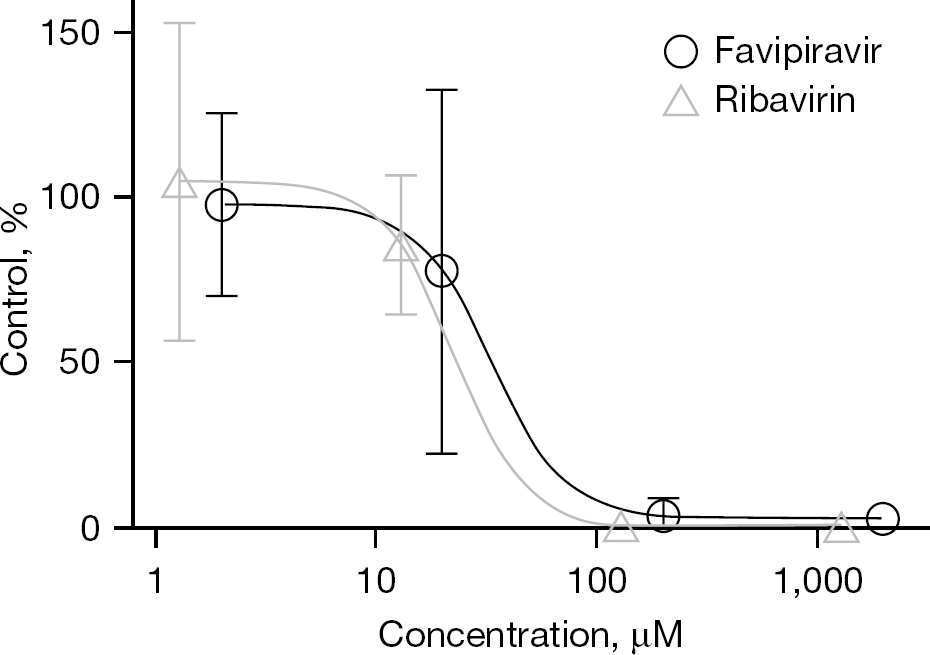
Favipiravir reduces MPRLV RNA during infection
Discussion
Our findings suggest that MPRLV infectivity in Vero cells is principally mediated by β3 integrins. To date, only pathogenic hantaviruses have been demonstrated to ultilize β3 integrins for gaining entry into host cells, whereas the non-pathogenic hantaviruses require β1 integrins [22,23]. αvβ3 Integrins help maintain vascular integrity through interaction with extracellular matrix and participate in the regulation of vascular permeability [24,25,45,46]. Alteration of αvβ3 integrin function through interaction with pathogenic hantaviruses may contribute vascular dysregulation associated with severe forms of disease. The preponderance of evidence suggests that β3 integrin usage by hantaviruses is highly predictive of their ability to cause disease in humans.
In our studies characterizing MPRLV, pretreatment of cells with function-blocking β3 integrin antibodies and vitronectin significantly reduced infectivity. Consistent with previous work by others [22], anti-β3 integrin and vitronectin also inhibited the HFRS-causing DOBV. Notably, complete inhibition could not be achieved, suggesting that other mechanisms of virus entry are likely. Recent studies have shown that in addition to the β integrins, hantaviruses also exploit decay-accelerating factor (DAF) to successfully infect cells [47,48]. This may explain why, after incubation with ECM proteins or antibodies to the β integrins, viruses still were able to infect cells with ‘blocked’ receptors resulting in less than complete inhibition. Studies blocking both β integrin and DAF proteins would likely result in more complete blockage of hantaviral infection. Partial inhibition of pathogenic hantavirus replication following treatment of cells with β1 integrin-specific blocking antibodies or fibronectin may be attributed to steric hindrance of viral binding, as suggested in experiments with West Nile virus [49].
In addition to integrin utilization pattern as a predictor of pathogenicity, several motifs present in the cytoplasmic tail of the G1 glycoprotein of hantaviruses can also serve as distinguishing features. Our comparative analysis focused on the stretch of amino acids encompassing both the ITAM and degron sequences demonstrate extensive similarity between MPRLV and other HCPS-causing hantaviruses. ITAM motifs participate in intracellular signalling that directs cellular mechanisms. Viruses may be able to regulate ITAM signalling responses of host cells [29]. The presence of such motifs almost exclusively in pathogenic HCPS-causing hantaviruses may suggest an additional mechanism by which virulence and host cell function can be regulated by the virus. Notably, the apathogenic hantavirus, Tula virus (TULV), contains the ITAM sequence. However, its G1 tail is missing an adjacent cysteine residue conserved in all other hantaviruses that likely alters the conformation of the ITAM in the TULV G1 tail [28].
The degron sequence is a C-terminal hydrophobic domain of the G1 cytoplasmic tail that directs proteasomal degradation and is also linked to the virulence of hantaviruses. A recent study comparing differences between PHV and the New York-1 hantavirus (NYV) identified a series of four amino acid residues that are crucial to the unstable state of the NYV G1 tail [27]. In MPRLV, like other pathogenic hantaviruses, the C-terminal hydrophobic region of the G1 cytoplasmic tail has interspersed hydrophilic residues. Replacement of hydrophobic amino acids in this region of the non-pathogenic PHV with polar residues results in its destabilization [27]. Experiments investigating tail-directed proteasomal degradation of the MPRLV and other pathogenic and non-pathogenic hantavirus G1 cytoplasmic tails would further support the hypothesis that MPRLV may emerge as a human pathogen.
In our phylogenetic analysis restricted to the ITAM and degron sequences, Puumala virus (PUUV), a hantavirus that causes a mild form of HFRS [50,51], segregates with the apathogenic PHV. Phylogenetic analysis not only serves to reveal relationships between hantaviruses that cause the same disease, but also a correlation with the phylogeny of their rodent hosts [52,53]. To this end, the sharing of rodents hosts, such as that of PHV and PUUV (Arvicolinae), likely drives these associations. Notably, MPRLV had significant phylogenetic similarities with pathogenic and, more specifically, HCPS-causing hantaviruses such as SNV and ANDV.
In the present study, ribavirin and favipiravir were evaluated against MPRLV and found to be active by FFU and qRT-PCR assays measuring infectious virus and viral RNA, respectively. DOBV was also found to be sensitive to the effects of favipiravir. We hypothesize that the mechanism by which favipiravir inhibits hantavirus replication is through inhibition of the viral polymerase, as has been suggested for influenza virus and arenaviruses [54,55]. However, functional assays for hantavirus RNA-dependent RNA polymerases are not presently available to thoroughly investigate this proposed mode of action. The 65–93 μM inhibitory range observed was consistent with previously reported data for other members of the Bunyaviridae family, including the Punta Toro phlebovirus [32,33]. More importantly, favipiravir has been demonstrated to be highly efficacious in the mouse and hamster Punta Toro virus infection models [32,33]. It is hypothesized that the remarkable in vivo activity may be due to increased activity or abundance of the host cellular enzymes that covert favipiravir into the active triphosphate form of the drug.
Favipiravir is presently in clinical trials in Japan (Phase III) and the United States (Phase II) for the treatment of influenza virus infections, and so the safety of the compound is being comprehensively addressed. Based on the well-documented activity of favipiravir in mouse and hamster models of Punta Toro virus infection and the IC50 values for this related bunyavirus ranging from 55 to 191 μM [32,33], we would predict efficacy in hantavirus infection models. Collectively, our findings support future preclinical development of favipiravir in the hamster MPRLV or ANDV infection models [18,19] or the mouse Hantaan virus infection model [56,57] as proof-of-concept for in vivo efficacy.
Footnotes
Acknowledgements
This work was supported by contract N01-AI-30048 from the Virology Branch, National Institute of Allergy and Infectious Diseases, National Institutes of Health.
YF is an employee of the Toyama Chemical Co., Ltd., the manufacturer of T-705. All other authors declare no competing interests.