
Abstract
Sertoli cells (SCs), which are recognized as the “nurse cells” of the testis due to their important biofunctions, have been used in cotransplantation with neurons in cell therapy. However, it is not clear whether SCs influence neuronal communication and survival. In this study, we showed that approximately 60% of cortical neural stem cells (NSCs) cocultured with SCs differentiated into mature neurons. In addition, the neurite outgrowth and neuronal survival rates were significantly enhanced in the coculture system compared with differentiated neurons induced by a differentiation medium. The neuroleukin (NLK) secretion of SCs was also identified at the RNA and protein level, and the roles of NLK in neuromorphology and physiological regulation were systematically investigated for the first time. These results not only highlight the significance of paracrine regulation of NSCs by SCs but also confirm the role NLK plays in the differentiation and survival of NSCs. Finally, we proposed a possible hypothesis for the mechanism of NLK in the growth and survival of SC-induced neurons based on Western blotting results, which is that NLK secreted by SCs activates the Ras/Raf/MEK/Erk, Jak/Stat, and PI3K/Akt pathways, but not the NF-κB pathway, in neurons resulting in their growth and survival.
Keywords
Introduction
Embryonic neural stem cells (NSCs), characterized by self-renewal, potential capability of differentiating into multiple cell types, low immunogenicity, and migratory function, originate from the cortex and can propagate into neural tissues with appropriate stimuli. NSCs can differentiate into three types of cells, neurons, astrocytes, and oligodendrocytes, which positively express β-tubulin III (17), glial fibrillary acidic protein (GFAP) (29), and galactosylceramidase (Gal-C) (40), respectively. These unique qualities render NSCs useful in transplantation therapy to radically repair and replace degenerative neurons, bridge severely injured spinal cords, and help reconstruct neurotransduction pathways (2). It has been reported that human fetal spinal NSCs transplanted into the lumbar of G93A mutated superoxide dismutase (SOD1-G93A) rats can not only widely infiltrate into host neurons but also secret glialderived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF) to facilitate communication between host neurons (41). The transplantation of Lewis X [LeX or stage-specific embryronic antigen 1 (SSEA-1)] and chemokine C-X-C motif receptor 4 (CXCR4)-positive NSCs into SOD1-G93A transgenic mice has been shown to significantly prevent disease progression and extend the mouse survival time by 23 days (7).
The pursuit of cures for neurodegenerative diseases and nerve injury is hampered by many problems due to the limited capacity for repair and failure of the neurons to regenerate. Studies have shown that the main causes of neurodegenerative diseases include the aggregation of proteins (28), intrinsic and irreversible lack of repair abilities, presence of an inhibitory cellular environment (30), or severely disrupted axonal transport and communication (11). Although injured mammalian peripheral neurons can successfully undergo regeneration, neurogenesis in the central nervous system (CNS) is influenced in response to environment and inhibited by inflammation (12). Transplantation of neurons from human fetal brains as a therapy for neurodegenerative diseases is fairly effective, but the treatment of Parkinson's disease (PD) for one patient requires dopaminergic neurons from 6 to 10 human fetuses (19), making such cell therapy not conventionally available. Chemotherapy for neurodegenerative diseases is not satisfying and also causes side effects, such as lesion of postsynaptic receptors, during long-term administration (8).
It has been reported that Sertoli cells (SCs) can promote the survival and growth of cells cocultured and cografted, such as NSCs, due to many trophic factors secreted by SCs. In the testis, SCs, the somatic cells of the seminiferous epithelium, play an active role in blood–testis barrier comprised of anatomical, physiological, and immunological component. The anatomical barrier is formed through SC–SC tight junctions and restricts passage of molecules and cells from entering or exiting the lumen. The physiological barrier is constructed by transporters, which regulates movement of substances in or out of the lumen. The immunological barrier sequesters the majority of the autoantigenic germ cells (24). SCs have the potential capability of downregulating the local immune response by inhibiting the proliferation of cluster of differentiation 8-positive T (CD8+T) cells in SC–lymphocyte cocultures (9). In addition, Fas ligand (FasL) secreted from SCs regulates the apoptosis of infiltrated immunocytes by activating the FasL/Fas-induced apoptotic pathway (14). It has been hypothesized that SCs produce growth factors and neurotrophins that promote the neuronal differentiation of NSCs, axonal outgrowth, and neuronal survival (31). Cotransplantation of SCs with NSCs into the brain enhances brain development and promotes the differentiation of cografted tissues. SCs not only promote the signal transduction among the differentiated neurons but also enhance the effectiveness of the transplantation (26).
In addition, Sheng et al. (32) demonstrated that mesoderm-derived SCs can be directly converted into a multipotent state and exhibit neural stem/progenitor cell properties. These advantageous characteristics of SCs provide a promising approach for stem cell transplantation therapies.
In previous studies, we reported that expanded SCs could effectively stimulate outgrowth of NSCs in a coculture system (33,34). In this study, we showed that SCs promoted the survival and neurite outgrowth of neurons without direct contact. More importantly, we investigated a novel trophic factor secreted by SCs, neuroleukin (NLK), which also plays vital roles in promotion of the neuronal differentiation in addition to GDNF (42) and sulfated glycoproteins 1 (prosaposin) (23). For the first time, we identified the biochemical functions of NLK produced by SCs in terms of stimulating neuron outgrowth and promoting neuronal survival. The signaling pathways of embryonic cortical NSCs, which were activated by NLK, were also investigated. Our findings showed that NLK secreted by SCs plays an important role in activation of the v-akt murine thymoma viral oncogene homolog 1 (AKT), mitogen-activated protein (MAP) kinase, and Janus kinase/signal transducer and activator of transcription (Jak/Stat) pathways and promotion of the survival and outgrowth of induced neurons.
Materials and Methods
Isolation and Maintenance of Primary Cortical NSCs
Cortical NSCs were isolated from the frontal lobes of fetal cerebral cortices obtained from Imprinting Control Region (ICR) mouse embryos 14 days postcoitus (dpc). Pregnant ICR mice were purchased from the Chinese Academy of Science in Shanghai, China, and maintained in accordance with the NIH guidelines for the care and use of laboratory animals and institutional approval for animal studies. The serum-free proliferation medium consisted of Dulbecco's modified Eagle's medium/Ham's F-12 (DMEM/F12; Gibco, Grand Island, NY, USA) supplemented with 2 mM l-glutamine, 1% B27, 1% N2 supplements (Gibco), 20 ng/ml basic fibroblast growth factor (bFGF; Invitrogen, Carlsbad, CA, USA), 20 ng/ml epidermal growth factor (EGF; Invitrogen) (3,26,27), 100 units/ml penicillin, and 100 units/ml streptomycin (Gibco). The cell suspension was incubated at the concentration of 1.5 × 105 cells/ml in a 50-ml flask (Corning, Shanghai, China) at 37°C in 5% CO2–95% air for 5 days. The cellular morphology was observed each day. For subculturing, monoclonal neural spheres were digested with 0.25% trypsin in phosphate-buffered saline (PBS) with 0.05% EDTA and phenol red (all Biowest, Nuaillé, France). DMEM/Ham's F-12 medium (1:1) supplemented with 10% fetal calf serum (FCS; Gibco) was used to stop the digestion.
SC Culture and Identification and Preparation of Conditioned Medium
The SCs were isolated from ten 15-day-old ICR male mice as previously described (5). The white testicular films were removed from the testis tissues. The testicular tubules were then cut into 1-mm3 pieces and digested with a mixture of 0.25% trypsin with 0.05% EDTA and 0.1% collagenase (1:1; Invitrogen) in a water bath and shaken at 60 cycles per minute at 31°C in 5% CO2–95% air for 30 min. Thereafter, the tubule pieces were washed two times with DMEM/F12 supplemented with 10% FCS in order to cease the digestion and then centrifuged for 5 min at 900 × g. The individual cell was separated from the tissues by gently blowing with pipet and filtered using a cell sieve for purification. The cells were resuspended at 5 × 105 cells/ml and cultured in DMEM/F12 with 10% FCS at 37°C in 5% CO2–95% air for 4 days. The SC-enriched monocultures contained more than 97% SCs, as described previously (19). The adherent cell colonies at the bottom of culture flask were washed, gently trypsinized, and centrifuged twice at 900 × g for 5 min in DMEM/F12 with 10% FCS. The SCs were identified by hematoxylin and eosin (H&E; Sigma, St. Louis, MO, USA) staining and labeled with an anti-FasL antibody (Boshide Technology, Wuhan, China) (34).
To prepare conditioned medium, the near-confluence SCs in passage 3 were washed three times with sterile PBS to remove the residual FCS. And then SCs were further cultured for 3 days in serum-free medium. The conditioned medium was collected and centrifuged at 5,600 × g for 10 min. The supernatant was immediately used to induce the differentiation of cortical NSCs.
Cell Viability Assays
After isolation and expansion, cortical NSCs attained a morphological purity of 96 ± 2.1%, as observed by trypan blue (Sigma) exclusion tests that quantitatively assessed cells viability under a light microscope.
Preparation of SC Feeder Layer
SCs isolated from ten 15-day-old ICR male mice were plated at a cell density of 2,500 cells/cm2, and the culture medium was replaced by DMEM/Ham's F-12 medium (1:1) every 3 days. At near confluence, these cells were subcultured until passage 3. The SCs at passage 3 were seeded at a cell density of 2,500 cells/cm2 as a feeder layer using Chinese hamster ovary (CHO) cells and human embryonic kidney (HEK293) cells (both from ATCC, Manassas, VA, USA) as negative controls (34).
Transwell Filter Assay
Transwell permeable supports were used to determine whether SC-induced neural differentiation was due to products secreted by SCs or required the direct contact between the two types of cells. Cortical NSCs were plated in six-well plates (Corning). Filter membrane inserts (with 0.4-μm filter membrane and 24-mm insert diameter; Corning) were laid over the NSCs, and SCs were cultured on the filter membrane. These inserts provided separation and independent access to both the SCs and cortical NSCs.
Immunocytochemistry Fluorescence Staining
To detect specific proteins in the cells, cortical NSCs were plated at the concentration of 5 × 104 cells/ml on poly-l-lysine-coated six-well plates (Corning) at 37°C in 5% CO2–95% air and serum-free proliferation medium. Filter membrane inserts, on which SCs were plated, were laid over the cortical NSCs the next day. At 72 h after the induction, the differentiated cells were fixed with 4% paraformaldehyde (Sigma) at 4°C for 30 min, washed three times with 0.01 M PBS for 5 min, permeabilized with 1% PBST (PBS with 1% Triton X-100; Millipore, Billerica, MA, USA) for 30 min, washed three times with 0.01 M PBS again for 5 min, and blocked with 5% goat serum (Millipore) in 0.3% PBST for 30 min. The cells were incubated with a mouse monoclonal anti-β-tubulin III antibody (TUJ clone; 1:2,000 dilution; Sigma) or rabbit anti-GFAP antibody (1:300; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 2 h at room temperature and washed three times with 0.01 M PBS for 5 min. The cells were then incubated with fluorescence-tagged secondary antibodies, goat anti-mouse IgG (1:200 dilution; Santa Cruz Biotechnology), or goat anti-rabbit IgG (1:200 dilution; Jackson Immuno-Research Laboratories, Inc., West Grove, PA, USA) for 2 h at room temperature in a dark moist box. After the incubation, the cells were washed with 0.01 M PBS three times for 5 min each and nuclear stained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma) at 1 μg/ml in the darkness.
The specificities of anti-β-tubulin III and anti-GFAP antibodies were respectively detected using the male rat pheochromocytoma (PC12; ATCC) cell line and the male human astrocytoma cell line, U251 (CLS Cell Lines Service, Eppelheim, Germany). The randomly collected colonies were immunostained at different stages of differentiation, and nuclear staining was also performed at the same time to determine the total number of cells and the percentage of positively stained cells (38). The fluorescent immunostaining of NSCs with anti-Nestin antibody (1:3,000 dilution; Sigma) was performed in the same way as described above.
Measurement of Neurite Outgrowth
Images of β-tubulin III-positive differentiated neurons were obtained using fluorescence microscopy (Olympus, Tokyo, Japan). The length of a neurite was defined as the distance from the cyton to the end of the longest axon branch. Photos of the cells were taken from randomly selected areas in at least three independent experiments. Morphological characteristics were evaluated using SigmaScan Pro.5.0 (SPSS, Chicago, IL, USA).
RNA Isolation and RT-PCR Analysis
Total cellular mRNA was isolated using TRIzol reagent (Invitrogen) according to manufacturer's instructions. The purity of RNA was checked using a UV spectrophotometer (SmartSpec plus, Biorad, Hercules, CA, USA). RNA (5 μg per sample) was reverse-transcribed into cDNA with the PrimeScript II 1st Strand cDNA Synthesis Kit (Takara Shuzo, Kyoto, Japan) and prepared for RT-PCR analysis. The cDNA was mixed with each primer (listed in Table 1) and Taq DNA polymerase (Takara Shuzo) and then amplified according to the manufacturer's instruction. The PCR reaction products were analyzed by electrophoresis in a 1.0% agarose gel (Invitrogen).
Genes and Primers Used in This Study

Nlk: neuroleukin; gdnf: glial derived neurotrophic factor; il-1βinterleukin 1 beta; myox: myosin X; lrrc4: leucine rich repeat containing 4; map2: microtubule-associated protein 2.
Anti-NLK Antibody Blockade and Exogenous NLK Supplementation
Primary cortical NSCs were cultured in three different conditions to investigate the effects of NLK on their differentiation: culture medium with NLK blocking antibodies (final concentration 6 μg/ml, Santa Cruz Biotechnology), SC feeder layers and SC feeder layers plus NLK (final concentration 12 μg/ml, Sigma Aldrich) for 7 days in a CO2 incubator at 37°C. Medium was replaced on the third day of coculture. The length and density of axons were recorded every other day.
Sodium Dodecyl Sulfate Polyacrylamide Gel Electropheresis (SDS-PAGE) and Ultra Performance Liquid Chromatography Electron Spray Ionization Tandem Mass Spectrum (UPLC-ESI- MS/MS) Analyses
The serum-free culture supernatant of SCs was concentrated to 2.13 mg/ml with ultrafiltration devices with a 3-kDa filter membrane (Millipore) at 4°C and spun at 5,600 × g for 30 min. Proteins in the samples were separated by 10% SDS-PAGE (Sigma) and analyzed by Coomassie Brilliant Blue 250 staining (Sigma). Proteins in the gels were excised and analyzed by UPLC ESI MS/MS (nano-ACQUITY UPLC, Waters, Milford, MA, USA).
The ultra performance liquid chromatography binding ethane with bariegated pellet column was BEH C18 with 75 μm × 250 mm specifications and 1.7 μm pellets (Waters, Kitashinagawa, Tokyo, Japan). The column temperature was 35°C. The enrichment column was Symmetry C18 with 180 μm × 20 mm specifications (Waters). The mobile phase A consisted of water solution with 0.1% formic acid, and the mobile phase B was acetonitrile solution with 0.1% formic acid (Sigma). The detached segments were assessed by Synapt High-Definition Mass Spectrometer (Waters). The mass spectral data obtained after processing with the data analysis software PLGS version 2.3 (SAS Institute, Inc., Marlow, Buckinghamshire, UK) were used to search the NCBInr database with an MS/MS ion strategy using Mascot software (http://www.matrixscience.com). Experimental parameters included trypsin digestion, M oxidation, iodoacetamide alkylation (variable modification), and one missed cleavage. The mass error tolerance in both the MS and MS/MS data was 0.2 Da.
Western Blotting
Proteins were resolved by electrophoresis in 10% Tris-Tricine gels (Sigma), transferred onto polyvinylidenedifluoride (PVDF) membranes (GE Bioscience, Little Chalfont, Buckinghamshire, UK) and then incubated with primary antibodies against phosphorylated (p)-Akt (Ser473), p-Akt (Thr308), p-mitogen-activated protein kinase kinase 1 (MEK1; Ser221), p-MEK1 (Thr291), p-ribosomal S6 kinase 1 70 kDa splicing form (p70S6k; Ser424), p-cAMP response element-binding protein (CREB; Ser133), p-V-Raf-1 murine leukemia viral oncogene homolog 1 (Raf1; Ser338), p-B-cell CLL/lymphoma 2 [BCL2]-associated agonist of cell death (Bad; Ser136), p-Tau (Thr212), p-protein kinase D (PKD; Ser738), p-v-myc avian myelocytomatosis viral oncogene homolog (Myc; Thr358), p-nuclear factor of k light polypeptide gene enhancer in B-cell 1 (NF-κB; Ser529), p-NF-κB (Ser536), p-Jak1 (Tyr1022), or p-p70S6k (Ser133) (Nanjing Chuanbo Biotech, Nanjing, China). Immunoreactivity was detected with goat anti-mouse horseradish peroxidase (HRP)-labeled IgG (Nanjing Chuanbo Biotech). After incubation with substrate, the immunoblots were developed by exposure to film in a darkroom.
Statistical Analysis
All results are reported as mean ± SD. Analysis was performed using the SPSS statistical software (version 11.5.1). After testing for Gaussian distribution, data of two independent variables were analyzed using Student's t test for equally distributed data or the Mann–Whitney test (not normally distributed data), respectively. For three or more variables, ANOVA or ANOVA on ranks was performed followed by post hoc analysis using Tukey's test. Values of p < 0.05 were considered to be statistically significant and are indicated by single asterisks.
Results
Characterization of Cortical NSCs and SCs
The subcultured cortical NSCs, isolated from cerebral frontal lobes (Fig. 1A), proliferated and formed a suspension of monoclonal neural spheres at 37°C in 5% CO2–95% air. The size of each soma was approximately 10 ± 0.45 μm, and the cytoplasm appeared bright, round, and transparent under a light inverted microscopy. However, after cultivation for 24 h, some dark cells with low viability suffered a diminution in size and quantity when observed by microscopy and trypan blue exclusion assays. During the culture period, the flasks were not moved to allow the neural spheres to be formed by clonal cells. Viabilities of cortical NSCs that had been passaged for five times were significantly reduced.

Identification of cortical NSCs and SCs by immunofluorescence staining. Embryonic neural stem cells (NSCs) were isolated from fetal cerebral frontal lobes (A) at 14 days, and Sertoli cells (SCs) were from mouse testis at 10–15 days postcoitus (dpc). NSCs (C–E) were stained to label nestin (C) and 4′,6-diamidino-2-phenylindole (DAPI; D). The merged image shows an overwhelming majority of cells exhibiting positive nestin staining and maintaining the undifferentiated characteristics of NSCs (E). Identification of NSCs (B, F, G) in different media by immunofluorescence staining with primary antibodies against Nestin (B), β-tubulin III (F), and glial fibrillary acidic protein (GFAP; G). The unique binuclear feature of SCs was identified by hematoxylin and eosin (H&E) staining (H), and its morphology was observed by scanning electron microscopy (I). Scale bar: 30 μm (A–H) and 5 μm (I).
To validate that the cloned cells were NSCs, the sub-cultured neural spheres (Fig. 1B) were trypsinized, and then the single cells, which were plated at a concentration of 5 × 104 cells/ml, became partially adherent with one or two projections after attaching onto the bottom of the poly-l-lysine coated six-well plates. These cells positively stained for the biomarker Nestin in immunocytochemical analysis, indicating that they were NSCs. The purity of the NSCs was 93 ± 3.4% as determined by double staining for Nestin and the nucleus with DAPI (Fig. 1C–E). After culture in differentiation medium for 7 days, the cells expressed the neuronal marker β-tubulin III (Fig. 1F) and astrocyte marker GFAP (Fig. 1G), confirming that the previously undifferentiated cells acquired from mouse fetal brains were NSCs. The SCs isolated from newborn ICR mice and used in the induction of cortical NSCs were identified by observation of the unique karyosome structure following H&E staining and immunocytochemically labeled with FasL antibody (Fig. 1H and Fig. S1). Meanwhile, the ultrastructure of the SCs was also observed by electronic scanning microscopy (Fig. 1I). There are some small holes on the cell membrane of SC, which are very thick.
Induction of Neural Differentiation From Cortical NSCs on SCs
In order to optimize the cell culture condition for neural differentiation, the cortical NSCs were further characterized by culturing in different conditions, either with SC-conditioned medium (SCCM), with SCs directly, with SCs separated by a transwell filter (TFSCs), or with differentiation medium (DM) as a positive control. After 7 days of induction with SCs, the cortical NSCs were efficiently induced into postmitotic neurons (60 ± 2.1%) that were β-tubulin III positive (antibody clone TUJ+), and extensive neurites formed in the majority of the NSCs containing 35 ± 1.4% neural precursors (Fig. 2A) as confirmed by the fluorescent imaging (Fig. 2B–D). Some of the cells gradually spread out and formed relatively larger somas than others and expressed GFAP (Fig. 1G). Of note, the ratios of β-tubulin III-positive cells in the TFSCs and SCCM induction groups were 48.5 ± 2.1% (Fig. 2E–G) and 38 ± 1.5% (p < 0.05) (Fig. 2H), respectively, which were significantly lower than that in the SC induction group (p < 0.05), but still much higher than the 13 ± 0.5% in the DM induction group (p < 0.05) (Fig. 2K–M). The results indicate that paracrine regulation by SCs is involved in the neural differentiation, and direct cell contact between SCs and NSCs can enhance the differentiation of primate cortex NSCs. Therefore, the direct contact coculture style (NSCs or neurons cocultured on SC feeders) was chosen in the following experiments.

Induction of cortical NSC differentiation into β-tubulin III-positive neurons. The levels of β-tubulin III (TUJ antibody clone) staining were quantified (A). Cortical NSCs were cultured with SCs (B–D), with SCs separated by a transwell filter (SCTF; E–G), SC conditioned medium (SCCM; H–J), or differentiation medium (DM; K–M). The cells were stained with anti-β-tubulin III antibody (B, E, H, K) and DAPI (C, F, I, L). Merged images (D, G, J, M) show differences in differentiation between the four conditions. Scale bar: 50 μm. Differentiation medium mainly contained Dulbecco's modified Eagle's medium/Hams F12 (DMEM/F12; 1:1) and 2% fetal calf serum (FCS). Significant differences: *p < 0.05; n = 3.
Additionally, CHO cells and HEK293 cells were also used to induce neural differentiation of primate cortex NSCs under serum-free conditions as control, but neither significantly induced the neural marker β-tubulin III after a week in culture (data not shown). These findings demonstrate that promotion of neuronal differentiation by SCs may be specific feature of SCs.
Role of NLK in Promotion of Axonal Outgrowth and Neural Survival
When the differentiated neurons were further cultured in a 24-well plate in the absence of SCs, the axon density and length were lower or shorter compared with those cultured with SCs in the same medium (Fig. 2). This result suggests that SCs also play a prominent role in axonal outgrowth and neural survival after NSC differentiation.
NLK is a potential mediator of the neurite outgrowth and protrusions along axons (13,16), but not as a stimulus for priming neural differentiation. In our study, the presence of transcripted NLK gene in SCs was confirmed by total RNA extract and reverse transcription-PCR (Fig. 3A). To further verify and characterize the function of NLK, since the NLK primers also recognize glucose-6-phosphate isomerase (GPI), we systematically analyzed the effects of exogenously applied NLK and the anti-NLK antibody on neurite outgrowth, promotion of protrusions, and neural survival by culturing the primary cortical NSCs in different conditions: SCs + anti-NLK (negative control), SCs (positive control), and SCs + NLK (Fig. 3B–I). At the beginning of the experiment, the average axonal lengths in the different culture conditions were similar (Fig. 3C). However, after 7 days, the average axonal length of neurons in coculture with SCs plus NLK reached 176.4 ± 6.1 μm (Fig. 3C, F), which was significantly higher than those of the SC/anti-NLK mAB group (110 ± 4.2 μm) (Fig. 3C, D) and SC group (145.2 ± 6.9 μm) (p < 0.05) (Fig. 3C, E). The findings indicated that NLK could remarkably enhance the growth of axons in length compared with the other groups (p < 0.05) (Fig. 3C–F). Interestingly, the proliferation medium with NLK did not elicit neural differentiation.

The effect of NLK on SC-induced neurite outgrowth and synapse formation. Neurotrophic factors were detected by RT-PCR analysis of RNA isolated from SCs. The expression levels of the effective trophic factors GDNF, NLK, IL-1β, and other factors including MyoX, MAP2, LRRC4, and drebrin, acting as stimuli in neurite outgrowth, are shown (A). An increasing trend of the number of axonal protrusions/neuron in the bar chart illustrates the pivotal role of NLK in enhancing the density of axonal protrusions (B). The bar charts (C) show that the average axonal length of neurons under different culture conditions for 7 days is greater when cultured under the condition of Sertoli/NLK compared to the other two conditions (p < 0.05). While the SC feeders gradually diminished by 7 days in the serum-free medium, the exogenous NLK continued to be effective in neurite outgrowth. β-Tubulin III-positive immunofluorescence images of representative neurons cultured on SC feeders treated with an anti-NLK antibody (D, G), a SC feeder layer alone (E, H), and SCs with exogenous NLK (F, I) are shown. White linear profiles (D–F) indicate the extension of each axon influenced by SCs and NLK. Scale bar: 30 μm (D–I). Abbreviations: NLK, neuroleukin; GDNF, glial cell-derived neurotrophic factor; IL-1β, interleukin-1β; MyoX, myoglobulin X; MAP2, microtubule-associated protein 2; LRRC4, leucine-rich repeat containing-4. Significant differences: *p < 0.05; n = 3.
The average number of protrusions along an axon was slightly enhanced by NLK in these experiments and increased up to 11.8 ± 0.4 (Fig. 3B, G, H, I), which was more than that on axons blocked by an anti-NLK antibody at day 7 (p < 0.05). This result indicated that NLK could effectively stimulate axonal outgrowth (p < 0.05).
Furthermore, after incubation for 7 days, trypan blue exclusion tests showed that the survival rate of neurons fell to 52 ± 2.2% in the culture with DM, which was significantly lower than that (66 ± 3.1%) induced by DM with added NLK (p < 0.05) (Fig. 4). There were also remarkable differences on the survival rates of neurons between conditioned medium group (62± 2.1% without additional NLK vs. 83 ± 4.2% with additional NLK) and SC group (82 ± 4.2% without additional NLK vs. 95 ± 4.8% with additional NLK) (p < 0.05) (Fig. 4). These results suggest that NLK is an important component of the neurotrophic substances secreted by SCs.

Survival rates of cortical NSCs cultured in DM, SCCM, or on SC feeders. The survival rate was evaluated by trypan blue staining after culture for 7 days. Significant differences: *p < 0.05; n = 3.
SDS-PAGE and UPLC-ESI-MS/MS Analyses of SC Secreted Protein
To identity the functional proteins secreted by SCs, the serum-free culture supernatant of SCs was concentrated and analyzed by SDS-PAGE gel and UPLC-ESI-MS/MS. As shown in Figure 5, most of the proteins were located between the molecular weight (MW) of 55 and 70 kDa, and some of them were around 56 kDa. It was reported that the MW of NLK is about 56 kDa (37); therefore, the result of SDS-PAGE gel indicates that serum-free culture supernatant of SCs may contain NLK. In addition, the UPLC-ESI-MS/MS analyses demonstrate that 7% sequence coverage matches with the peptide sequences of NLK, which provides further support for the existence of NLK in the serum-free culture supernatant of SCs.

Detection of NLK in the SC culture supernatant by SDS-PAGE analysis of histones and UPLC-ESI-MS/MS. The 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Coomassie Brilliant Blue 250 staining were performed to separate protein of various masses from serum-free supernatant of SCs cultured in DM, and the bands were excised from the gel for protein identification (A). Peptides analyzed by Ultra Performance Liquid Chromatography Electron Spray Ionization Tandem Mass Spectrum (UPLC-ESI-MS/MS). NLK [mus] (gi 200065) was identified by the individual ion score of 59 (>37) (p < 0.05) (B). 7% sequence coverage (TLASLSPETSLFIIASK, HFVALSTNTAK, and VFEGNRPTNSIVFTK) matches the peptide sequences of NLK. M: Marker (or KD ladder).
NLK Treatment Activates Akt, MEK1, CREB, Jak1, and p70S6k
Neurons were incubated for 3 days in serum-free media with or without NLK stimulation. To investigate whether antiapoptotic and cell growth-related signaling pathways were involved in neural outgrowth and survival induced by NLK, activities of the following 15 phosphorylated kinases were examined by Western blotting analysis: p-Akt (Ser473), p-Akt (Thr308), p-MEK1 (Ser221), p-MEK1 (Thr291), p-p70S6k (Ser424), p-CREB (Ser133), p-Raf1 (Ser338), p-Bad (Ser136), p-Tau (Thr212), p-PKD (Ser738), p-Myc (Thr358), p-NF-κB (Ser529), p-NF-κB (Ser536), p-Jak1 (Tyr1022), and p-p70S6k (Ser133). As shown in Figure 6, the levels of p-Akt (Ser473), p-MEK1 (Ser221), p-MEK1 (Thr291), p-Raf1 (Ser338), p-Akt (Thr308), p-CREB (Ser133), p-Jak1 (Tyr1022), and p-p70S6k (Ser133) were significantly increased in NLK-stimulated neurons, while the expression levels of p-p70S6k (Ser424), p-Bad (Ser136), p-Tau (Thr212), p-PKD (Ser738), p-NF-κB (Ser529), p-Myc (Thr358), and p-NF-κB (Ser536) did not show any significant difference under various culture conditions (data not shown). The results indicated that NLK activated the Akt-S6K, Raf/MEK/extracellular signal-regulated kinase (Erk), and Jak/Stat pathways, but not the NF-κB and Myc pathways.
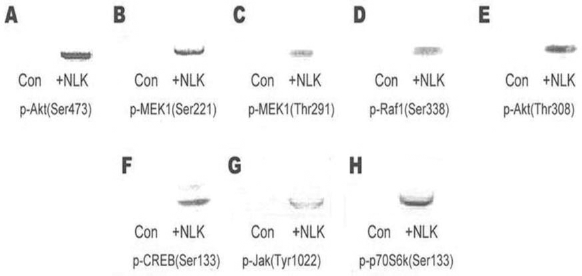
Detection of phosphorylated signaling proteins by Western blot in NSCs cultured with NLK and the control on day 3. (A) p-v-akt murine thymoma viral oncogene homolog 1 (Akt; Ser473); (B) p-MEK1 (Ser221); (C) p-mitogen-activated protein kinase kinase 1 (MEK1; Thr291); (D) p-V-Raf-1 murine leukemia viral oncogene homolog 1 (Raf1; Ser338); (E) p-Akt (Thr308); (F) p-cAMP response element-binding protein (CREB; Ser133); (G) p-Janus kinase (Jak; Tyr1022); (H) p-ribosomal S6 kinase 1 70 kDa splicing form (p70S6k; Ser133).
Discussion
In the SC–NSC coculture system, axonal outgrowth can be stimulated and the survival of the induced neurons promoted by SCs. However, the mechanisms underlying these processes are unclear. In the present work, our results revealed that the SCs still played an effective nursing role even without direct contact and that the NLK secreted by the SCs had a prosurvival function in protecting the differentiating neurons.
In addition to coculturing with SCs, NSCs can be induced to differentiate into neurons in other ways, such as by transferring the neuron-specific gene sex-determining region Y box 2 (Sox2) into NSCs or adding exogenous BDNF, neurotrophin-3 (NT-3), or GDNF. Undeniably, the coculture system has the advantages of simplicity and high efficiency in contrast to previous protocols (37) and does not require growth serum, retinoic acid, exogenous cytokines, or other special treatment.
In previous study, we reported that SCs can enhance the growth and survival of NSCs and neurons (33,34). In this study, a transwell coculture system was employed in order to determine the mechanism: whether the promotion of neurite outgrowth and prosurvival were mediated only by putative secreted factors or regulated by cell surface-anchored factors. We used filter membrane inserts to identify whether direct physical contact between the cortical NSCs and SCs was critical for the differentiation and prosurvival effect. By comparison, cortical NSCs were plated at the density of 1 × 105 cells/ml directly on the SC feeder layer. As shown in Figures 3 and 4, NSCs were still induced even when physically separated from the SCs in coculture as expected, albeit at a relatively lower rate with a shorter average axonal length, indicating that direct contact does not play a vital role in the induction during cocultivation. The results indicated that one or more bioactive molecule(s) produced by SCs may have been critical for promoting differentiation of NSCs and enhancement of their survival.
It has been reported that NLK, a multifunctional protein of relative molecular mass (Mr) 56 kDa, was found in skeletal muscle, brain, heart, and kidneys, which acts as neurotrophic factor promoting growth of embryonic spinal and sensory neurons (15). Glucose-6-phosphate isomerase (GPI) is also able to support the survival of cultured sensory and specific embryonic spinal neurons, which is considered as a NLK secreted by lectin-stimulated T-cells (6). Interestingly, the dimeric form of NLK performs the isomerase activity, whereas the monomeric form of the protein carries out its neurotrophic activity. In the presence of monomeric GPI, neuroblastoma cells have enhanced neurite extension and a reduced proliferation rate (22). In most case, GPI has been found to be identical to NLK (6,13) with the PCR primers recognizing both genes in this study.
In this study, we discovered a new SC-secreted factor, NLK, which exerted prosurvival effects on the differentiating neurons. Many different active proteins produced from SCs have been reported, such as a cocktail of neural trophic factors, including GDNF and interleukin (IL)-1β, and the regulatory proteins associated with axonal outgrowth, such as microtubule-associated protein 2 (MAP2), leucine rich repeat containing 4 (LRRC4), drebrin (18), and myosin X (MyoX). First, we have confirmed the presence of NLK/GPI at the transcription level using RT-PCR as shown in Figure 3A and their existence at the protein level using UPLC-IES-MS/MS analysis for the first time (Fig. 5). The existence of NLK secreted by SCs was further identified due to the high similarities between NLK and GPI. It is known that the monoclonal anti-NLK antibody can react in an enzyme-linked immunosorbent assay (ELISA) with GPI as well, but cannot inhibit the enzymatic activity of GPI (13). Our results shows that the average axonal length of neuron significantly decreased after adding the monoclonal anti-NLK antibody in the SC–NSC coculture system (Fig. 3C), which demonstrates that NLK was blocked in the SC–NSC coculture system and also reveals that SCs secrete NLK. To investigate the prosurvival function of NLK secreted from SCs, a moderate amount of exogenous NLK was supplied to the SC–NSC coculture system. We found that the exogenous NLK could promote the survival and outgrowth of neurons. Moreover, even if the viability of SCs was reduced in serum-free media on day 7, the exogenous NLK could significantly and consistently prevent the reduction in survival of SC-induced neurons. This result was similar to that of a previous study demonstrating that NLK plays an antiapoptotic role in neurons, and those cells with inhibited NLK expression would be more sensitive to apoptosis compared with noninduced cells (27). In addition, we found that the average axonal length of induced neurons cocultured with SCs was longer than that of cells cultured only with NLK in basic media, DMEM/F12. This result uncovers the fact that multiple factors function synergistically in neurite outgrowth and promotion survival.
To date, how NLK promotes the survival of neurons has not yet been reported. In this work, we have systematically investigated the NLK signal pathways involved in induction of neuron growth and survival. Based on results of the protein analysis in Figure 6, we hypothesized that NLK signal transduction occurs as follows during NSC differentiation. First, activation of the NLK monomer is mediated by its binding to a specific receptor, glycoprotein 78 (gp78) or autocrine motility factor receptor (AMFR), which belongs to the G protein-coupled receptor family (4). The Akt phosphorylation-oriented pathway and perhaps the Ras/Raf/MEK/Erk and Jak/Stat pathways are then activated to participate in the prosurvival effects on the neural cells based on the positive detection of p-Akt (Ser473), p-MEK1 (Ser221), p-MEK1 (Thr291), p-CREB (Ser133), p-Jak1 (Tyr1022), and p-p70S6k (Ser133) (Fig. 6). Akt is classically described as a prosurvival serine/threonine kinase activated in response to trophic factors. After activation by phosphoinositide 3-kinase (PI3K), it can translocate into the nucleus where it can catalyze phosphorylation of transcription factors (35). It has been reported that, upon phosphorylation of Ser473 in the C-terminal regulatory domain of Akt, it can transmit survival signals to many cell types by regulating the expression of antiapoptosis molecules such as Bcl-2 and inhibitor of apoptosis protein (cIAP)-1/2 or by modifying the activity of apoptosis molecules (36). Furthermore, Tsuruta et al. reported that Akt also inhibits conformational change in the proapoptosis Bax protein and its translocation to mitochondria, which effectively prevents the activation and apoptosis of caspase 3 (39). It has also been reported that PI3K/Akt is able to inhibit the Raf/MEK1/Erk1/2 pathway (21,25). However, according to the results of our study, following NLK treatment, both the MEK1 and Akt pathways are activated independently and probably play roles in cellular survival, which is consistent with the results of Abkhezr and colleagues (1).
Regarding involvement of the Raf/MEK1/Erk1/2 transduction in promoting neuronal survival, it is widely accepted that this pathway can phosphorylate and inhibit the degradation of XIAP, the X-linked inhibitor of apoptosis, which inhibits such critical apoptosis regulators as caspase 3 and caspase 9 (1,10). We confirmed that NLK treatment also resulted in the phosphorylation of p70S6k at Ser133. Lin et al. (20) had demonstrated that the p70S6k pathway, a downstream signaling molecule in the PI3K/Akt pathway, may also play an antiapoptotic role. The phosphorylation of Jak1 kinase allows the transcriptional factors Stat1 and 3 to translocate and bind to active CREB, and the Stat1, 3 and CREB transcriptional complex stimulates the transcription of Bcl-2 and B-cell lymphoma-extra large (Bcl-xL), which have prosurvival effects in cells. Our proposed regulatory pathways of the NLK secreted by SCs as described above are presented diagramatically in Figure 7.

Putative regulatory pathways of NLK secreted by SCs. NLK can enhance the growth of neurites and axonal synapses and the survival of neurons. The possible signal transduction pathways involved in these processes are proposed as follows: NLK with its isomerase activity can promote the formation of glycogen when it exists as a dimer in SCs. However, NLK is depolymerized into monomers and exerts neurotrophic effects when secreted out of the cell. The monomeric NLK can bind its specific receptor glycoprotein 78 (gp78; also known as AMFR, autocrine motility factorreceptor), activating three signal transduction pathways: (1) The phosphatidylinositol 3-kinase (PI3K)/Akt pathway can be activated by the NLK/gp78 complex based on the simultaneous double phosphorylation at the Akt (Ser473) and (Thr308) sites. PI3K is first activated, followed by p70s6k and Akt activation. Both p70s6k and Akt can inhibit the phosphorylation of B-cell CLL/lymphoma 2 (BCL2)-associated agonist of cell death (Bad), resulting in inhibition of the mitochondrial apoptotic pathway. Akt is able to inhibit cellular apoptosis by prohibiting the production of caspase 9, p53, and Bax directly or indirectly. PI3K can also prohibit the secretion of poly ADP ribose polymerase (PARP) to inhibit cellular apoptosis. (2) The Raf/MEK/extracellular signal-regulated kinase (Erk) pathway can be activated based on the phosphorylation of Raf (Ser338), MEK1 (Ser221), and Erk (Thr291), and the degradation of X-linked inhibitor of apoptosis protein (XIAP) can be further inhibited by its phosphorylation. In that case, activities of the apoptotic factors, caspase 3 and caspase 9, are inhibited, which can downregulate the apoptosis pathway of cellular signal transduction. (3) NLK/gp78 complexes can promote the phosphorylation of the kinase Jak1, which can enhance the nucleus transport and formation of complexes of signal transducer and activator of transcription 1 (Stat1), Stat3, and CREB. The complexes can bind to the promoters and activate transcription of the genes for Bcl2 and Bcl-XL, resulting in highlevel production of the proteins and ultimately inhibition of cellular apoptosis. Abbreviations: MDM2, mouse double minute 2; PDK1, phosphoinositide-dependent kinase-1; PGI, phosphoglucose isomerase.
Conclusions
The nursing role of SCs was confirmed to effectively function without direct contact with NSCs in this study. Furthermore, NLK was identified for the first time as one of critical active factors for neuronal survival and axonal outgrowth in SC–NSC coculture system. A hypothesis for the underlying mechanism of NLK was proposed in which binding of monomeric NLK to a specific receptor, gp78 or AMFR, activates the Ras/Raf/MEK/Erk, Jak/Stat, and PI3K/Akt pathways, but without triggering activation of NF-κB.
Footnotes
Acknowledgments
We thank Tao Fang (Chinese Academy of Military Medical Science) for technical supports. This work was supported by grants from the National Basic Research Program of China (2013CB733605), the National High Technology Research and Development Program (863 Program) of China (2012AA021201 and 2012AA02A401), the Shanghai Municipal Government (11DZ2292700), and the Fundamental Research Funds for the Central Universities of ECUST. The authors declare no conflicts of interest.