
Abstract
The central branches of the C7 and C8 dorsal roots were avulsed close to their entry point into the spinal cord in adult rats. The forepaw responses to heat and cold stimuli were tested at 1, 2, and 3 weeks after injury. Over this period, the paws were sensitive to both stimuli at 1-2 weeks and returned toward normal at 3 weeks. Immunohistology showed no evidence of axonal regeneration into the spinal cord in a control group of rats with avulsion only, implying that adjacent dorsal roots and their corresponding dermatomes were involved in the recovery. In a further group of rats, a mixture of bulbar olfactory ensheathing cells and olfactory nerve fibroblasts were transplanted into the gap between the avulsed roots and the spinal cord at the time of avulsion. These rats showed no evidence of either loss of sensation or exaggerated responses to stimuli at any of the time points from 1 to 3 weeks. Immunohistology showed that the transplanted cells formed a complete bridge, and the central branches of the dorsal root fibers had regenerated into the dorsal horn of the spinal cord. These regenerating axons, including Tuj1 and CGRP immunoreactive fibers, were ensheathed by the olfactory ensheathing cells. This confirms our previous demonstration of central regeneration by these transplants and suggests that such transplants may provide a useful means to prevent the development of abnormal sensations such as allodynia after spinal root lesions.
Introduction
Avulsion of spinal roots is a clinically relevant injury that often results from high-impact trauma such as road accidents. The sensory changes in response to peripheral stimuli may involve anesthesia or altered sensation such as allodynia with variable possibility of sensory recovery over time1-3. Pain is an early defining characteristic of brachial plexus injuries, affecting more than three in four patients, with many describing their pain state as distressing and unbearable4-6. The severity of this pain correlates with the number of roots avulsed during the trauma and is often accompanied by abnormally perceived mechanical or thermal sensations in the affected limb 7 . For example, an exaggerated response to cooling sensations on the injured area, cold intolerance, has been reported after brachial plexus injury, but the precise time course and pathophysiology are still not fully understood7,8. In this study, we present data on the generation of accentuated forepaw responses to heat and cold stimuli during recovery from selective unilateral avulsion of C7 and C8 dorsal roots in the rat.
Transplantation of cultured olfactory ensheathing cells (OECs) has been shown to induce axon regeneration, remyelination, and restoration of function after experimental spinal cord injury9-15. For dorsal root injuries in particular, we developed a matrix method of transplantation, which involves scraping cells from a mixed culture of OECs and olfactory nerve fibroblasts (ONFs) grown within their own endogenous matrix 16 . Subsequent work is beginning to reveal the importance of the role played by ONFs in laying down a path in front of OECs, and thus, we refer to the transplants as “OEC/ONFs”17,18. Recently, an autologous transplant of OEC/ONFs was shown to give partial recovery of superficial and deep sensation along with an increase in muscle mass in the lower limb after a thoracic level injury in a patient 19 .
In a previous rat study, we provided evidence that transplantation of a mixture of OECs and ONFs into a unilateral C6–T1 four root transection induced regeneration of cut dorsal root axons into the spinal cord 20 . This was determined by histology and electrophysiological transmission and reinstated sufficient proprioceptive function to restore forepaw control in a climbing test. Since we found that these animals did not show any sensitivity to heat/cold stimuli, in this current study we opted to avulse (i.e., pull) the dorsal roots from the spinal cord rather than transect, to better mimic what occurs clinically after brachial plexus injury.
Although a number of our earlier studies showed regenerating axons within the spinal cord, we have not yet assessed the impact of these fibers on pain outcomes11,16,20,21. For example, it is not known what effect regeneration of sensory afferents into the damaged dorsal horn would have on neuropathic pain in a brachial plexus injury. Aberrant sprouting of sensory fibers that terminate in the “wrong” lamina of the superficial dorsal horn has been implicated in chronic pain states22,23. Neurotrophic factors are reported to facilitate this sprouting, which is relevant to our study as OECs are known to release factors such as nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), or glial cell line-derived neurotrophic factor (GDNF)23-25.
In this present study, we have examined the effect of OEC/ONF transplants on the recovery of forepaw responses to thermal stimulation following avulsion of dorsal roots of cervical spinal nerve 7 (C7) and C8.
Materials and Methods
Cell Culture
A detailed description of the cell culture protocol is given in our previous study 26 . Briefly, under aseptic conditions the nerve and glomerular layers of the olfactory bulbs from adult male Sprague–Dawley rats were dissected out, dissociated using 0.1% trypsin (0.1%; Worthington Biochemical Corporation, Lakewood, NJ, USA), and cultured in a medium consisting of Dulbecco's modified Eagle's medium (DMEM)/F-12 and 10% fetal bovine serum supplemented with penicillin–streptomycin (100 U, 100 μg/ml; all from Invitrogen, Paisley, UK). The cultures were monitored until confluent (reaching approximately 1.5 × 106 cells/dish at 14-16 days), at which time the OECs were accompanied by roughly equal numbers of ONFs and embedded in an endogenous matrix. Two days prior to transplantation, the cells were infected with a lentiviral vector expressing green fluorescent protein (GFP) 27 . One hour before surgery, the cells were viewed using an epifluorescence microscope to ensure that GFP labeling was visible and that the dishes contained a uniform distribution of OEC/ONFs in roughly equal proportions.
Surgery
Under isoflurane (Abbott Laboratories Ltd., Berkshire, UK) anesthesia, dorsal roots C7 and C8 were exposed as previously described 20 . Briefly, in a posterior approach, a skin incision was made through the dorsal midline, the paraspinal muscles were separated, and the prominent spinous process of the second thoracic vertebra (T2) was located. Hemilaminectomies were carried out from C6 to T1, and the dura was incised with microscissors to reveal the dorsal roots.
To avulse the C8 dorsal root, thin watchmaker forceps (Dumont Number 5; Fine Science Tools, Heidelberg, Germany) were used to grasp the root close to the point of entry to the spinal cord, including all of its rootlets. The root was then rapidly pulled away from the cord before the adjacent C7 root was similarly avulsed. Complete separation from the spinal cord was confirmed before the avulsed roots were then placed back to lie at their former entry point. Dorsal roots C7 and C8 were chosen because Ramer et al. previously showed that, following their transection, there was transient cold hyperalgesia in the plantar area of forepaw 28 . Furthermore, our preliminary studies showed that avulsion of too many cervical dorsal roots increased the chance of autotomy or led to the inversion of the forepaw such that stimulation by heat/cold was not possible.
At the time of avulsion, in 16 out of 34 rats, matrix containing GFP-labeled OEC/ONFs was scraped from the dish to give around seven 0.5- to 1-mm2 portions, each containing a roughly equal proportion of OEC/ONF (see the Results section for an illustration of GFP-labeled cells prior to transplantation, the cell scraping method, and portions of OEC/ONF pellets). These cells produce their own matrix, which makes them sticky, and they were placed between the avulsed ends of the C7 and C8 dorsal roots to bridge the gap with their former point of entry to the spinal cord and were retained in position with fibrin glue (Tisseel Kit; Baxter Healthcare Ltd., Thetford, UK). For the control group of 18 rats, the avulsed roots were similarly repositioned close to their original entry points into the spinal cord but without the additional OEC/ONFs. The muscles were repaired in layers, the skin sutured, and the rats were given postoperative pain relief [0.05 mg/kg buprenorphine (Vetergesic; Reckitt Benckiser Healthcare, Hull, UK), subcutaneously (SC), daily for 3 days]. Control and transplanted rats were mixed together randomly in the same cage to make blinding easier.
The rats showed no adverse effects following surgery and took part in behavior tests 1 week after injury. One nontransplanted rat with bleeding under the nails of the ipsilateral forepaw at 12 days postsurgery was terminated. In terms of housing conditions, room temperature was maintained between 20°C and 22°C under standard lighting conditions (12:12 light–dark cycle) with food and water available ad libitum. Animals were handled according to the UK Home Office regulations for the care and use of laboratory animals, the UK Animals (Scientific Procedures) Act 1986 with the ethical approval of the University College London (UCL) Institute of Neurology.
Behavior
Thirty-four male Sprague-Dawley rats weighing 150-175 g were acclimated by presentation to the testing cage and environment for 1 week. The following week, all rats were handled each day for around 5 min each. Toward the end of the second week, rats were placed singly in a Perspex (17 cm × 23 cm × 14 cm) behavior testing box for 15 min daily. During behavior tests, rats were randomly assigned to these boxes to mix up controls and transplanted animals, making blinding assessment easier and allowing for the fact that rats are empathic animals and could be influenced by their neighbor's responses.
Hargreaves Test
An automated thermal plantar test (Ugo Basile, Varese, Italy) was used to assess sensitivity of the forepaw plantar surface (dermatomes C7 and C8) 29 to increasing thermal stimulation. Animals were tested at 2:00 to 4:00 in the afternoon. The time taken until forepaw withdrawal was detected automatically, and each trial was separated by at least 5 min. Alternating ipsilateral and contralateral (i.e., uninjured) forepaws were tested three times each, and the reaction time was averaged. Trials were performed pre-operatively and at 7, 14, and 21 days after injury by a blinded assessor.
Acetone Test
Acetone tests took place in the morning (10:00 a.m. to noon) and were carried out under baseline conditions and at 7, 10, 14, and 21 days after injury. To assess forepaw sensitivity to cold stimuli, rats were individually placed in a Perspex box on a mesh grid and allowed to acclimate for 15 min. From underneath and, hence, out of vision of the rat, 10 μl of acetone was applied to the center of the plantar surface of the forepaw (dermatomes C7 and C8). Ipsilateral and contralateral (i.e., uninjured) forepaws were tested three times each with at least 5 min between trials and video recorded (Fig. 1).

Examples of paw raising (A) and attention to paw (B, C) after acetone stimulation.
Video records were assessed blind using a software (Media Player Classic; Microsoft, Los Angeles, CA, USA) to enable behavior for 1 min following each acetone application to be studied in slowed time, scoring (a) the total length of time in which the stimulated forepaw was raised and (b) how often the rat attended to the stimulated forepaw (e.g., by shaking, licking, or examining) (see Fig. 1).
Tissue Processing and Immunohistochemistry
The rats were terminated under deep carbon dioxide anesthesia and transcardially perfused with 50 ml of 0.1 M phosphate-buffered solution followed by 400 ml of phosphate-buffered 4% paraformaldehyde (PFA). The vertebral columns were dissected out from the craniocervical junction to the upper thoracic level and left to harden in the same fixative for 48 h. The spinal cord and associated roots were carefully dissected out of the bony skeleton under a dissecting microscope to preserve the continuity across the avulsed dorsal roots and transplants to the spinal cord. The dissected tissues were placed into 10%-20% sucrose solution until sunk, frozen with cryospray (Cell Path, Newtown, UK), and 16-μm sections were cut on a cryostat (Leica CM3050; Leica, Heidelberg, Germany): n=20 in the coronal plane from levels C7 and C8, and n=8 in the horizontal plane from levels C6–T1. For immunostaining, the sections were incubated in 2% milk (Oxoid Ltd., Basingstoke, Hampshire, UK) containing primary antibodies outlined in Table 1 and incubated overnight at 4°C. Secondary antibodies are also detailed in Table 1 (all Invitrogen) and were incubated for 2 h in the dark at room temperature. Some sections were counterstained with either To-Pro 3 (1:1,000; Life Technologies, Renfrew, UK) or Sytox (1:1,000; Thermo Fisher Scientific, Waltham, MA, USA). Control sections were stained with primary antibodies, and secondary antibodies were omitted.
Antibodies Used in Immunohistochemistry

A separate group of six rats received the same C7 and C8 dorsal root avulsion with OEC/ONF transplantation as described above, but did not undergo any behavior testing and were terminated 1 week after injury for histological assessment.
Statistical Analysis
Results were expressed as means±standard error of the mean (SEM), with statistical comparisons between groups made using a one-way analysis of variance (ANOVA), to determine F-ratio significance. Post hoc analysis was performed with Bonferroni multiple comparisons and SPSS Statistics 22.0 (IBM, Armonk, NY, USA) software was used. Details of animal numbers at all time points are given below each graph.
Results
Functional Tests
Hargreaves Test
The normal response to the infrared beam is to raise the forepaw at a mean of 12 s (±0.4) after applying the stimulus. In the rats with avulsed roots, the response is significantly brisker, reaching a peak mean of 9.2 s (±0.4) at 7 days after avulsion and returning to normal by 21 days (Fig. 2). In the rats with transplanted OEC/ONFs, this abnormally brisk response was prevented, and the animals took around 2-3 s longer to withdraw than controls.
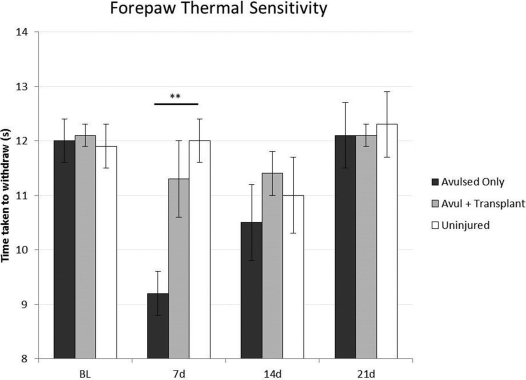
Hargreaves test. Rats that received cervical spinal nerve 7 (C7) and C8 dorsal root avulsion (dark bar) were transiently hypersensitive to thermal stimuli 1 week after injury. Rats that had olfactory ensheathing cell/olfactory nerve fibroblast (OEC/ONF) transplants at the time of avulsion injury (gray bar) showed thermal sensitivity similar to a normal, uninjured forepaw (blank bar). Error bars: mean±standard error of the mean (SEM) (**p < 0.01, post hoc Bonferroni after one-way analysis of variance). “Avulsed Only” and “Uninjured” groups: n = 14 for baseline (BL) and 7 days (d), n = 13 for 14d, n = 6 for 21d. “Avul + Transplant” group: n = 12 for BL and 7d, n = 11 for 14d, n = 10 for 21d.
Acetone Test
The normal response to the acetone application is to maintain the forepaw raised for under 5 s after applying the stimulus. The rats briefly look at the forepaw and may also shake it.
In the rats with avulsed roots, the forepaw is raised for a mean of 22.2 s (±4.9) at 7 days after avulsion, returning close to the normal range by 21 days. There was an exaggerated level of attention to the forepaw, which sometimes included licking (Fig. 3).

Acetone test. Raised paw duration: Rats that had cervical spinal nerve (C7) and C8 dorsal roots avulsed (dark bar) kept their stimulated forepaw raised for longer after acetone application than both uninjured (blank bar) and rats that received a transplant at the time of injury (gray bar). Error bars: mean±standard error of the mean (SEM) (*p < 0.05, **p < 0.01, ***p < 0.001, post hoc Bonferroni after one-way analysis of variance). “Avulsed Only” and “Uninjured” groups: n = 14 for baseline (BL), 7 days postinjury (dpi), and 10 dpi; n = 13 for 14 dpi; n = 6 for 21 dpi. “Avul + Transplant” group: n = 12 for all time points.
The rats with transplanted OEC/ONFs behaved as normal; no exaggerated responses were observed.
In both avulsed and transplanted groups, the heat- and cold-induced responses of the contralateral (uninjured) paw showed no statistical deviation from those before operation (Figs. 3 and 4).

Acetone test. Number of forepaw attends: Rats that had cervical spinal nerve (C7) and C8 dorsal roots avulsed (dark bar) attended to their stimulated forepaw after acetone application more frequently than both uninjured (blank bar) and rats that received a transplant at the time of injury (gray bar). Error bars: mean±standard error of the mean (SEM) (*p < 0.05, **p < 0.01, ***p < 0.001, post hoc Bonferroni after one-way analysis of variance). “Avulsed Only” and “Uninjured” groups: n = 14 for baseline (BL), 7 days postinjury (dpi), and 10 dpi; n = 13 for 14 dpi, n = 6 for 21 dpi. “Avul + Transplant” group: n = 12 for all time points.
Immunohistological Observations
In the normal situation, the traverse of the central branches from the C7 and C8 dorsal root ganglia to the entry into the spinal cord is less than 2 mm. The mechanical force involved in avulsion of the dorsal roots causes total destruction of the substantia gelatinosa and major damage to the superficial laminae of the dorsal horn at the lesion level (“x” in Fig. 5E). The central stumps of the detached roots remained in position close to the spinal cord.

(A, B) Epifluorescence image of green fluorescent protein (GFP)-labeled olfactory ensheathing cell/olfactory nerve fibroblasts (OEC/ONFs) on a culture dish 1 h prior to transplantation (A) and the cell scraping method used to detach sticky OEC/ONF pellets from the surface of the culture dish, ready for transferring to the animal (B) (inset: example of four pellets). (C, D) Images during microsurgery showing dorsal root cervical spinal nerve (C8) (arrow) immediately prior to avulsion (C) and then a pellet of OEC/ONFs (arrow) about to be placed between the avulsed end and its former entry point in the spinal cord (D). (E, F) Confocal images of cross sections of the dorsal spinal cord between cervical levels C7 and C8 after dorsal root avulsion in a control rat (avulsion only) (E) and in a rat that received avulsion plus an OEC/ONF transplant (F). Normal pattern of calcitonin gene-related peptide (CGRP) staining on the right side of the superficial dorsal horn in (E) and (F); disrupted pattern of CGRP+ staining on the left side of the dorsal horn in (E) and (F); transplanted cells identified with GFP labeling formed a continuous mass between the avulsed root and the spinal cord in (F). Arrow in (C), C8 dorsal root; arrow in (D), OEC/ONF pellet on forceps; av, avulsed dorsal root; cc, central canal; dc, dorsal columns; N, intact dorsal root; x, lesion of dorsal horn; R, reapposed dorsal root. Double immunostaining of CGRP (red) and glial fibrillary acidic protein (GFAP) (green) in (E), OEC/ONF (green GFP), CGRP (red), and 4′,6-diamidino-2-phenylindole (DAPI) (blue) in (F). Survival time: 1 week. Scale bars: 200 μm (A), 4 mm (C, D), 500 μm (E, F).
Rats with Avulsion Alone
Despite their proximity, there was no histological indication of continuity between the detached ends of the dorsal roots and the spinal cord. This was confirmed with double immunostaining of laminin/glial fibrillary acidic protein (GFAP) and, separately, with calcitonin gene-related peptide (CGRP)/GFAP (Fig. 5E).
Rats with Transplanted OEC/ONFs
The OEC/ONFs remained at the site of transplantation. They formed a densely packed mass from the detached root ends all the way to the spinal cord (Fig. 5F). The cells were generally elongated for up to 100 μm along the trajectory of the former dorsal root fibers. Peripherally, the cells migrated for around 100 μm into the dorsal root. Centrally, they invaded the lesioned area of the dorsal horn and penetrated for up to 500 μm into adjacent tissues. The areas with the transplanted cells were markedly hypervascularized. The new vessels were surrounded by GFP-labeled transplanted cells (Fig. 6I).

Immunohistology of the dorsal spinal cord at level C7–C8 in normal rats, control rats with avulsion only, and rats with avulsion and transplants. (A–C) Resting state microglia (Iba1, green) at the superficial dorsal horn in normal rats (A), proliferation of activated microglia with enlarged soma in rats with avulsion only (B) and in rats with avulsion and olfactory ensheathing cell/olfactory nerve fibroblast (OEC/ONF) transplant (C). (D–F) Isolectin (IB4; red) staining of small-diameter nociceptive afferents in lamina II of normal rats (D) in the disrupted dorsal horn of rats with avulsion only (E) or in rats with avulsion and OEC/ONF transplant (F). (G–I) Superficial dorsal horn of rats with avulsion and OEC/ONF transplant showing double immunostaining against immature (Tuj1+, green) and peptidergic nociceptive afferents [calcitonin gene-related peptide (CGRP+), red] (G), location of OEC/ONF transplants [green fluorescent protein (GFP), green] (H), and transplanted cells in the vicinity of, and intimately surrounding, new blood vessels [rat endothelial cell antibody (Reca1), red] (I). (J) Horizontal section showing transplanted cells (green, GFP) in the reapposed dorsal root (R), the junctional region (J), and the spinal cord (SC), and transplanted cells ensheathing CGRP+ axons (red, arrows). (K) Highpower micrograph of cross section showing elongated transplanted cells (GFP, green) ensheathing axons (arrows) in the dorsal root (neurofilament+ axons, red). Arrowheads, region of cells and axons turning down into the dorsal horn. Survival time: 3 weeks for (A)–(G) and (J), 1 week for (H), (I), and (K). Scale bars: 200 μm (A–G), 100 μm (H–J), 25 μm (K).
The entire area of the transplanted cells was densely infiltrated by CGRP and neurofilament-immunostained fibers. Immunostaining for Tuj1, a marker for immature neurons, colocalized with that for the small diameter, nociceptive CGRP afferents (Fig. 6G). Horizontal sections showed the fibers were ensheathed in a one-to-one fashion by the transplanted cells (Fig. 6J and K). Figure 6K shows the concentration of transplanted cells with their ensheathed axons over the point at which they change direction to turn from the root into the spinal cord.
Following injury, activated microglia with large soma were located around the cavity in both the control rats with avulsion only and in those with avulsion and transplanted cells (Fig. 6A–C). The number of IB4+ fibers was reduced within the superficial laminae after the dorsal root avulsion injury. Transplant and control groups showed a similar pattern of immunostaining for this nonpeptide nociceptive afferent, although accurate quantification was not possible because tissue sections had variable levels of damage at the superficial dorsal horn.
Discussion
Following avulsion of the central branches of the C7 and C8 dorsal roots, we demonstrate a transient period of exaggerated responses to heat and cold stimuli applied to the forepaw. In the absence of any histological indication of regeneration of the avulsed C7 and C8 roots to the spinal cord, these abnormal responses would be mediated through the intact roots of the nerves supplying the adjacent dermatomes of C6 and T1. It is known that sectioning of dorsal root(s) and the associated damage to the dorsolateral tract can lead to an enlarged area of mechanical and thermal sensitivity within the “border zone” of corresponding dermatomes 30 . There are many studies that use peripheral injuries to examine the same phenomenon at the thoracic or lumbar levels, but here we focus on the more clinically relevant upper limb31-35.
One crucial difference between our injury model and the real-life situation is that although we avulse the root from its entry point in the cord, after real trauma, the traction aspect of the injury causes the root with its ganglion to be dislodged more laterally 2 . This is important to note because a distant dorsal root ganglion (DRG) would likely offer less trophic support to dorsal horn neurons. Future studies should examine the influence of DRG proximity to the damaged spinal cord and whether OEC/ONF transplants aid in the survival or trophic support of these cells, particularly in the early stages after injury. Experiments carried out by Rodrigues-Filho et al. reveal how the location of injury heavily impacts the performance on thermal sensitivity tests 36 . They avulsed C8 and T1 peripherally in the lower trunk, close to the subclavian vessels, to cause a traction injury that resulted in long-term mechanical and cold allodynia, but interestingly no such sensitivity to heat. We chose to avulse the dorsal roots at their immediate point of entry to the spinal cord to investigate the “bridging function” of OEC/ONFs across the damaged dorsal root entry zone.
In a previous study of a four-dorsal root transection, we showed that transplanted OEC/ONFs form a bridge allowing cut peripheral axons to regenerate into the spinal cord and restore proprioceptive function 20 . Although the rats' ability to sense the position and movement of the forelimb was improved, they were nonresponsive to thermal stimulation on their injured side for the entire duration of the 8-week study (Collins A, Li Y, and Li D, unpublished observations and Cragg et al. 37 ). To generate sensitivity to heat/cold and replicate the damage that is done to the spinal cord in a real-life injury, dorsal roots C7 and C8 were avulsed rather than transected3,28. The resulting vascular and cellular destruction of the dorsal horn at the cervical level was similar to that shown by others who avulsed roots at the lumbar level38-40. Importantly, the extent and location of damage to the dorsal horn along at least two segments of the cervical spinal cord were also similar to that seen in imaging studies of patients with brachial plexus injuries41,42. We did not examine the response of the adjacent, uninjured C6 or T1 dorsal roots and their afferent input to the dorsal horn above or below the injured segments. Future axonal tracer studies could selectively label these adjacent roots and investigate their role in afferent sprouting within the dorsal horn; here we focus on transplant-mediated effects on the most damaged segments of the cervical spinal cord—levels C7 and C8.
The present study showed that, in contrast to control rats with avulsion only, rats that received OEC/ONF transplants did not show any signs of sensitivity to heat/ cold after injury and were behaviorally indistinguishable from normal rats. By the final week of the study, control rats with “avulsion injury only” had stopped showing exaggerated responses to thermal stimuli. The protracted response to acetone application has been shown in other studies involving transection of C7 and C8 dorsal roots or more commonly with injury to dorsal and/or ventral roots of the lumbosacral cord28,43,44. For example, Ramer et al. transected dorsal roots C7 and C8 and found a prolonged response to acetone at days 5 and 10 postinjury, but with only some animals showing sensitivity to radiant heat 28 . In our study, the sensitivity to cold was longer lasting and more pronounced. Most likely, this is because we avulsed dorsal roots C7 and C8, thus causing more damage to the vasculature and dorsal horn than would occur after rhizotomy surgery3,28.
Selective genetic ablation of CGRP sensory neurons has also resulted in rats attending to their paw for longer after acetone application 45 . It is apparent that our C7 and C8 avulsion injury also removes CGRP+ sensory afferents from the superficial dorsal horn, and this could account for the exaggerated response to cold. Given the natural tendency for injuries to occur at the cervical level and anecdotal accounts of “intense, unbearable pain” that may accompany a cool, breeze-like sensation on a patient's forearm, it is essential to have an animal model with which to test treatments for cold tolerance6,46,47. An absence of vocalization, coat piloerection, or porphyrin around the eye suggested that the rats were not in distress during the behavior tests or when resting in their home cages.
Although thermal sensitivity scores returned to baseline levels after 3 weeks, this still provides a suitable window to examine the early anatomical changes that occur in the dorsal horn and adjacent areas following a lifelike avulsion injury to the cervical spinal cord. In preliminary studies, we had attempted to assess forepaw mechanical sensitivity using von Frey filaments. However, recordings were inconsistent, as it was apparent that the injured rats distributed their weight differently between their forepaws after unilateral C7–C8 dorsal root avulsion injury (Collins A, Li Y, and Li D, unpublished data). For this reason and to focus on the heat/cold responses during the limited postinjury period without overtesting the animals, we did not assess mechanical sensitivity in this current study.
Histological analysis indicates that neurofilament+, CGRP+, and Tuj1+ fibers from the avulsed C7 and C8 dorsal root have regenerated into the spinal cord in the group of rats that received transplantation of OEC/ONFs. We have previously used biotinylated dextran amineaxonal tracer and electrophysiological approaches to show that OEC/ONF transplants into injured spinal cord or spinal roots led to axonal regeneration and the reinstatement of sensory afferents into the dorsal column11,16,20,21,48. In the present study, we observed regenerating CGRP+ and immature (Tuj1+) neurons adjacent to the cavity area in the dorsal horn of rats that had received OEC/ONF transplants. Short, small-diameter neurofilament+ and CGRP+ fibers were oriented toward the dorsal horn with many ensheathed by the OECs. However, we did not see this pattern of CGRP+ and Tuj1+ staining in the control group of rats with avulsion only.
Ensheathment by OECs could protect the newly sprouted axons and, perhaps with enhanced trophic support, help to prevent the ectopic firing associated with pain states25,49,50. In contrast, nonpeptide nociceptive afferents such as those revealed by isolectin–IB4 binding showed an obvious reduction after injury, but there was no quantifiable difference between control and transplanted animals. Although studies from other groups have described nociceptive afferent sprouting within the dorsal horn after experimental spinal cord injury22,23,51,52, they do not show the ensheathment of regenerative sprouts, which we see in our study. Given the extent of mechanical destruction of the superficial dorsal horn following avulsion, it was not possible to delineate the laminar terminations of regenerating fibers and assess their potential contribution to functional responses. The extent and variable nature of destruction of the dorsal horn tissue of C7–C8 segments made it hard to accurately quantify immunoreactivity in this area; future work will focus on such changes at the intact adjacent levels of C6 and T1.
As early as 7 days after injury, there was a difference in thermal sensitivity between control and transplanted rats. The briskness of this transplant-induced functional benefit suggests that axonal regeneration may not be the only or principal explanation behind the prevention of this heightened response to thermal stimuli. We know from our previous work that there can be some degree of transplant-induced functional return even in the absence of axonal regeneration 53 , suggesting that other factors may dampen dorsal horn excitability or limit the secondary damage following avulsion 54 . For example, given that one third of neurons in laminae I-III are γ-aminobutyric acid (GABA)ergic interneurons, avulsion is likely to reduce the local inhibitory tone, which normally prevents pain signaling from innocuous stimuli52,55. We did not measure levels of glutamic acid decarboxylase (GAD) 67 or GABA receptor expression, and so we cannot be sure whether different levels of dorsal horn excitability were involved in the divergent functional responses of control and transplanted rats.
The contribution of glial cells to behavioral hypersensitivity after avulsion injuries has been demonstrated previously38,56-58. Chew's group found that riluzole prevented thermal sensitivity after lumbar level avulsion by reducing microglial activation within the dorsal horn 38 . Although in our study we could see no discernible difference between control and transplanted rats in terms of the morphology or abundance of microglia (Iba1+) within the dorsal horn, we only investigated at 3 weeks postinjury and not at any earlier time points. There is a close interplay between degenerating nociceptors and the early immune response in terms of the local coordination required to reestablish homeostasis 59 . Indeed, several studies have shown a temporal correlation between the invasion of blood-borne macrophages and the generation of allodynia60,61. Our previous electron microscopy study demonstrated the active phagocytic role played by OECs in their removal of degenerating olfactory axons 18 . Given this capacity to remove axon debris as well as provide trophic support and closely interact with new blood vessels, they may assume aspects of immune system function in the crucial, early stages after injury62-64. The precise mechanism by which OEC/ONFs prevent abnormal, heightened responses to heat/cold in our current study is not yet fully understood.
We have found that acute transplants of OEC/ONFs prevented any detectable sensitivity to increasing heat or innocuous cold stimuli on the forelimb after unilateral avulsion of dorsal roots C7 and C8 in rats. Regenerative sprouts of CGRP+ fibers in areas outside of lamina I–II did not appear to worsen any pain outcomes following avulsion injury. On the contrary, rats that received transplants showed a complete absence of the abnormal functional responses seen for 1-2 weeks in controls. This finding has important implications since such transplants in a clinical setting would be presumed unlikely to worsen any existing pain symptoms. The OEC/ONF-induced ensheathment of small-diameter neurofilament+ and CGRP+ fibers within the dorsal horn, along with early inflammatory responses such as clearing of debris or trophic nourishment of new blood vessels could all partly act to prevent generation of forepaw sensitivity.
Patients with brachial plexus injuries often describe paroxysmal attacks of spontaneous shooting pain, which are obviously difficult if not impossible to directly model in rodents. However, the abnormal responses to heat/cold stimuli on the rodent forepaw that we have observed may serve as a surrogate for these clinical symptoms. For instance, although exaggerated responses to innocuous cooling sensations are not reported by all patients, our use of the acetone test gives a relatively objective picture of exaggerated forelimb responses in the early stages following avulsion injury47,65. As such, this experimental design may provide a useful basis for investigating potential strategies to ameliorate the inappropriate sensations, which can have a profound negative impact on the quality of life following spinal root or peripheral injuries.
Footnotes
Acknowledgments
This project was supported by the Medical Research Council (MR/J015369/1), the UK Stem Cell Foundation, and the Nicholls Spinal Injury Foundation. The authors are grateful to Dr. Xuenong Bo for his help with the lentiviral vector and Miss Sara Bowie for her excellent technical support. The authors declare no conflicts of interest.