
Abstract
Despite the increasing importance of minipigs in biomedical research, there has been relatively little research concerning minipig-derived adult stem cells as a promising research tool that could be used to develop stem cell-based therapies. We first generated immortalized neural stem cells (iNSCs) from primary minipig olfactory bulb cells (pmpOBCs) and defined the characteristics of the cell line. Primary neural cells were prepared from minipig neonate olfactory bulbs and immortalized by infection with retrovirus carrying the v-myc gene. The minipig iNSCs (mpiNSCs) had normal karyotypes and expressed NSC-specific markers, including nestin, vimentin, Musashi1, and SOX2, suggesting a similarity to human NSCs. On the basis of the global gene expression profiles from the microarray analysis, neurogenesis-associated transcript levels were predominantly altered in mpiNSCs compared with pmpOBCs. These findings increase our understanding of minipig stem cells and contribute to the utility of mpiNSCs as resources for immortalized stem cell experiments.
Introduction
Neural stem cells (NSCs) are multipotent cells that originate from the nervous system and are capable of self-renewal 1 . In particular, NSCs are promising for stem cell therapies for a variety of human neurological disorders, such as spinal cord injury, stroke, Alzheimer's disease (AD), Parkinson's disease (PD), Huntington's disease (HD), and amyotrophic lateral sclerosis (ALS), as well as for basic research in areas including neural development 2 . Although the importance of NSCs in biomedical research is increasing, their limited proliferation capacity in vitro is a major obstacle to expanding their usability. Therefore, several researchers have attempted to generate immortalized NSCs (iNSCs) with high proliferation potentials and without the loss of neural differentiation3–5. However, research is necessary on iNSCs derived from a variety of species to improve their feasibility as resources for basic research and stem cell therapy.
Pigs have been used widely in preclinical studies because of their high anatomical, physiological, genetic, and biochemical similarities to humans 6 . The most common experimental data using mammalian models are produced from rodents and dogs 7 , and preclinical data from rodents are often necessary to support the use of larger animals in clinical applications 8 . However, minipig models are becoming increasingly promising based on their close phylogenic distance to humans and their large litter size, as well as the relatively few ethical issues related to this species 6 ; thus, these models could increase the ability to produce more reliable data for the discovery of new drugs 9 . Many research groups have tried to isolate resources from a variety of minipig tissues8,10 as well as to generate disease models and bio-organ donors via transgenesis methods 11 . Despite the increasing importance of minipigs as a larger higher animal model for human diseases, there has been little or no research concerning the derivation and use of minipig iNSCs (mpiNSCs).
In this study, we generated mpiNSCs via stable transfection of primary minipig olfactory bulb cells (pmpOBCs) with a retroviral vector encoding the v-myc gene. The mpiNSCs successfully expressed NSC-specific markers, including nestin, vimentin, and (sex-determining region Y)-box 2 (SOX2). Moreover, the microarray analysis demonstrated that mpiNSCs highly expressed neurogenesis markers, particularly genes related to the Wnt signaling pathway. The results support that mpiNSCs can be used as efficient models in studies on neural differentiation and stem cell therapy.
Materials and Methods
Culture of pmpOBCs
Minipig cells (Micro-pig®; Medi Kinetics, Gyeonggido, Republic of Korea) were isolated from 7-day-old neonatal olfactory bulb tissue, dissociated using papain (Sigma-Aldrich, St. Louis, MO, USA), and plated as primary cultures. Use of minipig cells was approved by the Chung-Ang University Ethics Committee (Certificate #09-0041). The pmpOBCs were grown in Dulbecco's modified Eagle's medium (DMEM) with high glucose (Welgene, Kyungsangbuk-do, Republic of Korea) and F12 medium (1:1; Welgene), supplemented with 10% fetal bovine serum (FBS; GenDEPOT, Katy, TX, USA), 20 mg/ml gentamicin, 10 ng/ml basic fibroblast growth factor (bFGF; Sigma-Aldrich), and epidermal growth factor (EGF; Gibco, Carlsbad, CA, USA). The cells were incubated at 37°C with 5% CO2 in a humidified incubator.
Immortalization of Minipig Cells Using v-myc Gene
pLNCX2.v-myc plasmid containing full-length avian v-myc cDNA derived from Gallus gallus was used in this study. The v-myc gene was inserted into a pLNCX2 retroviral vector using BglII and HindIII restriction. Retroviral vectors were transfected into a packaging cell line to produce v-myc retrovirus using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). These viruses were transduced into minipig cells and selected for by supplementing the medium with 1,000 μg/ml neomycin for 4 weeks. Neomycin-resistant minipig colonies were isolated and used for further study.
Karyotypic Analysis
The cell lines were plated at a density of 0.5 × 106 cells in 60-mm culture dishes (BD Falcon, Tewksbury, MA, USA) coated with 1% gelatin. After 24 h, the growth medium was replaced with medium containing 0.1 μg/ml colcemid (KaryoMAXTM Colcemid; Gibco) and incubated for 4 h. After dissociation, cells were incubated with 0.075 M KCl for 15 min in a 37°C water bath. Then three to five drops of freshly prepared fixative (3:1 methanol/glacial acetic acid) were added to stop the reaction, and cells were fixed in fixative at 4°C. One drop of cell suspension was placed on glass slides, and the slides were dried at 60°C for a minimum of 18 h and incubated in a trypsin solution for 30 s. Slides were stained with KaryoMAXTM Giemsa stain (Gibco) for 5 min. After drying, the slides were mounted and observed under a microscope.
RT-PCR Analysis
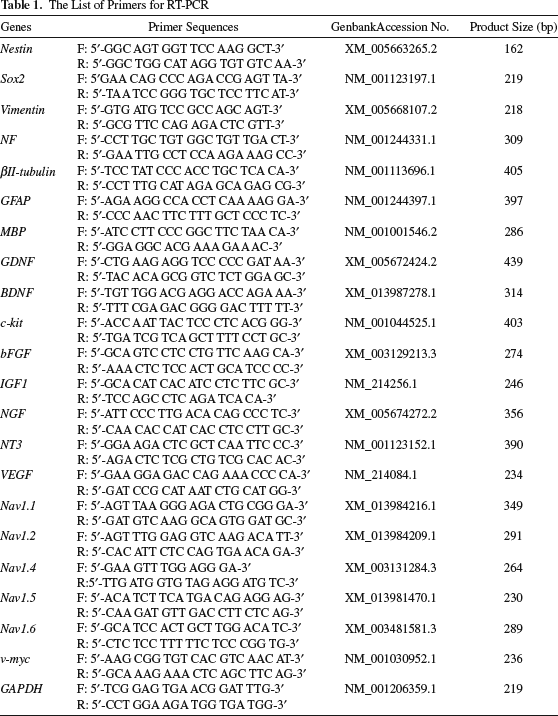
Total RNA was isolated from pmpOBCs and minipig NSCs using an miRNA Isolation Kit (Qiagen, Hilden, Germany). First, 1 μg of total RNA was reverse transcribed into first-strand cDNA using oligo-dT primer. Reverse transcription was performed with TOPscriptTM RT DryMIX (Enzynomics, Daejeon, Republic of Korea) for 1 h at 42°C, inactivated for 5 min at 95°C, and cooled to 4°C. The cDNA was amplified using 30 polymerase chain reaction (PCR) cycles, and real-time (RT)-PCR products were separated in 1.5% agarose gels and visualized under UV light. The primers used for RT-PCR are listed in Table 1.
The List of Primers for RT-PCR
Immunocytochemistry
mpiNSCs were plated on poly-L-lysine-coated 9-mm-wide ACLAR plastic coverslips (SPL Life Sciences, Seoul, Republic of Korea) and incubated in growth medium for 2 days. Then the mpiNSCs were rinsed in phosphate-buffered saline (PBS) and fixed in cold acidic alcohol (5% glacial acetic acid in 95% ethanol) for 10 min at −20°C. After washing, the fixed cells were incubated in blocking solution (10% normal goat serum) at room temperature (RT) for 1 h; this was followed by primary antibodies specific for nestin (1:200; sc-21248; Santa Cruz Biotechnology, Dallas, TX, USA), βIII-tubulin (1:200; MAB1637; Merck Millipore, Darmstadt, Germany), neurofilament [1:200; NF-L (AB9568), -M (AB1987), and -H (AB1989); Merck Millipore], A2B4 (1:200; MAB312R; Merck Millipore), glial fibrillary acidic protein (GFAP; 1:1,000; Z0334; Dako, Glostrup, Denmark), 2′,3′-cyclic-nucleotide 3′-phosphodiesterase (CNPase; 1:200; MAB326; Merck Millipore), Nav 1.1 (1:200; ad106514; Abcam, Cambridge, UK), and Nav 1.6 (1:200; ab83764; Abcam) overnight at 4°C and then labeling with Alexa Fluor 488- or 594-conjugated secondary antibodies (Molecular Probes, Eugene, OR, USA) for 1 h at RT. Cells were mounted using antifading solution containing 4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes) and viewed under a microscope (IX71; Olympus, Tokyo, Japan).
Microarray Hybridization and Data Analysis
To analyze the global gene expression profiles of pmpOBCs and mpiNSCs, Agilent Porcine Gene Expression (V2) 44K GeneChips (Agilent Technologies, Santa Clara, CA, USA) were used for the microarray analyses and processed by eBiogen Inc. (Seoul, Republic of Korea). Double-stranded cDNA was synthesized, amplified, and labeled with the Agilent Low RNA Input Linear Amplification Kit PLUS (Agilent Technologies) from total RNA, and the probes were hybridized using the Agilent Gene Expression Hybridization Kit (Agilent Technologies), followed by washing with the Agilent Gene Expression Wash Buffer Kit (Agilent Technologies). Images were processed using an Agilent DNA microarray scanner and Feature Extraction software (Agilent Technologies). Raw data were normalized using the GeneSpring GX software (ver. 7.0; Agilent Technologies) for analysis.
Statistical Analysis
Differences between pmpOBCs and mpiNSCs were calculated using t-tests as appropriate. All tests were two sided, and statistical significance was set at a threshold of p < 0.05. Statistical analyses were performed using the R program (R foundation, Vienna, Austria). Data are presented as means ± standard error of the mean (SEM).
Results
mpiNSCs Generation
To generate the mpiNSCs, primary neural cells were prepared from minipig olfactory bulb tissue and transfected with a retroviral vector carrying the v-myc oncogene (Fig. 1). Among several stable transfectants isolated after completion of antibiotic selection, a representative clone was used for the subsequent analyses. The two cell types differed slightly in cell morphology. Whereas pmpOBCs were triangular, mpiNSCs exhibited a bipolar morphology with relatively thin, long body shapes (Fig. 2A and B). The RT-PCR analysis showed that v-myc was highly expressed in mpiNSCs (Fig. 2C). In addition, the mpiNSC clone had a normal karyotype (38, XY) (Fig. 2D).

Diagram of v-myc retroviral construction.
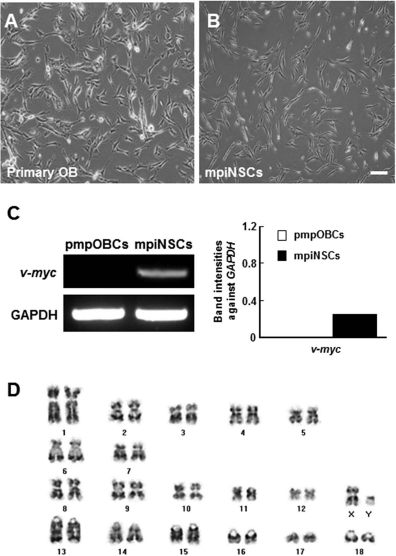
Generation of immortalized minipig neural stem cell (NSC) line (mpiNSCs). (A, B) mpiNSCs had bipolar morphology. Scale bar: 100 μm. (C) RT-PCR analyses indicate that the v-myc gene was overexpressed in mpiNSCs after infection with v-myc retro-virus, but not in primary NSCs. (D) mpiNSCs expressed normal karyotype (38,XY).
mpiNSCs Gene Expression
From the RT-PCR analysis of several cell-specific markers, we found that mpiNSCs expressed markers for NSCs (i.e., Nestin, SOX2, and Vimentin), neurons (i.e., NF and βIII-tubulin), and glia [i.e., GFAP, myelin basic protein (MBP), and CNPase], similar to the pmpOBCs. However, the marker expression levels differed slightly between the pmpOBCs and mpiNSCs. Nestin, SOX2, NF, MBP, receptor of stem cell factor (c-Kit), and stromal cell-derived factor 1 (SDF-1) receptor (CXCR4) were relatively higher, and βIII-tubulin and GFAP were lower in the mpiNSCs. There were no changes in growth factor transcript levels between the two cell types (Fig. 3). In immunocytochemistry analysis using pmpOBCs, specific markers for NSC (nestin and A2B5), neuron (βIII-tubulin and NF), and oligodendrocyte (CNPase) were detected in a large part of cell population, whereas the astrocyte marker (GFAP) was rarely stained (Fig. 4A–F). In addition, we further found that immunoreactivities against NSC and neuronal markers were increased in mpiNSCs compared to those in pmpOBCs (Fig. 4G–L). In contrast, immunoreactivities against astrocyte (GFAP) and oligodendrocyte (CNPase) markers were decreased in mpiNSCs.

RT-PCR of primary OB cells and mpiNSCs. Minipig primary OB and mpiNSCs expressed neural cell markers including nestin, βIII-tubulin, neurofilament (NF), glial fibrillary acidic protein (GFAP), and myelin basic protein (MBP).
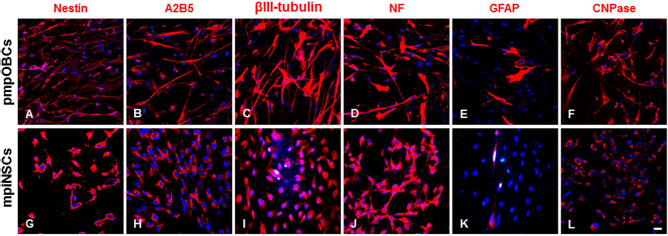
Immunocytochemical characterization of pmpOBCs and mpiNSCs. Immunocytochemistry analysis of markers for NSCs (nestin and A2B5), neurons (β-III tubulin and NF), and glia (GFAP and CNPase) was carried out using pmpOBCs and mpiNSCs. Abbreviations: NF, neurofilament; GFAP, glial fibrillary acidic protein; CNPase, 2′,3′-cyclic-nucleotide 3′-phosphodiesterase. Scale bar: 20 μm.
Further analysis using RT-PCR and immunocytochemistry revealed that neuron-specific sodium ion channel isotypes were differently expressed in pmpOBCs and mpiNSCs. Nav 1.5 was preferentially expressed in pmpOBCs, whereas Nav 1.2, 1.4, and 1.6 were detected only in mpiNSCs. However, high expression levels of Nav 1.1 were detected in both cell types (Fig. 5).

Expression of sodium ion channel in mpiNSCs. To identify the activity of ion channels, Na+ channel activity was measured by RT-PCR and immunocytochemistry. Nav 1.1 and 1.6 were expressed and localized in the nucleus in mpiNSCs. Scale bars: 50 μm.
Global Gene Profiling in mpiNSCs
To examine changes in global gene expression during immortalization, total RNA was isolated from pmpOBCs and mpiNSCs in triplicate and subjected to a micro array analysis using the Agilent Porcine Gene Expression cDNA Chip containing 43,604 genes. On the basis of hierarchical clustering data from Agilent and eBiogen Inc., overall transcript levels of mpiNSCs were downregulated compared with those of pmpOBCs (Fig. 6). Among 7,781 genes that more than doubled their gene expression levels, 515 were upregulated, whereas 7,266 were downregulated.

Gene chip-based clustering data for candidate genes activated in the initial phase of mpiNSCs compared to primary OB cells. To determine the expression level of microarray data, selected twofold changed genes were processed using cluster (version 2.11) and TreeView (ver. 1.6) programs. The figure shows a heatmap of the top up- and downregulated genes of over twofold responded genes of mpiNSCs compared to primary OB cells. The cluster shows the utility of this approach in studying persistent alterations in gene expression. Red and green intensities indicate higher and lower expressions, respectively, relative to median, and black intensity indicates expression near median.
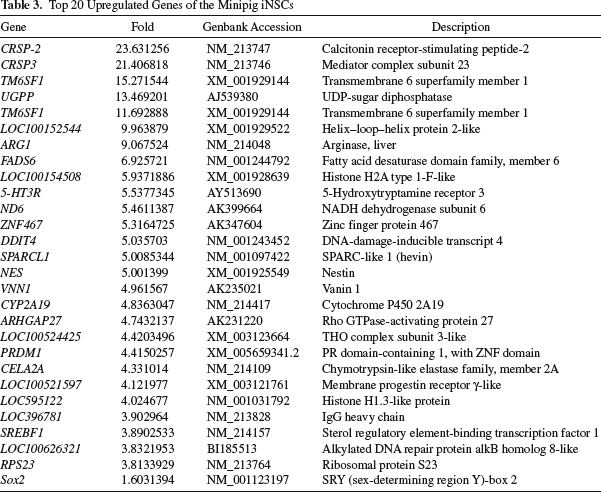
On the basis of the gene ontology analysis (eBiogen Inc., http://www.e-biogen.com), the altered global gene expression profiles were classified into several functional clusters, including neurogenesis (26.1%, 158 of 606 genes), regulation of neurogenesis (27.3%, 72 of 264 genes), stem cell differentiation (28.4%, 61 of 215 genes), stem cell development (31.4%, 54 of 172 genes), Wnt receptor signaling pathway (27.1%, 48 of 177 genes), stem cell maintenance (38.1%, 40 of 105 genes), regulation of Wnt receptor signaling (28.6%, 32 of 112 genes), stem cell proliferation (25.9%, 29 of 112 genes), negative regulation of Wnt receptor signaling pathway (27.3%, 24 of 88 genes), and negative regulation of Wnt receptor signaling pathway (47.1%, 16 of 34 genes) (Table 2). The most up- and downregulated genes are listed in Tables 3 and 4.
Functional Classification of Gene Differential Expression of the Minipig iNSCs

Top 20 Upregulated Genes of the Minipig iNSCs
Top 20 Downregulated Genes of the Minipig iNSCs

From further analysis of the global gene profile, we found that NSC stemness-associated transcript levels were relatively high in mpiNSCs compared to pmpOBCs. Indeed, NSC markers, nestin (5-fold) and Sox2 (1.6-fold) transcripts were upregulated in mpiNSCs, which is greatly similar to the results from the RT-PCR analyses (Fig. 3). In contrast, no changes between both cell types were found in the transcript levels of markers for astrocyte (GFAP; 1.007-fold) and oligodendrocyte (MBP; 1.072-fold). In addition, neurofilament heavy chain was upregulated (1.6-fold) in mpiNSCs, whereas neurofilament light chain was decreased (3.725-fold) in mpiNSCs.
Discussion
In this study, we established a stable immortalized minipig NSC line for the first time via transfection using a v-myc-expressing retroviral vector. The mpiNSCs expressed both NSC and neural markers. Thus, these results suggested that mpiNSCs may be pluripotent and able to self-renew. In addition, we characterized the expression profiles of neurotrophic factors and sodium ion channels. The results indicate that the cell lines can be effectively used for a variety of biomedical studies associated with stem cell therapy.
Minipigs have become increasingly important and useful as large animal models in preclinical studies because of their many similarities to humans in terms of anatomy and physiology6,12. Therefore, there are many studies that have used minipigs in various biomedical applications, including stem cell therapies. For example, Raore and colleagues used a minipig model to assess the long-term risks of multiple intraspinal cell injections 13 . Furthermore, Usvald et al. used an immunosuppressed minipig model to analyze the reproducible therapeutic effects of human spinal stem cells via intraspinal grafting into immunosuppressed minipig models 14 . Although NSCs might be a valuable source of cells for stem cell-based therapies for neurological diseases 15 , there have been relatively few studies on the derivation of immortalized minipig stem cells for preclinical studies 12 .
NSCs can be used to repair damaged brain or spinal cord by replacing degenerated neurons or producing neuroprotective molecules16,17. Several researchers have reported that transplanted NSCs are recruited by signal molecules, such as cytokines, from injured regions18–25. In addition, various cytokines and their receptors were expressed in human F3 NSCs, which were involved in NSC migration toward injury sites. Cytokines are important in increasing the injury-induced proliferation and migration of neural progenitor cells26–28. Moreover, secreted cytokines have a neuroprotective effect in damaged brains 15 . Furthermore, they can provide stable and homogeneous populations with which to investigate neuronal development and neurological diseases 29 . To expand the usability of NSCs, we generated iNSCs based on ease of maintenance and proliferation in vitro.
We found that mpiNSCs expressed several neuro trophic factors, such as brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), bFGF, vascular endothelial growth factor (VEGF), nerve growth factor (NGF), neurotrophin-3 (NT3), and insulin-like growth factor 1 (IGF1). Further studies are needed to determine whether grafted mpiNSCs can be integrated into host brain networks to support the generation of new synaptic connections and replace damaged neurons. Currently, we are working to further define the characteristics of the mpiNSCs using an in vivo transplantation study.
On the basis of the microarray results, 5,011 of 43,604 genes had significantly different expression levels after immortalization. These genes were mainly classified into three categories: neurogenesis, neuronal physiology, and stem cell-related ontology, which comprised ∼9.5% of the total significantly modulated genes. In addition, the top 20 up- and downregulated genes were closely associated with transmembrane receptors, transcription factors, extracellular matrices, physiological enzymes, and histones. These results suggest that the physiological activity and properties of mpiNSCs might differ. Immunocytochemistry analysis showed that most of pmpOBCs were intensively stained with NSC, neuron, and oligodendrocyte markers, whereas astrocyte marker was rarely stained, indicating NSC and neuronal characteristics of pmpOBCs. In addition, we further found that immunoreactivities against NSC and neuronal markers were increased in mpiNSCs compared to those in pmpOBCs. On the contrary, immunoreactivities against glia markers were decreased (Fig. 4). Together with similar results from the RT-PCR analysis (Fig. 3), immunocytochemistry data demonstrated that the v-myc-mediated immortalization process may contribute to enhancement of NSC stemness and neuronal differentiation potential.
mpiNSCs were transduced with v-myc for immortalization. However, recent reports have already revealed that most of the v-myc-transfected cell lines including F3, F3. ChAT, and F3.BDNF cells were successfully differentiated into neurons after transplantation in rodent30–32 as well as nonhuman primate brains 22 . Safety of oncogene has also been debated for a long time. In previous reports, v-myc gene has shown that no tumorigenic phenotypes were proved by transplanting v-myc dependently immortalized human stem cell lines into rodent animal models30–32. Especially, human NSCs can survive for a long time (24 months) in the brain of cynomolgus monkey without tumorigenesis 22 . These results indicated that v-myc can be used as a useful and safe gene to immortalize primary cells.
In this study, we successful generated mpiNSCs with characteristics similar to those of human NSCs, which expressed NSC- and neuron-specific markers, as well as several neuroprotective factors. These results could increase the usability of minipig adult stem cells. Furthermore, the mpiNSCs could be useful in examining the applicability of the immortalization technique to stem cell therapy.
Footnotes
Acknowledgments
This study was supported by grants from the KRIBB Research Initiative Program (KGM4251723 and KGM4611714), Republic of Korea, and the Next-Generation BioGreen 21 Program (No. PJ0099592016), Rural Development Administration, Republic of Korea. The authors declare no conflicts of interest.