
Abstract
Background:
The pathogenesis of severe acute respiratory syndrome coronavirus (SARS-CoV) is poorly understood. Several mechanisms involving both direct effects on target cells and indirect effects via the immune system might exist. SARS-CoV has been shown in vitro to induce changes of cytokines and chemokines in various human and animal cells. We previously reported that interferon (IFN) alfacon-1 was more active against SARS-CoV infection in human bronchial epithelial Calu-3 cells than in African green monkey kidney epithelial cells on day 3 post-infection.
Methods:
In the current study, we first evaluated the efficacy of IFN-alfacon 1 in Calu-3 cells during the first 7 days of virus infection. We then used the two-antibody sandwich ELISA method to detect IFN-γ-inducible protein 10 (IP-10). We further evaluated the efficacy of antivirals directed against SARS-CoV infection in BALB/c mice.
Results:
A potent, prolonged inhibition of SARS-CoV replication in Calu-3 cells with IFN-alfacon 1 was observed. Furthermore, IP-10, an IFN-inducible leukocyte chemoattractant, was detected in Calu-3 cells after SARS-CoV infection. Interestingly, IP-10 expression was shown to be significantly increased when SARS-CoV-infected Calu-3 cells were treated with IFN alfacon-1. IP-10 expression was detected in the lungs of SARS-CoV-infected BALB/c mice. Significantly high levels of mouse IP-10 in BALB/c mice was also detected when SARS-CoV-infected mice were treated with the interferon inducer, polyriboinosinic-polyribocytidylic acid stabilized with poly-L-lysine and carboxymethyl cellulose (poly IC:LC). Treatment with poly IC:LC by intranasal route were effective in protecting mice against a lethal infection with mouse-adapted SARS-CoV and reduced the viral lung titres.
Conclusion:
Our data might provide an important insight into the mechanism of pathogenesis of SARS-CoV and these properties might be therapeutically advantageous.
Introduction
Chemoattractant cytokine (chemokine) receptors are a group of membrane proteins; they belong to the superfamily of G-protein-coupled receptors (GPCRs) that possess seven transmembrane helices and transmit signals from extracellular ligands to intracellular biological pathways via heterotrimeric G-proteins [1–3]. Chemokines are a family of small secreted proteins of 70–80 residues that direct the recruitment of leukocytes to sites of inflammation [4]. Based on the positions of two conserved cysteine residues in their N-termini, chemokines can be divided into four subfamilies: CC, CXC, CX3C and C [5,6]. The two chemokine main subfamilies are CXC and CC. They are important for the selective activation and recruitment of a large variety of cell types in inflammation. Interferon (IFN)-γ-inducible protein 10 (IP-10), also known as CXCL10, is a member of a subfamily of CXC chemokines. IP-10 is a chemoattactant that signals via binding and activation of the seven transmembrane G-protein-coupled receptor CXCR3, a CXC chemokine receptor that is expressed in T-lymphocytes. The epithelial cells play an important role in this process.
In 2003, a coronavirus was identified as the causative agent of an emerging human infectious disease: severe acute respiratory syndrome (SARS). SARS posed a new threat for respiratory medicine and represented a challenge for antiviral drug development and administration [7,8]. Its immunopathological mechanisms have not yet been fully characterized. The clinical progression contains three phases: active viral infection, hyperactive immune response, and recovery or pulmonary destruction and death. One hypothesis is that the pathogenesis of SARS coronavirus (SARS-CoV) is caused by a disproportionate immune response, illustrated by increased levels of inflammatory cytokines and chemokines, such as IP-10, monocyte chemoattractant protein-1 (MCP-1; CCL2), interleukin 6 (IL-6) and interleukin 8 (IL-8) [9,10]. Chemokines are involved in the recruitment of leukocytes into sites of tissue inflammation [11]. SARS-CoV has been shown in vitro to induce changes of cytokines and chemokines in various human and animal cells [12,13].
In a recent study, we evaluated a few compounds approved for therapeutic use in humans and some in vitro inhibitors of SARS-CoV for inhibition in the mouse SARS-CoV replication model [14]. We found IFNs and IFN inducers to inhibit virus replication. A hybrid IFN, IFN-α B/D, and a mismatched double-stranded RNA IFN inducer, ampligen (poly I:poly C124), were shown to potently inhibit virus titres in the lungs of infected mice [14]. In another report, we described a cell-based assay using SARS-CoV infection of human lung epithelial cells (Calu-3) and three African green monkey kidney epithelial cell lines to evaluate IFN alfacon-1 (Infergen™) against SARS-CoV. IFN alfacon-1 was shown to be more active against SARS-CoV when tested in Calu-3 cells than in African green monkey kidney epithelial cells on day 3 post-infection [15]. In the current study, we evaluated the efficacy of IFN alfacon-1 in Calu-3 cells during the first 7 days of virus infection. A potent, prolonged inhibition of SARS-CoV replication in Calu-3 cells with IFN alfacon-1 was observed. Furthermore, human IP-10 was detected in Calu-3 cells after SARS-CoV infection. Meanwhile, human IP-10 expression was shown to be significantly increased when SARS-CoV-infected Calu-3 cells were treated with IFN alfacon-1. Because small animal models are needed in order to evaluate the efficacy of antivirals directed against SARS-CoV infection, we investigated the ability of SARS-CoV to infect the BALB/c mice and discovered that mouse IP-10 was increased in the lungs of SARS-CoV-infected BALB/c mice. Significantly high levels of mouse IP-10 in BALB/c mice was also detected when SARS-CoV-infected mice were treated an IFN inducer – a synthetic, double-stranded polyriboinosinic-polyribocytidylic acid stabilized with poly-L-lysine and carboxymethyl cellulose (poly IC:LC).
Methods
Cells
Calu-3 cells are a human lung epithelial cell line originating from a human pulmonary adenocarcinoma and were grown in Dulbecco's modified Eagle's minimal essential medium supplemented with 20% fetal bovine serum (FBS [D-20]; Hyclone Laboratories, Logan, UT, USA) [15,16]. Vero 76 cells, which were obtained from American Type Culture Collection (Manassas, VA, USA) [17], are an African green monkey kidney epithelial cell line and were routinely grown in minimal essential medium (MEM) supplemented with 10% heat-inactivated FBS. For antiviral assays, the serum was reduced to 10% in Calu-3 cells and 2% in Vero 76 cells, and gentamicin was added to the medium at a final concentration of 50 μg/ml.
Virus
SARS-CoV, strain Urbani (200300592), was obtained from the Centers for Disease Control and Prevention (Atlanta, GA, USA). Urbani strain was propagated and titrated in Vero 76 cells.
Mouse-adapted virus
The mice were infected with Urbani strain of SARS-CoV. Three or 5 days after infection, the lungs were removed, homogenized and then re-infected into a subsequent group of mice. This infection step continued for 25 times through mouse lungs. The virus was then plaque purified three times and yielded a virus causing severe lung disease and mortality in infected mice.
Test compounds
Stock solutions of IFN alfacon-1 (InterMune, Inc., Brisbane, CA, USA) were provided at a concentration of 15 μg/0.5 ml and stored at −20°C. IFN alfacon-1 was solubilized in MEM for in vitro experiments at a final concentration of 3.0 ng/ml. IFN inducer, poly IC:LC, was obtained from Ribopharm Corporation (Bethesda, MD, USA).
Infectious virus titration assay
Calu-3 cells were seeded to the 24-well tissue culture plates (Corning Incorporated Costa, NY, USA) at 4×105 cells/ml 1 day before and treated with IFN alfacon-1 at the different concentrations 60 min prior to the addition of virus. IFN alfacon-1 was removed and the cells were then infected with SARS-CoV at the multiplicity of infection equal to 0.001 and IFN alfacon-1 was added back to the cells. IFN alfacon-1 and virus were added in equal volumes (0.1 ml). After a 4 h attachment period, the supernatant was removed, the monolayers rinsed once, and fresh Dulbecco's modified Eagle's minimal essential medium with or without IFN alfacon-1 and supplemented with 10% FBS was added back to each well. The plates were incubated at 37°C for 1–7 days. Infected supernatant fluids from Calu-3 cells were harvested daily and frozen at −80°C for subsequent virus titration by cytopathic effect assay in Vero 76 cells. Three assays were performed and each test was done in triplicate.
Assay for human IP-10
Calu-3 cells (4×105 cells/ml) were seeded to 24-well tissue culture plate. Next day, the cells were treated with IFN alfacon-1 and then infected with SARS-CoV Urbani strain at the multiplicity of infection equal to 0.001. Incubation was continued for 7 days. The medium was collected during incubation and stored at −80°C until use. Secreted IP-10 concentrations were determined by ELISA. We used the two-antibody sandwich method (R. Briefly, samples (30× diluted with dilution buffer) were applied to 96-well polystyrene microplate, which had been coated with mouse monoclonal antibody against IP-10. Recombinant human IP-10 was used as a standard. Then, polyclonal antibody against IP-10 conjugated to horseradish peroxidase was added to each well. Colour reagent A and B (hydrogen peroxide and chromogen) were used as a substrate and the reaction was stopped by adding 2 N sulfuric acid. The plate was read at 450 nm wavelength with a microplate reader (Opsys MR™, Dynex Technologies, Chantilly, VA, USA). The minimum detectable dose was determined by adding two standard deviations to the mean opitical density value of 20 zero standard replicates and calculating the corresponding concentration. Three assays were performed and each test was done in triplicate.
Animal studies
Animal studies were done to evaluate SARS-CoV replication in the female 18–20 g specific pathogen-free BALB/c mice from Charles River Laboratories (Wilmington, MA, USA). The BALB/c mice were housed in a biosafety level 3+ animal facility, and fed standard mouse chow and tap water ad libitum. The mice were anesthetized with a 0.1 ml intraperitoneal injection of 20 mg/kg of ketamine® and 0.05 ml SARS-CoV was administered intranasally. Prior to poly IC:LC treatment, a dose–range experiment was done to determine the maximum tolerated concentration using dosage regimens that were to be used in the actual efficacy studies. Ten mice per group were pre-treated 24 h prior to infection by intranasal injection administration with poly IC:LC at 5 mg/kg. On day 0, these 10 mice received the second dose of poly IC:LC 8 h after exposure to virus and received the third dose 24 h after the second dose. Another 10 mice per group received the first dose of poly IC:LC by intranasal administration 8, 16, 24, 48 and 72 h after exposure to virus. Following intranasal administration of SARS-CoV, five mice from each group were sacrificed on day 3 and 6. Each mouse lung was collected and homogenized, and the samples were assayed in triplicate for viral titration in Vero 76 cells by cytopathic effect assay; 50% cell culture infectious dose values were calculated. SARS-inoculated and mock-infected mice were weighed every day and observed for the clinical signs of disease; both weight loss and clinical signs were observed in the infected groups.
Determination of viral titres
Lung samples were homogenized in MEM supplemented with 10% FCS and gentamicin (Invitrogen, NY, USA). Tissue homogenates were clarified by low-speed centrifugation, and the virus titres were determined in Vero 76 cells in 96-well plate as described previously.
Results
Potent, prolonged inhibition of SARS-CoV replication in Calu-3 cells with IFN alfacon-1 during 7 days of infection
The effects of IFN alfacon-1 on the replication of SARS-CoV in Calu-3 cells at various times after virus exposure were compared with infected cells not treated with IFN alfacon-1. Released virus titres were evaluated in Vero 76 cells. The SARS-CoV virus replicated in high numbers in Calu-3 cells (4.8 log10 50% cell culture infectious dose per 0.18 ml, Figure 1). In Calu-3 cells, virus production titres peaked on day 5 in untreated samples and on day 4–6 for treated samples. IFN alfacon-1 markedly inhibited extracellular virus production in Calu-3 cells from days 1–3 at a dose previously found to reduce virus titres by 90% at day 3 (3.0 ng/ml) [15]. The inhibition of extracellular virus from Calu-3 cells was nearly total from days 1–7. IFN alfacon-1 at 3.0 ng/ml almost completely inhibited viral replication except for a few random wells within some of the replicates on day 6. At day 6, some virus, albeit a very modest amount, was produced, overcoming the total inhibition on infectious virus production induced in response to the exogenous IFN at other time periods. Lower concentrations of IFN alfacon-1 slowed replication and reduced the virus titres compared with untreated controls at most time points. The 90% effective concentration for IFN alfacon-1 on day 3 of this study was 0.49 ±0.56 ng/ml, which was higher than the 0.046 ±0.011 ng/ml observed in previous work [15]. This difference was likely due to the addition of 10% FBS to the medium rather than 2% FBS, which might affect production of antiviral chemicals stimulated by IFN alfacon-1.
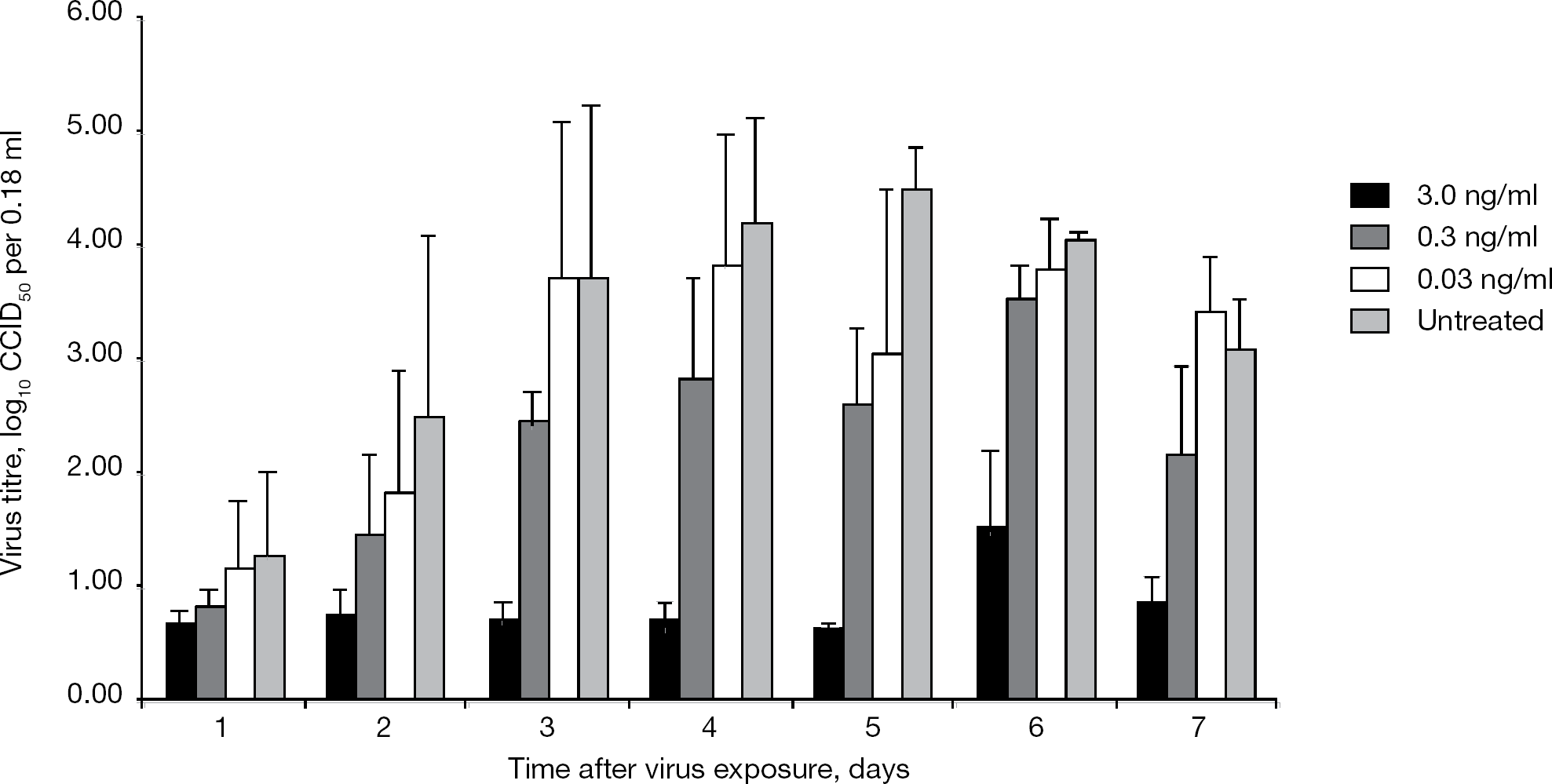
Growth of SARS-CoV in human lung adenocarcinoma Calu-3 cells with and without various concentrations of IFN alfacon-1
SARS-CoV can induce human IP-10 expression in Calu-3 cells
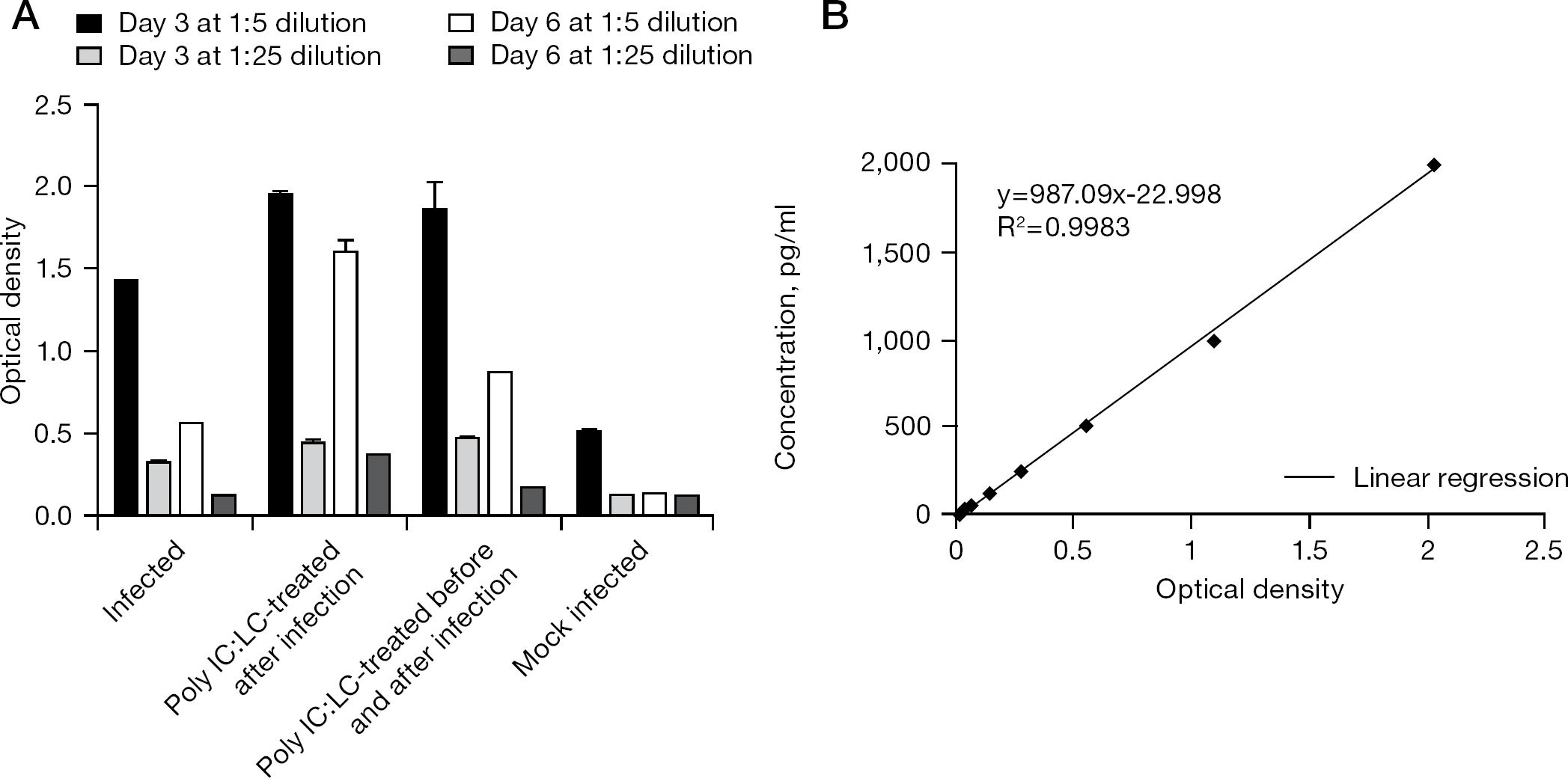
We next examined whether SARS-CoV can induce IP-10 expression in Calu-3 cells. The human IP-10 expression was detected after SARS-CoV infection, starting from day 2 (Figure 2A). IP-10 expression rate was approximately threefold less in the uninfected cells than in infected cells on day 2 and sevenfold less than in infected cells on day 4. Moreover, the effect of IFN alfacon-1 on human IP-10 expression from the culture fluids of Calu-3 cells was further tested. Interestingly, IP-10 expression was shown to be significantly increased when SARS-CoV-infected Calu-3 cells were treated with IFN alfacon-1 (Figure 2A). IP-10 expression rate in the presence of IFN alfacon-1 was increased eightfold compared with controls 5 days after plating and remained at a high level until day 7. To our knowledge, this is the first time that the IP-10 expression has been clearly demonstrated in Calu-3 cells after SARS-CoV infection. The standard curve was used to calculate pg/ml from the optical density using regression (Figure 2B).
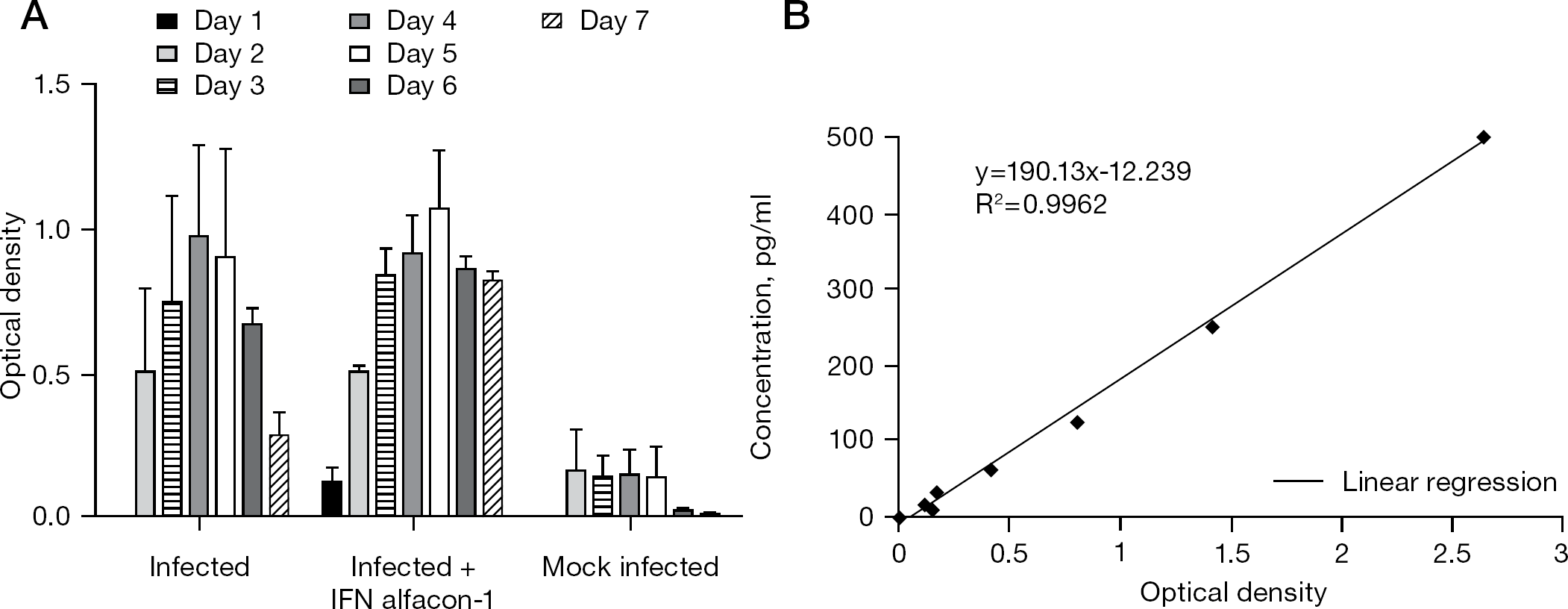
Expression of human IP-10
Replication and clinical response to mouse-adapted SARS-CoV infection
To evaluate the prophylactic potential of poly IC:LC against SARS-CoV infection, the establishment of an infection model was pivotal to permit efficacy determination of the antiviral activity. We developed a SARS-CoV-infected BALB/c mice model where death and lung pathogenesis occur as in human disease. BALB/c mice were inoculated intranasally with mouse-adapted SARS-CoV and efficient viral replication was observed from day 3–6 in the lungs (Table 1) [18]. Virus titres often exceeded 106/ml at peak replication during day 3–4. The total amount of virus present in the lungs was much greater than the inoculum, indicating that SARS-CoV replicates efficiently in the lungs of BALB/c mice. The infected mice died between 4–8 days, with 90–100% mortality rate achieved by day 8, but survivors gained back the weight by day 14. The lungs were severely inflamed and exhibited extreme lung consolidation. Weight loss was excessive; the mice that died lost >30% of their total weight by day 5 after virus exposure.
Effect of intranasal treatment with poly IC:LC on SARS-CoV replication in female BALB/c mice
Only one mouse in this group was still alive on day 6 after SARS-CoV infection. CCID50, 50% cell culture infectious dose; ND, not done; poly IC:LC, polyriboinosinic-polyribocytidylic acid stabilized with poly-L-lysine and carboxymethyl cellulose; PSS, physiologically sterile saline; SARS-COV, severe acute respiratory syndrome coronavirus.
A number of compounds were tested for efficacy in the BALB/c mice. Infected or uninfected poly IC:LC-treated mice significantly lost weight during the experiment when measured at day 0 before infection, suggesting that the dose used was near the threshold of tolerance in the mice. In the efficacy study, different groups of BALB/c mice were pre-treated or post-treated intranasally with poly IC:LC at a dose of 5 mg/kg (Table 1). At 24 h pre-treatment and 8 h post-treatment, treatment with poly IC:LC at 5 mg/kg by intranasal route were effective in protecting mice against a lethal infection with mouse-adapted SARS-CoV and reduced the viral lung SARS-CoV titres. The mice receiving the poly IC:LC treatments were also significantly protected against weight loss due to the infection. Poly IC:LC was effective in reducing virus lung scores, yet the compound did not reduce virus lung titres by day 3. Meanwhile, the survival rates of both groups were monitored daily and determined at day 14 after infection (Figure 3). Poly IC:LC significantly protected mice against death. Our data suggest that the poly IC:LC treatment of SARS infection in mice leads to a substantial therapeutic effect that protects mice against death and weight loss that results from the infection.
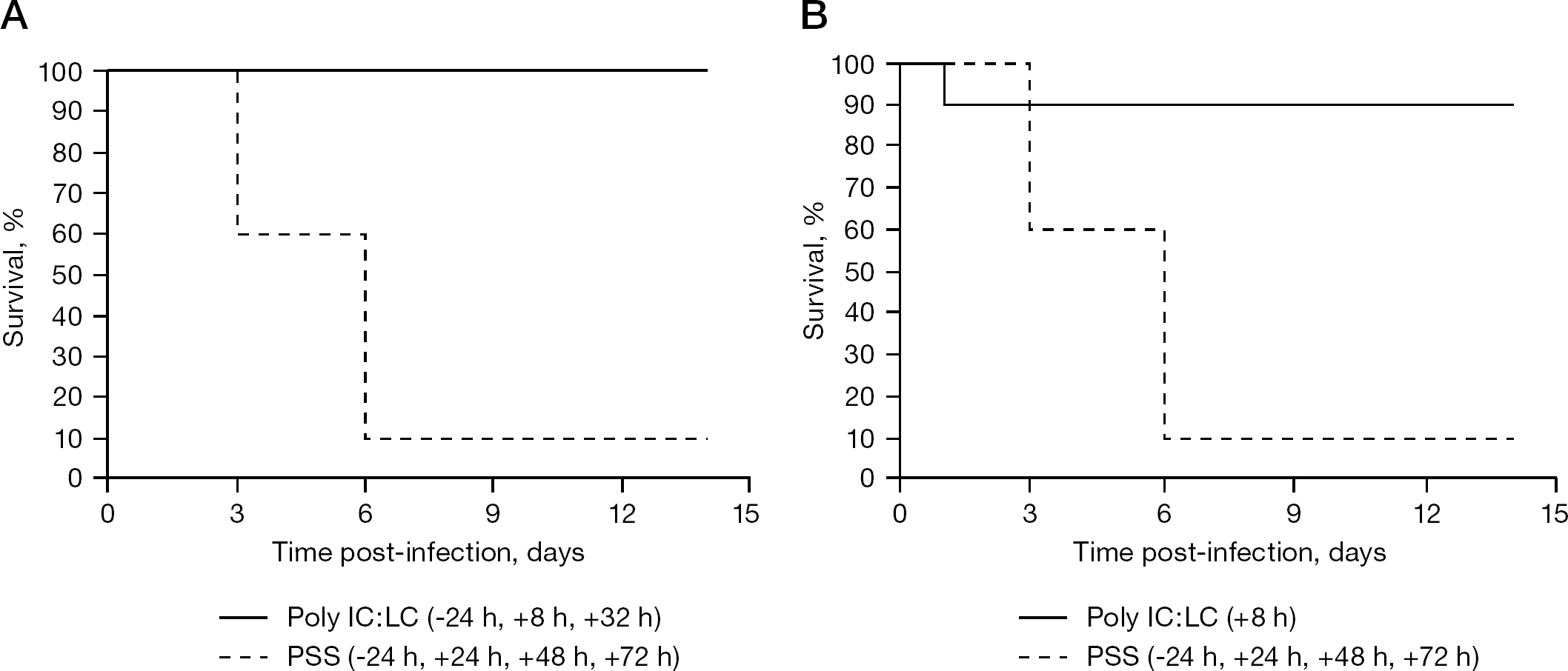
Effect of intranasal treatment with poly IC:LC on SARS-CoV replication in lethal SARS-CoV-infected BALB/c mice
Mouse-adapted SARS-CoV can induce mouse IP-10 expression in BALB/c mice
Because IP-10 seems to be coordinated with a protective response in cells, we evaluated the efficacy of antivirals directed against SARS-CoV infection in BALB/c mice. The mouse IP-10 expression was detected in the lung tissue homogenates of mouse-adapted SARS-CoV-infected BALB/c mice at day 3 and 6 (Figure 4). Moreover, the effect of poly IC:LC on mouse IP-10 expression was further tested. Significant high levels of mouse IP-10 expression in BALB/c mice was also detected when SARS-CoV-infected mice were treated with the IFN inducer, poly IC:LC.
Expression of mouse IP-10
Discussion
IFNs have been considered as the first line of defence against viral infections. One IFN that has been developed for treating a variety of virus diseases is IFN alfacon-1 [19–24]. Cinatl et al. first reported that IFN-α effectively inhibited SARS-CoV replication [25]. Paragas et al. reported that the IFN alfacon-1 had anti-SARS-CoV activity only before infection in a cell-based model in vitro [26]. Loutfy et al. demonstrated that IFN alfacon-1 seemed to ameliorate SARS-CoV infection in humans [27]. In the previous studies done in our laboratory, IFN alfacon-1 was shown to be more active against SARS-CoV when tested in human lung epithelial Calu-3 cells than in African green monkey kidney epithelial cells on day 3 post-infection [15]. In the current study, we noticed a potent, prolonged inhibition of SARS-CoV replication in Calu-3 cells with IFN alfacon-1 during the first 7 days of infection. In addition, the induced antiviral response was prolonged at near 100% inhibition. Our data from the studies in Calu-3 cells suggests that the IFN response in humans to SARS-CoV will be important in ameliorating disease and that augmenting the IFN induction response might be therapeutically beneficial.
IFN alfacon-1 is a novel, synthetic recombinant type-1 IFN-α developed by comparing the amino acid sequences of several natural IFN-α subtypes and assigning the most frequently observed amino acid in each corresponding position to generate a consensus molecule. The 166 amino acid sequence of IFN alfacon-1 was derived by scanning the sequences of several natural IFN-α subtypes and assigning the most frequently observed amino acid in each corresponding position [28]. Four additional amino acid changes were made to facilitate the molecular construction and a corresponding synthetic DNA sequence was prepared using chemical synthesis methodology. IFN alfacon-1 differs from IFN-α2b at 20/166 amino acids (88% homology), and comparison with IFN-β shows identity at over 30% of the amino acid positions. IFN alfacon-1 is produced in Escherichia coli cells that have been genetically altered by insertion of a synthetically constructed sequence that codes for IFN alfacon-1. Prior to final purification, IFN alfacon-1 is allowed to oxidize to its native state, and its final purity is achieved by sequential passage over a series of chromatography columns. This protein has a molecular weight of 19,343 Da.
The activated bronchial epithelium is an important source of IP-10, which might, in pulmonary diseases, play an important role in the recruitment of activated T-cells [29]. T-lymphocytes are involved in host defence and mucosal immunity, particularly in the respiratory tract. Activated T-lymphocytes are recruited into the lungs, where recruitment of T-lymphocytes into the sites of airway inflammation is a process involving adherence to and migration across the pulmonary endothelium, and moving into and through the airway epithelium. It has been reported previously that SARS-CoV can infect the human intestinal epithelial (Caco-2) cells to induce IP-10 expression [12,30]. In the present study, we demonstrate that human bronchial epithelial Calu-3 cells can be induced to express IP-10 after SARS-CoV infection. As IP-10, MIG and I-TAC share a common receptor, CXCR3, which is specific for these three chemokines [11], it needs to be further determined whether Calu-3 cells express MIG and I-TAC after SARS-CoV infection.
Jiang et al. [10] reported that IP-10 was augmented markedly in the blood of patients with SARS at an early stage and remained at a high level until convalescence. Moreover, IP-10 was highly detected in the lung and lymphoid tissues [10]. Induction of IP-10 is a crucial event in the initiation of immune-mediated acute lung injury and lymphocyte apoptosis during the development of SARS [10]. De Lang et al. analysed early host responses to SARS-CoV infection in the lungs of adolescent cynomolgus macaques and showed lung pathology similar to that observed in human adults with SARS [31]. In our animal study, IP-10 was also detected in the infected mouse lung after SARS infection. This result is supported by a report that IP-10 expression induced by viral infection caused a rapid recruitment of activated T-lymphocytes in the lung tissue [32]. Superinfection after immune injury and acute respiratory distress as well as lung failure are the main causes of death. The rapid increase of IL-6, IL-8 and MCP-1 is a sign of superinfection, indicating a high risk of death [10].
Poly IC:LC, an active, potent immunomodulator, was effective when therapy was initiated 24 h before infection or as late as 8 h after virus inoculation, at a time when clinical signs of SARS were being manifested in the BALB/c mice. Poly IC:LC was shown to have a potential role to play against yellow fever [33], simian hemorrhagic fever [34], Japanese encephalitis [35], rabies [36] and Rift Valley fever [37]. The antiviral activity of poly IC:LC is believed to be mediated by its ability to augment the production of IFN-α, -β and -γ [38]. Poly IC:LC also activates natural killer cells and macrophages. The antiviral efficacy of poly IC:LC was evaluated against avian influenza virus, and poly IC:LC appeared to provide effective and broad-spectrum prophylaxis against avian influenza virus [39].
In this report, IP-10 expression was shown to be significantly increased by IFN alfacon-1 in infected Calu-3 cells and with poly IC:LC in SARS-CoV-infected BALB/c mice. Taylor et al. reported that the chemokine gene for IP-10, has been induced by IFN-α in peripheral blood mononuclear cells, but were not induced by IFN alfacon-1 in human pulmonary adenocarcinoma epithelial A549 cells [40]. Sanda et al. induced IP-10 by IFN alfacon-1 and IFN-γ1b in A549 cells [41]. Sauty et al. demonstrated that human bronchial epithelial A549 cells highly express IP-10 in response to IFN-γ stimulation [11]. Yen et al. showed that the A549 and THP-1 cell lines were susceptible to SARS-CoV. A549 cells produced MCP-1 and IL-8 after interaction with SARS-CoV. Moreover, SARS-CoV induced human monocytic THP-1 cells to express CCL2/MCP-1, CXCL8/IL-8, CCL3/MIP-1α, CXCL10/IP-10, CCL4/MIP-1β and CCL5/RANTES [13]. Except for Calu-3 cells, no human respiratory epithelial cell lines have been shown as yet to support SARS-CoV infection. The human lung Calu-3 cell line provides a useful model for evaluating efficacy and mechanisms of anti-SARS compounds. As CXCR3 is a receptor for IP-10, our findings might be beneficial in the development of new anti-SARS drugs designed specifically to target CXCR3 or IP-10.
Footnotes
Acknowledgements
We thank Meeyeon Ahn for kindly providing technical assistance, and Anna Marie H Vaughan for excellent secretarial help. This work was supported by contracts NO1-A1–30048 and NO1-AI-15435 from the Virology Branch, National Institute of Allergic and Infectious Diseases, National Institutes of Health, Bethseda, MD, USA.
The authors declare no competing interests.