
Abstract
Human hepatocytes are used for liver cell therapy, but the small number of engrafting cells limits the benefit of cell transplantation. We tested whether cotransplantation of hepatocytes with hepatic stellate cells (HSCs) could improve hepatocyte engraftment in vivo. Human primary hepatocytes were transplanted into SCID mice either alone or in a mixture with HSCs (quiescent or after culture activation) or LX-2 cells (ratio 20:1). Four weeks after transplantation into mouse livers, human albumin-positive (huAlb+) hepatocytes were found scattered. When cotransplanted in a mixture with HSCs or LX-2 cells, huAlb+ hepatocytes formed clusters and were more numerous occupying 2- to 5.9-fold more surface on the tissue section than in livers transplanted with hepatocytes alone. Increased huAlb mRNA expression in livers transplanted with the cell mixtures confirmed those results. The presence of HSCs increased the number of hepatocytes entrapped in the host liver at an early time point posttransplantation but not their proliferation in situ as assessed by cumulative incorporation of BrdU. Importantly, 4 weeks posttransplantation, we found no accumulation of αSMA+ -activated HSCs or collagen deposition. To follow the fate of transplanted HSCs, HSCs derived from GFP+ mice were injected into GFP- littermates: 17 h posttransplant, GFP+ HSCs were found in the sinusoids, without proliferating or actively producing ECM; they were undetectable at later time points. Coculture with HSCs improved the number of adherent hepatocytes, with best attachment obtained when hepatocytes were seeded in contact with activated HSCs. In vivo, cotransplantation of hepatocytes with HSCs into a healthy liver recipient does not generate fibrosis, but significantly improves the engraftment of hepatocytes, probably by ameliorating cell homing.
Keywords
Introduction
Alternatively to orthotopic liver transplantation, liver cell therapy represents an attractive strategy for treatment of acute liver failure and liver genetic-based metabolic disorders. Human hepatocytes isolated from unused donor livers and from resected liver segments are the main cell source currently investigated for liver cell therapy (1). In human clinical trials, a maximum of 1 to 2 × 108 hepatocytes per kg body weight can be safely transplanted on one occasion, hereby representing ≈5% of total recipient liver cell mass (19). However, replacement of 10% of the hepatocellular mass is needed to compensate for missing or inactive proteins in genetic inborn errors of metabolism (2,8).
Human primary hepatocytes are cryopreserved for banking. However, it is long known that cryopreservation profoundly hampers hepatocyte viability and function, mainly due to mitochondrial dysfunction and failure in ATP production (23). This speaks to the moderate success of clinical transplantation using cryopreserved hepatocytes in inborn metabolic disorders (1,22).
Upon transplantation, the efficiency of liver repopulation is conditioned by the course of several events during the migration, homing, and engraftment of transplanted hepatocytes to the host parenchyma. One of the challenges is the survival of the hepatocytes before they cross the endothelial barrier. More than 70% of transplanted hepatocytes are cleared from the vascular bed within 24 to 48 h after transplantation, even in syngenic hosts (11,21). Surviving hepatocytes must then cross the sinusoid barrier and integrate the liver plate to engraft. Successful engraftment is characterized by adhesion of transplanted cells to native hepatocytes, establishment of cell polarity, and maintenance of mature hepatocyte functions. As shown in mouse experimental models, when engrafted and in the absence of immune rejection, donor human hepatocytes do function in the host liver (6).
As engraftment rate is thought to depend on the capacity of transplanted cells to escape from apoptosis and sinusoidal clearance, experimental procedures aiming at minimizing cell loss during portal–sinusoidal crossing have been used to improve hepatocyte repopulation after transplantation. Such procedures include the use of splanchnic vasodilators to mechanically facilitate cell entry into and across hepatic sinusoids (21) or cotransplantation with bone marrow or mesenchymal stromal cells, which exert an immunomodulatory effect protecting hepatocytes from immune clearance (24). Interestingly, hepatic stellate cells (HSCs) have also been shown to protect grafts from rejection in islet transplantation models through local immune inhibition (5).
The capacity of engrafted hepatocytes to proliferate into the host liver is another crucial parameter. In experimental procedures, significant and long-lasting liver repopulation has been attained when transplanted cells have a proliferative and survival advantage over native cells (10). Those models include enzyme deficiency impairing liver function [such as fumaryl acetoacetate hydrolase (FAH)-/- or the urokinase plasminogen activator (uPA+/+) mouse models] (6,7), or host liver previously exposed to chemical or physical agents such as retrorsine, to block proliferation of host hepatocytes (15).
Studies in a dipeptidyl peptidase IV-deficient rat model showed that activation of HSCs and matrix remodeling in the recipient liver, events triggered by and transiently observed after cell transplantation, are helpful for cell engraftment (3). Other strategies such as cell coating in extracellular matrix (30) or intraportal injection of matrix components (14) appear to provide a protective environment for transplanted hepatocytes.
Based on all those observations, we tested here the hypothesis that hepatic stellate cells improve engraftment of hepatocytes.
Materials and Methods
Animals
Immunodeficient SCID mice (females, 20-24 g) (Charles Rivers, L'Arbresle, France) were used as recipients for transplantation. GFP+ transgenic male mice (males, 35-44 g), kindly gifted by Prof. A Goffinet (Unit of Developmental Neurobiology, Institute of Neurosciences, Université catholique de Louvain, Brussels, Belgium) were used as cell donors (29). Their wild-type littermates (WT) were used as recipients for syngenic transplantation. Mice were housed and handled in accordance with local ethical guidelines of the laboratory animal care of the Université catholique de Louvain, and the study protocol has been approved by the local ethics committee (UCL/MD/2009/017 and UCL/MD/2012/026).
Cell Isolation
Human material was used with permission of the local ethics committee and experiments conducted in accordance with human and animal ethical guidelines of the Université catholique de Louvain. Human tissues were obtained from the Health Ministry-accredited tissue bank at the center of cell therapy from Cliniques Universitaires Saint Luc. Tissues authorized for research use were obtained from healthy liver donors (n=5) or from explanted livers with a single metabolic defect (n=3) (Table 1). Infant donors were non-heart-beating donors. Liver tissue was harvested 15 to 30 min after cardiac arrest, following a procedure approved by the St-Luc Hospital Ethical Review Board and after obtaining informed consent of the families. Quality control of the isolated liver cells is systematically performed and meets the release criteria of the Hepatocyte and Hepatic Stem Cell Bank.
Characteristics of the Donors Used in the Study

F, female; M, male; nr, not relevant.
For in vitro studies.
For transplantation study.
Human hepatocytes were isolated using a classical two-step collagenase digestion of liver segments (25), modified as previously described (18). The hepatocyte parenchymal fraction (pellet) was progressively enriched by a gradual filtration and by several washes and low-speed centrifugation steps. Hepatocytes were used immediately after isolation or cryopreserved at a concentration of 1 × 106 cells per milliliter. Hepatocyte viability was evaluated using the trypan blue (Sigma-Aldrich, Germany) exclusion assay after isolation and after thawing.
The nonparenchymal fraction (supernatants collected after wash/centrifugation steps) was pelleted and cryopre-served. After thawing, the nonparenchymal cell fraction was centrifuged on a Nycodenz (Sigma-Aldrich, Irvine, UK) gradient, and the HSC-enriched fraction collected following a modified mouse HSC isolation protocol as previously published (9). The so-obtained human HSCs have been characterized elsewhere (4).
Mouse hepatocytes were isolated from GFP+ transgenic mice as follows. Livers were perfused in situ through the vena cava with Krebs–Ringer containing, consecutively, EGTA (0.1 mM; Sigma-Aldrich, Germany) then 0.07 mg/ml Liberase Blendzymes (Roche, Mannheim, Germany), and 0.2 μg/ml calcium (Sigma-Aldrich). Liver tissue was mechanically disrupted in calcium containing Krebs–Ringer buffer, filtered through a 100-μm pore size cell strainer (BD Biosciences, Erembodegem, Belgium), and the hepatocyte fraction was enriched by two wash/centrifugations at 20 × g, 5 min, in RPMI (Invitrogen, Merlebeke, Belgium) supplemented with 10% fetal bovine serum (FBS) (Invitrogen).
Mouse HSCs were isolated from GFP+ transgenic mice by a two-step pronase–collagenase digestion as previously described (9). The human HSC-derived cell line LX-2, generated by transformation of cultured primary HSCs obtained from a male human liver (28), was kindly provided by S. Friedman (Mount Sinai School of Medicine, New York, NY, USA).
Cell Transplantation
Recipient SCID mice were anesthetized with ketamine (Pfizer, Brussels, Belgium)/xylazine (Bayer Healthcare, Diegem, Belgium) before transplantation of donor cells via the spleen.
Human cryopreserved hepatocytes were transplanted alone (1 × 106 Hep) or in a mixture with HSCs (1 × 106 Hep; 5 × 104 HSCs, Hep+HSC). We used freshly isolated quiescent (q) or culture-activated (P7 in culture-a) primary human HSCs (from donors 1 and 8) or LX-2 cells (immortalized human activated HSC line) and repeated the experiment with three batches of hepatocytes from donors 2, 8, and 9. In other experiments, we transplanted mouse primary GFP+ hepatocytes with or without culture-activated primary mouse GFP+ HSCs (at passage 2). Viability of thawed hepatocytes declines rapidly. For that reason, cells had to be used within 2 h after thawing allowing for transplantation of 12 animals. In all experiments, injection of HSCs alone (HSC controls) was performed at the end of the transplantation session, as we did not observed significant change in cell viability after storage of the cell suspension on ice for up to 5 h. In some experiments, BrdU (Sigma-Aldrich)-charged osmotic pumps (Alzet®, model 1004) were implanted into the peritoneal cavity at the time of transplantation to assess 4-week cumulative cell proliferation. Animals were sacrificed 6 h or 4 weeks posttransplantation.
To evaluate the fate of transplanted HSCs, WT mice were transplanted with 5 × 104 GFP+ quiescent or activated HSCs (isolated from GFP+ transgenic mice), and animals were sacrificed 17 h, 1 week, or 4 weeks after transplantation (n=4 per group).
In all experiments, we transplanted 1 × 106 hepatocytes, 1 × 106 Hep+5 × 104 HSCs for the cell mixture, or 5 × 104 HSCs alone per mouse in a volume of 100 μl phosphate-buffered saline (PBS) (Invitrogen). The ratio of Hep/HSC in the cell mixture was 20:1 mimicking the hepatocyte to HSC ratio in the liver parenchyma. Control mice (CTL, n=3) were injected with 100 μl PBS.
At the time of sacrifice, livers were excised. A small portion of each lobe was immediately snap frozen in liquid nitrogen and kept at −80°C until use. The rest of the liver was fixed in 10% formalin (Labonord, Templemars, France) for histology. Each lobe was sliced into two to five pieces and mounted in paraffin blocks to largely sample all liver lobes.
Histology and Immunostaining
Formalin-fixed and paraffin-embedded 5-μm-thick liver sections were deparaffinized and rehydrated in graded alcohol baths. Sirius red was prepared with 500 mg Direct Red 80, 500 mg Fast Green FCF, and 500 ml of saturated picric acid (all purchased from Sigma-Aldrich), and staining was performed following standard protocol. For immunostainings, endogenous peroxidases were blocked in 3% H2O2 methanol (both from VWR BDH Prolabo, Fontenay-sous-Bois, France) and antigen retrieval performed in citrate buffer (pH 8.0) for 45 min at 100°C. The citrate buffer was prepared with 18.3 g citric acid monohydrate, 140.7 g trisodium citrate, 4.8 g Tris buffer (all from Sigma-Aldrich), and 1 L of deionized water. Sections were covered with 5% normal goat serum (Invitrogen) at room temperature followed by overnight incubation at 4°C with primary antibodies against human albumin (huAlb, no cross reactivity with mouse albumin) (126584; 1:2,500; Calbiochem, Darmstadt, Germany), human ornithine transcarbamylase (huOTC) (1:200; Sigma-Aldrich), and/or BrdU (1:100; Dako, Glostrup, Denmark). For CD10 and E-cadherin immunodetection, sections were incubated with mouse IgG blocking reagent (Vector Laboratories, Burlingame, CA, USA) then with anti-human/mouse CD10 (1:50; Thermoscientific, Waltham, MA, USA) or anti-human/mouse E-cadherin (1:100; BD Biosciences, Belgium) overnight. Anti-rabbit or anti-mouse Envision (Dako) was used for detection.
GFP immunostaining was used to identify GFP+ mouse cells in GFP- recipient livers. Briefly, liver sections were incubated overnight at 4°C with anti-GFP antibody (1:250; Abcam, Cambridge, UK) and detected using peroxidase-coupled anti-goat secondary antibody (1:200; Dako).
Peroxidase activity was revealed with diaminobenzidine (Dako, Carpenteria, CA, USA) and sections counter-stained with hematoxylin (Labonord). We used anti-rabbit Alexa Fluor 488 or anti-mouse Alexa Fluor 594 secondary antibodies (1:1,000; Invitrogen) for immunofluorescent analyses with diamidino-2-phenylindole (DAPI) (Sigma-Aldrich) counterstaining.
Morphometric Analysis
Morphometric quantification (huAlb+, huOTC+, GFP+ areas) was performed using a Zeiss microscope coupled to an Axiocam camera (MR3; Carl Zeiss, Munich, Germany) and Axiovision software (Zeiss). We analyzed ≈20 images per liver lobe (≈100 images per liver), covering the entire surface of the sections (200x magnification). Positively stained area was expressed as a ratio of stained area to the total image area, leaving out white and vascular areas. The percentage of stained areas was recorded for each lobe, and an average was made per liver.
Coculture Experiments
Human and mouse primary HSCs were used immediately after isolation (“quiescent cells”) or after culture-activation (one to seven passages, “activated cells”) in DMEM (Invitrogen) supplemented with 20% FBS (Invitrogen) and penicillin/streptomycin (Invitrogen). LX-2 cells were cultured in DMEM supplemented with 5% FBS and penicillin/streptomycin (Invitrogen) (26).
Freshly isolated and cryopreserved/thawed human and mouse hepatocytes were cultured in William's E medium (Invitrogen) supplemented with 10% FBS, 1 μM dexamethasone (Schering-Plough, Brussels, Belgium), 10 μg/ml insulin (Sigma-Aldrich), 25 ng/ml epidermal growth factor (EGF) (Invitrogen), and penicillin/streptomycin at a density of 5 × 104 hepatocytes per cm2 on plastic 24-well cell culture plates (CellBind®; Corning Inc., Corning, NY, USA), alone or in coculture with HSCs or LX-2 (2.5 × 103; 20:1 Hep:HSC ratio). Cell attachment was evaluated 24 h postplating indirectly by counting floating cells and directly by quantification of attached cells using the crystal violet colorimetric assay (12). Briefly, cells were fixed in 10% formalin and incubated with crystal violet dye (0.05%; Sigma-Aldrich). The amount of dye retained is proportional to the number of adherent cells. After washing, the dye was eluted in methanol (VWR BDH Prolabo, Fontenay-sous-Bois, France) and quantified spectrophotometrically (540 nm). The contribution of the HSCs to attached cells, in wells in which hepatocytes were seeded in contact with HSCs, was taken into account by subtracting the spectrophotometric values of HSCs alone from the value of the Hep + HSCs. Therefore only attached hepatocytes were considered.
Immunocytofluorescence
Seven days after seeding, cells were fixed with 10% formalin for 5 min, washed with PBS (Invitrogen), and permeabilized for 2 min at room temperature in PBS with Triton X-100 (0.3%; Merck, Darmstadt, Germany). After incubation in 5% normal goat serum, primary antibodies against human albumin (1:2,500; Calbiochem) and αSMA (1:200; Dako) were added to incubate overnight at 4°C. Secondary antibodies anti-rabbit Alexa Fluor 488 and anti-mouse Alexa Fluor 594 (1:1,000; Invitrogen) were used for detection (1 h, room temperature) with DAPI counterstaining.
Evaluation of Cell Proliferation
Human Alb and BrdU double immunofluorescent staining was performed on 5-μm liver sections from different hepatic lobes per animal. Pictures of five to seven fields containing huAlb+ cells were taken (magnification 200x). The ratio of the number of BrdU+/huAlb+ human hepatocytes to the total number of huAlb+ human hepatocytes was expressed as a percentage. An average was made for each animal. Results are presented as an average value per animal group (Hep, n=4; Hep + HSCs, n=5).
RNA Extraction and RT-qPCR
We used human albumin, human/mouse (RPL19, αSMA, collagen-α1, MMP9), and mouse TIMP-1 primers. Total RNA was extracted and RT-qPCR analysis performed as previously described (16). Primers were designed using Primer Express software (Applied Biosystems, Lennik, Belgium) and are listed in Table 2. Mouse or human RPL19 mRNAs were chosen as invariant standards as appropriate. Results are expressed as fold expression relative to expression in human tissue (value arbitrarily set at 1,000 for human albumin) using the ΔΔCt method (17) or as mean Ct value.
Statistical Analysis
Data are as mean+SEM in bar graphs, and median, minimum, and maximum in box and whisker graphs. When there was a small sample size in which the normal distribution could not be attested and/or the presence of outliers with undetectable or null values, the nonparametric Mann–Whitney test was used for simple comparison and, for multiple comparison, the Kruskal–Wallis test (p provided in the text) followed by Mann–Whitney test with sequential Bonferroni correction for planned comparison between cell mixture groups (Hep + HSC or Hep + LX-2) versus hepatocyte alone (statistical significance denoted in the figures). huAlb+ area is a dependent variable, and so it is continuous and normally distributed (Kolmogorov–Smirnov test and D'Agostini and Pearson omnibus K2 normality test), and the interaction with independent variables (timing posttransplantation and transplanted cell mixture) was tested using the robust two-way ANOVA. Then to compare the two timings for the same cells and the two cell types for the same timing, we used the t-test and applied the Bonferroni correction for multiple comparisons. Statistical significance is denoted in the figures. The mean number of floating hepatocytes and the mean quantification of adherent hepatocytes were analyzed using the paired t-test. Values of p < 0.05 were considered as statistically significant. GraphPad Prism software (San Diego, CA, USA) was used for graphs and statistics.
Results
In Vivo, Cotransplantation with HSCs Improves Human Hepatocyte Repopulation of the Host Liver in Nonselective Conditions
To test whether HSCs will improve engraftment of hepatocytes, we transplanted human cryopreserved primary hepatocytes alone (1 × 106 Hep) or in a mixture with freshly isolated quiescent human HSCs (1 × 106 Hep: 5 × 104 qHSC, Hep + qHSCs) or with LX-2 cells (1 × 106 Hep: 5 × 104 LX-2, Hep+LX-2) through splenic injection into unharmed SCID mice (Fig. 1). We observed a 50% drop in hepatocyte viability when the thawed hepatocytes were kept on ice for 3 h (not shown). Therefore, to keep the total transplantation time below 2 h, we limited to 12 (n=4 per group) the number of animals to be transplanted with hepatocytes from one thawed hepatocyte cryovial. Controls received the same number of qHSCs or LX-2 cells alone. Four weeks after transplantation of human hepatocytes alone, human Alb+ hepatocytes were found scattered in the parenchyma of recipient livers. However, they formed clusters and were more numerous when they were transplanted in mixture with qHSCs and to a greater extent when transplanted with LX-2 cells (Fig. 1A). Morphometrical analysis of huAlb+ area revealed differences between groups (p=0.0014 using Kruskal–Wallis test) and confirmed a significantly higher number of engrafted hepatocytes when cotransplanted with LX-2 cells compared to hepatocytes alone (Fig. 1C). The expression of the huAlb gene correlated with huAlb+ morphometry (Spearman r=0.87, p< 0.0001) (Fig. 1E). The expression of the huAlb gene was different between groups (p = 0.0022 using Kruskal–Wallis test) and higher in Hep+LX-2 than in Hep recipient livers, but after correction for multiple testing, the difference was no longer statistically significant (Fig. 1D). huAlb+ cells and amplification of human albumin mRNA were not found in CTL livers, nor in livers transplanted with LX-2 cells or HSCs alone (Fig. 1A, C, and D). Transplanted hepatocytes also expressed other features of mature, functional hepatocytes such as human OTC both in Hep and Hep+HSC livers (Fig. 1A) with a good correlation between huOTC+ and huAlb+ immunoreactive area on liver section (Fig. 1F). As also shown by CD10 immunodetection, engrafted hepatocytes were polarized and formed bile canaliculi with both transplanted and host adjacent hepatocytes (Fig. 1B).
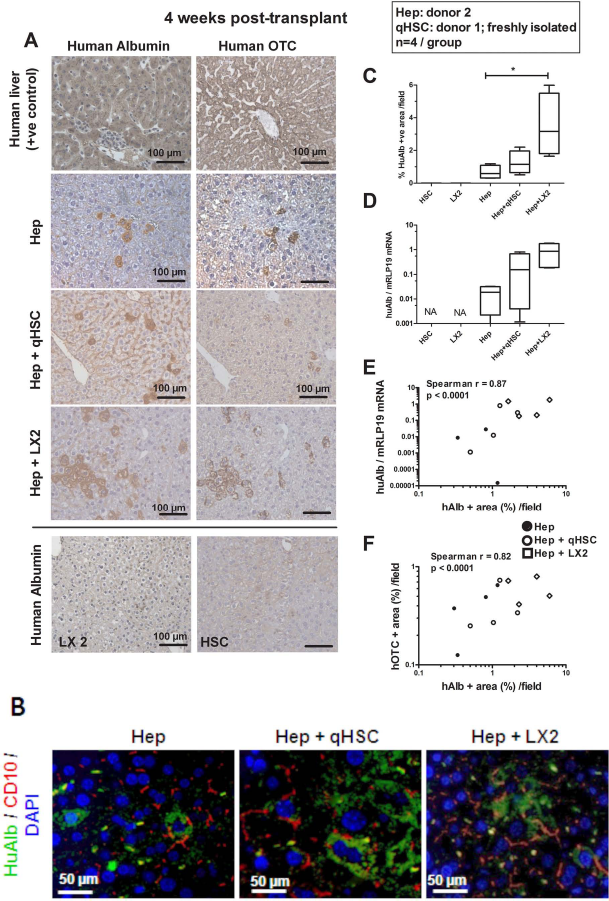
Cotransplantation with human hepatic stellate cells (HSCs) increases engraftment of human hepatocytes. Human cryopreserved hepatocytes were from donor 2, and human primary quiescent stellate cells were from donor 1. (A) Representative pictures of human albumin (huAlb) and human ornithine transcarbamylase (OTC) immunohistochemistry on human liver tissue (positive control), on consecutive sections of livers transplanted with human cryopreserved hepatocytes alone (Hep), Hep in the mixture with human primary quiescent HSC (Hep + qHSC) or with LX-2 cells (human HSC-derived cell line) (Hep + LX-2), and on a section of liver transplanted with LX-2 cells alone (LX-2), 4 weeks posttransplantation; magnification 200x (scale bar: 100 μm). (B) Pictures of human albumin/cluster of differentiation 10 (CD10) double immunostaining in sections of Hep, Hep + qHSC, and Hep + LX-2 transplanted livers; magnification 400x (scale bar: 50 μm). Graphs representing (C) morphometric analysis of huAlb+ surface and (D) HuAlb gene expression normalized to mouse RPL19 mRNA (regarded as invariant control for normalization of RNA load) for qHSC (n = 3), LX-2 (n = 3), Hep (n = 4), Hep + qHSC (n = 4), and Hep + LX-2 (n = 4) liver sections, 4 weeks posttransplantation. Box and whiskers (min–max). Data for RNA are fold increased using as reference human liver mRNA expression with value arbitrarily set at 1,000. Data analyzed using Kruskal–Wallis test followed by Mann–Whitney test with sequential Bonferroni correction for planned comparisons between cell mixture groups (Hep + HSC and Hep + LX2) versus hepatocyte alone (*statistical significance in graphs). Graphs representing (E) correlation between huAlb+ area and huAlb mRNA, and (F) correlation between huAlb+ surface and human OTC+ surface for Hep (•, n = 4), Hep + qHSC (○, n = 4), and Hep + LX-2 (◊, n = 4) livers, 4 weeks posttransplantation. Correlations were analyzed using Spearman test.
List of Primers Used in the Study

RPL, ribosmomal protein L19; MMP9, matrix metalloproteinase 9; αSMA, α smooth muscle actin; TIMP-1, tissue inhibitor of metalloproteinases 1.
In two subsequent independent experiments, hepatocytes from donors 8 and 9 were transplanted alone (n=8) or together with human activated stellate cells (P7 in culture, n=9). One month posttransplantation, the number of huAlb+ cells and the level of expression of huAlb mRNA were significantly higher in livers transplanted with the cell mixture than with hepatocytes alone (Mann–Whitney test; Fig. 2A, B).
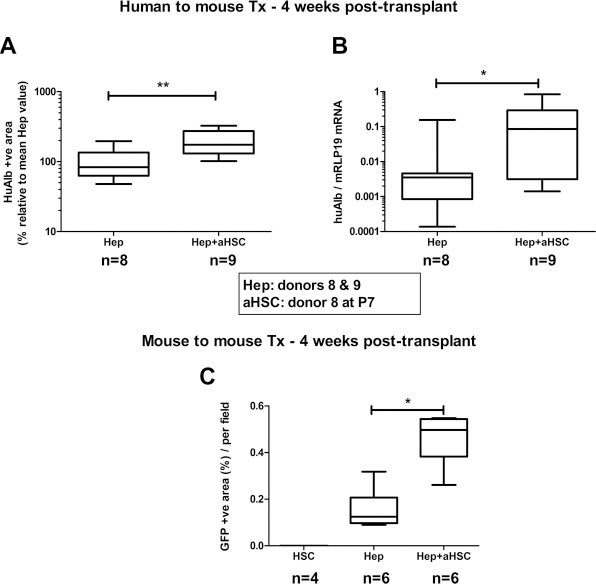
Activated HSCs enhance engraftment of human and mouse primary hepatocytes. (A) Graph representing morphometric analysis of huAlb+ surface for livers of severe combined immunodeficient (SCID) mice transplanted with human cryopreserved hepatocytes alone [Hep; from donor 8 (n = 4) and from donor 9 (n = 4)], or in mixture with human primary activated HSCs from donor 8 at passage 7 [Hep + aHSC; using Hep from donor 8 (n=4) and Hep from donor 9 (n=5)], 4 weeks posttransplantation. The mean Hep value for each donor was arbitrarily set at 100%. **p < 0.001 (Mann–Whitney test). (B) Graph representing huAlb mRNA expression in the same livers. *p < 0.05 (Mann–Whitney test). (C) Graph representing morphometric analysis of green fluorescent protein-positive (GFP+) surface for livers of SCID mice transplanted with GFP+ mouse activated HSCs alone (aHSC), with GFP+ mouse hepatocytes alone (Hep, viability of 81.8% before transplantation; n=6), or with Hep in mixture with GFP+ mouse aHSCs at passage 2 (Hep + aHSC; n=6) 4 weeks posttransplantation. Data analyzed using Kruskal–Wallis test (p=0.0005) followed by Mann–Whitney test with sequential Bonferroni correction.
Finally, GFP+ mouse hepatocytes (isolated from GFP+ Tg mice) were transplanted alone (n=6) or in a mixture with GFP+ murine HSCs (P3 in culture, n = 6) into SCID mice. Implantation of cells was determined by tracking of GFP+ cells in the host liver. One month posttransplantation, we observed significantly higher hepatocyte engraftment when cotransplanted with HSCs than when transplanted alone, while we detected no GFP+ cells in liver transplanted with mHSCs alone (p = 0.0005 using Kruskal–Wallis test) (Fig. 2C).
Improved Repopulation in HSC-Hepatocyte Cotransplanted Livers Is Related to Increased Homing
In order to determine whether increased repopulation of the host liver by transplanted hepatocytes in the presence of HSCs was related to a better homing of transplanted cells or to increased proliferation of homed cells, we analyzed livers 6 h and 4 weeks posttransplantation, in two series of transplantation experiments using hepatocytes from donors 8 and 9 and activated HSCs from donor 8 at passage 7. The experiment using hepatocytes from donor 8 was duplicated (two different vials of cryopreserved hepatocytes thawed and transplanted on different days). Six hours after transplantation, some huAlb+ hepatocytes were preferentially found in periportal parenchyma (Fig. 3A). Large clusters of human Alb+ cells were more readily found in livers transplanted with Hep+aHSC mixtures than when hepatocytes were transplanted alone. The number of hepatocytes entrapped in the liver was higher in Hep+aHSC livers compared with Hep livers (Fig. 3A, B). We further analyzed the integration of engrafted hepatocytes in the host liver parenchyma. Already at this early time point, huAlb+ hepatocytes had crossed the endothelial barrier and integrated the liver plate, restored their plasma membrane structures and polarity, and formed hybrid CD10+ bile canaliculi with adjacent transplanted hepatocytes (Fig. 3A). Engrafted hepatocytes also formed E-cadherin+ adherent junctions with adjacent donor or mouse hepatocytes (Fig. 3A). We then compared observations at the 6-h time point with those at the 4-week time point. While at 6 h posttransplantation human hepatocytes were preferentially seen in periportal areas, at 4 weeks they were found scattered in the entire lobule and no longer confined to the perivascular zones. Interestingly, however, only the cell/cell mixture used for transplantation, but not the timing of observation, significantly influenced the extent of parenchyma huAlb+ surface (Fig. 3B).

Increased engraftment is due to better homing of transplanted human hepatocytes into host mouse liver in the presence of HSCs. (A) Pictures of livers transplanted with human cryopreserved hepatocytes alone (Hep) or in the mixture with human activated HSCs (Hep + aHSC), 6 h posttransplantation, and representing human albumin (huAlb) immunohistochemistry (upper panels, magnification 100x, scale bar: 200 μm); representative pictures of human albumin/CD10 double immunostaining (middle panels, magnification 200x, scale bar: 100 μm, far left and far right panel are enlarged pictures as delineated), yellow arrows point toward bile canaliculi between donor hepatocytes and white arrows toward hybrid bile canaliculi between donor and mouse hepatocytes; and representative pictures of human–mouse E-cadherin/huAlb double immunostaining (lower panels, magnification 400x, scale bar: 100 μm), white arrows point toward hybrid adherent junction between donor and mouse hepatocytes. (B) Graphs representing huAlb morphometric analysis of livers transplanted with human primary activated HSCs from donor 8 at passage 7 (HSC), human cryopreserved hepatocytes alone (Hep), and in the mixture with human primary activated HSCs (donor 8, passage 7) (Hep + aHSC), evaluated after 6 h [HSC, n = 6; Hep, n = 14, Hep + HSC, n = 14 (hepatocyte donor 8 vial 1, n = 4; donor 8 vial 2, n = 5; donor 9 n = 5)] and after 4 weeks [HSC, n = 6, Hep, n = 8, Hep + HSC, n = 9 (hepatocyte donor 8 n = 4; donor 9, n = 5)]. Values were normalized on the 6-h mean value of Hep livers, in each experiment. Interactions were analyzed by two-way ANOVA; differences between groups using t-test with Bonferroni correction. *p < 0.05 and **p < 0.001. (C) Representative pictures of huAlb/BrdU double immunostaining of livers transplanted with human cryopreserved hepatocytes alone (Hep) and in the mixture with human primary activated HSCs (Hep + HSC), 4 weeks posttransplantation (magnification 400x, scale bar: 50 μm). (D) Graph representing the cumulative incorporation of bromodeoxyuridine (BrdU) into transplanted cells 4 weeks posttransplantation expressed as the percentage of huAlb+/BrdU+ cells (100% is the total number of huAlb+ cells) for Hep (n = 4) and Hep + HSC (n = 5) livers. No statistical difference (Mann–Whitney test).
As mice were implanted a BrdU-filled minipump at the time of transplantation (Hep n = 4, Hep + HSC n = 5), we performed huAlb/BrdU double staining on 4-week recipient livers to assess proliferation of transplanted cells (Fig. 3C). We observed that a similarly low number of engrafted hepatocytes had incorporated BrdU in both Hep and Hep + aHSC livers (Fig. 3D).
Coculture with HSC Improves Attachment of Cryopreserved Hepatocytes
It is well documented that cryopreservation hampers cell survival and function of human hepatocytes (23).
We next tested whether HSCs modify plating and survival of human hepatocytes in vitro. Using hepatocytes from five different donors, we repeatedly observed that the number of nonadherent hepatocytes was reduced when they were seeded together with HSCs compared to cryopreserved hepatocytes seeded alone (paired t-test; Fig. 4A). In accordance, the number of adherent cells was significantly higher in coculture conditions (paired t-test; Fig. 4B). Improvement of cell attachment was observed irrespective of the viability of hepatocytes as determined before plating (Table 1) and of whether quiescent or culture-activated human HSCs or LX-2 cells were used. In addition, the presence of quiescent HSCs also further increased seeding of freshly isolated human hepatocytes (open triangles in Fig. 4A, B). In contact with human HSCs, human hepatocytes formed clusters (huAlb+) surrounded by activated HSCs (αSMA+), whereas they appeared less numerous and more scattered when hepatocytes were seeded alone (Fig. 4C).

Coculture with quiescent or activated hepatic stellate cells (aHSCs) improves the attachment of primary hepatocytes. Graphs representing (A) the mean number of floating hepatocytes and (B) the mean quantification of adherent hepatocytes based on crystal violet staining analyzed 24 h postplating for fresh (from donor 1) and cryopreserved (from donors 2, 3, 4, or 5) hepatocytes seeded alone (Hep), in contact with primary HSCs (quiescent: from donors 1, 6, or 7; or culture activated: from donor 8, at passage 7) or with LX-2 cells (Hep + HSC). The quantification of hepatocyte attachment in Hep + HSC wells was obtained after subtracting HSC crystal violet spectrophotometric values from Hep + HSC values. *p < 0.05 indicates statistically significant difference (paired t-test). (C) Human cryopreserved hepatocytes (donor 2) were cultured alone (Hep) or with human primary quiescent HSCs (donor 1) and were stained with human albumin (green) and α smooth muscle actin (aSMA; red) 7 days after seeding. Magnification 400x; Scale bar: 50 4mUm. (D) Graph representing quantification of adherent GFP+ hepatocytes (Hep) when cultured alone or in contact with quiescent (Hep + qHSC) or with culture-activated GFP+ HSC (Hep + aHSC). Hep, qHSCs, and aHSCs were isolated from different mouse livers. Data are normalized on hepatocytes alone (mean value set at 1one) and presented as fold change for the association with HSC compared to hepatocytes alone. Points represent independent experiment, bar is mean ± SEM. Data were analyzed using Kruskal–Wallis (p = 0.029) followed by Mann–Whitney test with sequential Bonferroni correction.
We next wanted to clarify whether quiescent or activated HSCs would differentially influence the attachment of hepatocytes. For logistical reasons and to allow for better comparison, those experiments were performed using cells derived from GFP+ transgenic mice. Four individual experiments were performed using hepatocytes isolated from four different mouse livers. Mouse primary hepatocytes were cultured alone and in coculture with quiescent or culture-activated (passage 3) mouse primary HSCs. The presence of HSCs increased the attachment of hepatocytes (p = 0.03 using Kruskal–Wallis test) although with no statistical advantage of activated over quiescent HSCs (Fig. 4D).
Transplanted HSCs Do Not Trigger Fibrosis in the Host Liver
HSCs are the major matrix-producing cells in the liver, and implantation of exogenous HSCs may trigger fibrosis. To follow the fate of HSCs, we transplanted 5 × 104 quiescent and culture-activated GFP+ HSCs into WT GFP- healthy livers and traced transplanted cells using anti-GFP immunohistochemistry. Seventeen hours posttransplantation, HSCs were found in host sinusoids. After 1 or 4 weeks, GFP+ cells were no longer detected in recipient livers (Fig. 5A). Also, more importantly, no abnormal matrix deposition was found in HSC-transplanted livers (Fig. 5B).

Transplanted mouse GFP+ HSCs are cleared from the host liver without generating fibrosis. (A) Representative pictures of GFP-stained sections of mouse GFP- livers transplanted with GFP+ mouse-derived quiescent HSCs or activated HSCs, 17 h (left panels, magnification 400x, scale bar: 50 μm) and 4 weeks (middle panels, magnification 100x, scale bar: 200 μm) posttransplantation. Arrows indicate GFP+ cells in the host liver sinusoidal space 17 h after transplantation. (B) Sirius red staining in the same conditions 4 weeks posttransplantation (magnification 200x, scale bar: 100 μm).
Four weeks posttransplantation of human cryopreserved hepatocytes with HSCs, we found no accumulation of αSMA+ cells or extracellular matrix deposition in the vicinity of engrafted hepatocytes or elsewhere in the liver lobule (Fig. 6A). Also, there was no amplification of human fibrotic genes MMP9, αSMA, or collagen-α1 in transplanted livers (Table 3). This contrasted with detection of huAlb mRNA and huRPL19 mRNA (a human housekeeping gene) in mouse livers transplanted with human hepatocytes (with or without HSCs or LX-2), but not in those transplanted with human HSCs or LX-2 cells alone (Figs. 6B and 1D). Similarly, the expression of mouse profibrogenic genes was not induced 4 weeks after transplantation, irrespective of transplanted cells (not shown).
Amplification of Human Matrix-Related Genes (Ct Values) in Recipient Mouse Livers 4 Weeks Posttransplantation

NA, not amplified; HSC, hepatic stellate cell; LX-2, human HSC-derived cell line; Hep, hepatocyte.
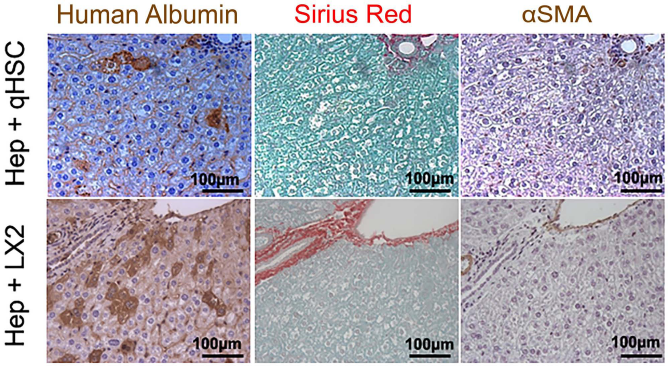
Fibrosis does not develop in livers transplanted with cell mixture containing HSCs. Representative pictures of consecutive sections of liver transplanted with the mixture of human cryopreserved hepatocytes with human primary quiescent HSCs (Hep + HSC) or with LX-2 cells (Hep + LX-2) 4 weeks posttransplantation stained for human albumin (left panels), Sirius red (middle panels), and αSMA (right panels). Magnification 200x (scale bar: 100 μm).
Discussion
We evaluated the benefit of interaction of hepatocytes with hepatic stellate cells for liver cell therapy. We showed by three independent experiments that cotransplantation of human cryopreserved hepatocytes in a mixture with HSCs enhanced the number of hepatocytes engrafted into mouse liver.
Currently, results of liver cell therapy are hampered by the relatively poor efficiency of hepatocyte engraftment and limited long-term survival of transplanted cells. Previous studies demonstrated that cryopreservation of hepatocytes impaired both their attachment and function (27). We demonstrated that coculture with human HSCs improved the attachment of human hepatocytes. Similarly, using mouse cells, attachment of hepatocytes was significantly higher when plated with activated HSCs, but not with quiescent HSCs. This suggests that factors and/or matrix overproduced by activated HSCs favor hepatocyte attachment. This is in accordance with previously published results showing a better maintenance of hepatocytes in vitro when seeded on collagen or on defined extracellular matrix (20). Recently, we analyzed the secretome of human primary HSCs in culture and demonstrated that, besides producing matrix, they significantly overexpress and release cytokines and growth factors such as transforming growth factor-β, hepatocyte growth factor, platelet-derived growth factor, or interleukin-6, (4) which may modulate hepatocyte attachment and survival.
In vivo, one of the fundamental challenges for hepatocyte engraftment into the host liver parenchyma is their survival. We showed by three independent experiments that cotransplantation of human cryopreserved hepatocytes in a mixture with HSCs enhanced the number of hepatocytes engrafted into mouse liver. We correlated this finding with a higher expression of human albumin mRNA in livers transplanted with the hepatocyte/HSC mixture. Besides expressing albumin, those hepatocytes also express other hepatocytic enzymes, such as urea cycle ornithine transcarbamylase (OTC), and formed bile canaliculi with adjacent hepatocytes, suggesting that engrafted human hepatocytes are terminally differentiated and equipped to participate in recipient mouse hepatic function. The observation that HSCs improve engraftment efficiency was further confirmed by using a mouse-to-mouse transplantation model with tracking of GFP+ hepatocytes to assess engraftment.
As early as 6 h posttransplantation, human hepatocytes were already seen in the host parenchyma and in higher numbers in the presence of HSCs. They were preferentially seen in portal areas and were already integrated into the mouse hepatocyte plate as they were adherent, polarized, and formed bile canaliculi with adjacent hepatocytes. The fact that other studies using syngenic transplantation of rat hepatocytes in DPPIV-deficient rat recipients reported that a delay as long as 16 h is required for integration of donor cells into host hepatocyte plates (11,13) suggests that the timing for engraftment may vary depending on the nature/strains of both donors and recipients. In accordance with that, when performing syngenic transplantation of GFP+ hepatocytes into GFP- mice, we found that donor GFP+ hepatocytes remained in the vascular space for at least 17 h, unable to cross the endothelium, causing little engraftment rate (not shown). This was absolutely not the case when SCID mice were used as recipients for human hepatocytes or GFP+ mouse hepatocytes; the crossing of endothelial barrier and recovery of polarity and the integration to the liver plate happened within 6 h after transplantation.
We observed thus that the presence of HSCs contributed to better entrapping of transplanted hepatocytes into host liver plates in the initial events posttransplantation. Thereafter, the number of hepatocytes engrafted remained steady. We saw no difference in DNA synthesis in implanted hepatocytes whether transplanted with or without HSCs, supporting that the additional cell type did not promote proliferation of grafted hepatocytes. Rather our observations support that better transplantation efficiency was conditioned by initial homing. The prerequisite for homing and integration into liver plates is for the hepatocytes to cross the endothelial barrier (11,21). Cells incapable of engrafting are rapidly cleared after cell transplantation (11). Thus, our observation at an early time point supports that HSCs have allowed a large number of hepatocytes to cross the sinusoidal endothelial barrier. As supported by our in vitro studies, HSCs may protect hepatocytes from dying while entrapped in the sinusoidal bed or promote adhesion to the endothelial wall. HSCs are also known to produce vasoactive peptide that may increase endothelial permeability and facilitate crossing of the hepatocytes (3). Those mechanisms may contribute to allow a greater number of hepatocytes to cross the vascular barrier. Additionally, as suggested by Qian's team in an islet transplantation model (5), HSCs could have provided a local immune barrier, protecting transplanted hepatocytes from sinusoidal clearance. As we used immunodeficient mice as recipients, immune rejection is most likely not at play. Nevertheless, this immunoprotective capacity of HSCs toward transplanted hepatocytes warrants further investigation.
Published strategies to improve hepatocyte engraftment used intraportal infusion of extracellular matrix components, for example, collagen and fibronectin (14), which also induced aggregates of transplanted cells in portal vein radicles. It is thus conceivable that matrix produced by HSCs during transplantation may play a similar role. To further evaluate this possibility, it will be interesting to analyze in depth whether HSCs activated to produce matrix and matrix-remodeling factors confer an additional benefit compared to quiescent cells.
We readily observed transplanted HSCs in host livers transiently after transplantation. These were cleared and no longer observed after 1 week. Although being the main effector cells in liver fibrosis, transplanted HSCs did not generate fibrosis in recipient healthy livers. Thus, and importantly, there was no proliferation of transplanted HSCs, activation of profibrogenic process, or accumulation of extracellular matrix in recipient livers. It appears that exogenous HSCs may not receive appropriate signals for their survival when they are transplanted into a healthy liver or are eliminated by apoptosis (28). A similar behavior of HSCs is expected in a context of cell therapy for single enzyme deficiency without liver injury. Obviously, further investigations are needed to determine the fate of exogenous HSCs when they are transplanted into a diseased fibrotic liver, and whether, when in a profibrotic environment, they would contribute to fibrosis progression.
In conclusion, significantly improved hepatocyte engraftment was attained upon transplantation of a mixture of cryopreserved hepatocytes and HSCs compared to transplantation of hepatocytes alone. This represents a promising strategy for liver cell therapy for the treatment of inborn error of metabolism using cryopreserved hepatocytes.
Footnotes
Acknowledgments
This research was performed as part of the BruStem projects 1 and 2, financed by Prospective Research program for Brussels (InnovIRIS), and as part of “Interuniversity Attraction Poles” (IAP) program (network P6/36 and 7/83) financed by the Belgian State-Belgian Federal Science Policy Office (BELSPO). A.-C.D. received a bursary from the “Patrimoine” of the Université catholique de Louvain. M.N. is a research associate at IREC. I.A.L. is a research associate with the “Fonds National de la Recherche Scientifique” (FNRS). We thank Dr. T. Tondreau, Ms. C. Vanhoorne, and Ms. N. Jazouli (Hepatocyte and Hepatic Stem Cells Bank, Cliniques Universitaires Saint Luc, Brussels, Belgium) for providing cryopreserved hepatocytes; and A. El Taghdouini (Liver Cell Biology Lab, Vrije Universiteit Brussel, Belgium) for preparing and providing human primary HSCs. We thank Ms. Marie-Pierre Berghmans for her technical assistance. The authors declare no conflicts of interest.