
Abstract
Insulin-like growth factor II (IGF2) is a growth-promoting peptide that increases β-cell proliferation and survival. The aim of the study was to determine the effect of IGF2 overexpression on β-cell mass in transplanted islets. Islets infected with adenovirus encoding for IGF2 (Ad-IGF2 group), for luciferase (Ad-Luc control group), or with uninfected islets (control group) were syngeneically transplanted to streptozotocin-diabetic Lewis rats. Eight hundred islets, a minimal mass model to restore normoglycemia, or 500 islets, a clearly insufficient mass, were transplanted. Rats transplanted with 800 Ad-IGF2 islets showed a better metabolic evolution than control groups. As expected, rats transplanted with 500 Ad-IGF2 or control islets maintained similar hyperglycemia throughout the study, ensuring comparable metabolic conditions among both groups. β-Cell replication was higher in Ad-IGF2 group than in control group on days 3 [1.45% (IQR: 0.26) vs. 0.58% (IQR: 0.18), p = 0.006], 10 [1.58% (IQR: 1.40) vs. 0.90% (IQR: 0.61), p = 0.035], and 28 [1.35% (IQR: 0.35) vs. 0.64% (IQR: 0.28), p = 0.004] after transplantation. β-Cell mass was similarly reduced on day 3 after transplantation in Ad-IGF2 and control group [0.36 mg (IQR: 0.26) vs. 0.38 mg (IQR: 0.19)], it increased on day 10, and on day 28 it was higher in Ad-IGF2 than in control group [0.63 mg (IQR: 0.38) vs. 0.42 mg (IQR: 0.31), p = 0.008]. Apoptosis was similarly increased in Ad-IGF2 and control islets after transplantation. No differences in insulin secretion were found between Ad-IGF2 and uninfected control islets. In summary, IGF2 overexpression in transplanted islets increased β-cell replication, induced the regeneration of the transplanted β-cell mass, and had a beneficial effect on the metabolic outcome reducing the β-cell mass needed to achieve normoglycemia.
Keywords
Introduction
Islet transplantation restores normoglycemia in type 1 diabetic patients (44), but with time hyperglycemia recurs in most patients despite the maintenance of partial graft function (40). The cause of the progressive loss of insulin independence is not well established and is probably multifactorial. Damage of transplanted islets due to rejection, recurrence of autoimmunity, β-cell toxicity of immunosuppressive therapy, and metabolic stress may contribute to the long-term failure of islet grafts (23). The amount of transplanted islet tissue determines also the outcome of islet transplantation. A high islet mass must be transplanted to achieve normoglycemia, probably due to the massive destruction of islets taking place in the initial days after transplantation (5,8), and the number of initially transplanted islets contributes to late function or failure of the graft (22,48,51). Since the islet mass that becomes engrafted is low, minimal additional losses may determine the recurrence of hyperglycemia (23). Thus, new strategies aimed to preserve or increase the engrafted β-cell mass are needed to improve the current success rate of islet transplantation.
Insulin-like growth factors I and II (IGF1, IGF2) are potent cell growth regulators that promote proliferation and survival in β-cells (50). The effects of IGF2 have been demonstrated in β-cell lines, in rat and human β-cells, and in transgenic mice (2,13,18–20,35,37). An autocrine role of IGF2 in protecting β-cells against apoptosis was also recently proposed (7). It may be expected that these beneficial effects of IGF2 could be useful to increase β-cell mass in islet transplantation. Interestingly, an association has been recently described between insulin-like growth factor II mRNA binding protein 2 (IGF2BP2), that binds to the leader elements in the untranslated regions of IGF2 transcripts and regulates IGF2 translation during late mammalian development (32) and type 2 diabetes (41,43,54). However, some studies have suggested that IGF2 overexpression may have deleterious effects on islet cells. Transgenic mice overexpressing IGF2 show large and irregular shape in islets with disrupted islet architecture (9,35), a left shift in the dose–response curve relating insulin secretion and glucose concentration, and they develop diabetes (9). Transgenic mice expressing IGF2 in β-cells have a higher susceptibility to streptozotocin-induced β-cell damage, and double transgenic interferon (IFN)-β/IGF2 mice develop spontaneous diabetes (1). Thus, although IGF2 may have a beneficial role on β-cells, acting as a mitogenic and prosurvival factor, the transgenic data suggest that sustained overexpression of IGF2 could increase the susceptibility of islets to damage. In order to determine the effects of IGF2 overexpression on islet transplantation, and in particular on the graft β-cell mass, we transplanted streptozotocin-diabetic rats with syngeneic islets infected with an adenovirus encoding for IGF2.
Materials and Methods
Animals
Animal experimental procedures were reviewed and approved by the ethical committee of the University of Barcelona. Male inbred Lewis rats (Harlan Laboratories), aged 7–10 weeks, were used as donors and recipients of transplantation. The recipients were made diabetic by a single intraperitoneal injection of streptozotocin (STZ) (Sigma Immunochemicals, St. Louis, MO, USA), 60 mg/kg body weight, freshly dissolved in citrate buffer (pH 4.5). Diabetes was confirmed by the presence of hyperglycemia, polyuria, and no weight gain. Only rats with a blood glucose of >20 mmol/L on a minimum of two consecutive measurements were transplanted. Blood glucose was determined between 9:00 and 11:00 AM in nonfasting conditions. Blood was obtained from the snipped tail, and glucose was measured with a portable meter (Glucocard Memory, A. Menarini Diagnostics, Barcelona, Spain).
Experimental Groups
To determine the effects of IGF2 overproduction on the metabolic outcome of the graft, STZ-diabetic rats were randomly transplanted with 800 islets infected with an adenovirus (Ad) encoding for IGF2 (Ad-IGF2 group, n = 7), encoding for luciferase (Ad-Luc, control vector group, n = 7), or with uninfected islets (control uninfected group, n = 7). Eight hundred islets is a borderline β-cell mass that can restore normoglycemia in some, but not all, STZ-diabetic rats (4). The grafts were removed on day 60 after transplantation. One day before graft harvesting an intraperitoneal glucose tolerance tests (IPGTT) was performed. After graft removal, blood glucose was determined to assess the recurrence of hyperglycemia. A group of control nontransplanted rats (n = 6) had their blood glucose and body weight determined weekly.
To determine the effects of IGF2 on β-cell replication, apoptosis, and mass, STZ-induced diabetic rats were randomly transplanted with 500 Ad-IGF2-infected islets or with 500 uninfected islets. Transplantation of 500 islets provides a clearly insufficient β-mass to restore normoglycemia in this model, and the animals were expected to remain hyperglycemic after transplantation (4). Since glucose is known to modify β-cell replication, apoptosis, and mass, we used this model to ensure comparable metabolic conditions among groups throughout the study. Since cytopathic effects have been reported in association with adenoviral infection (3), we compared the Ad-IGF2-infected islets with the potentially better control group of uninfected islets. Grafts were removed 3 (n = 12), 10 (n = 12), and 28 (n = 12) days after transplantation (six groups, n = 6 for each experimental group). A group of 8- to 10-week-old normal rats (n = 5) had their pancreas harvested to determine nomal β-cell replication and apoptosis.
Recombinant Adenoviruses
E1/E3-deleted adenoviral vector was used for islet transfection. Adenovirus encoding for green fluorescent protein (Ad-GFP) was used to assess the efficiency of infection. The adenoviruses Ad-GFP, Ad-Luc, and Ad-IGF2 were generated by Pacific Northwest Research Institute (Seattle, WA, USA). The transgene was driven by the cytomegalovirus (CMV) promoter.
Islet Isolation and Gene Transfer
Islets were isolated by collagenase (Collagenase P; Roche Diagnostics, Mannheim, Germany) digestion of the pancreas as previously described (31). Isolated islets were hand-picked under a stereomicroscope two or three times, until a population of pure islets was obtained. Islets were washed in serum-free RPMI 1640 11.1 mmol/L glucose (Sigma Immunochemicals) supplemented with 100 U/ml penicillin and 100 μg/ml streptomycin before infection. Groups of 200 islets were infected with Ad-IGF2 or with Ad-Luc at a plaque-forming unit (pfu) of 7 × 107 in 400 μl of serum-free RPMI 1640 11.1 mmol/L glucose for 2 h at 37°C and 5% CO2. After infection, islets were washed three times in RPMI 1640 containing 10% heat-inactivated fetal calf serum (FCS) and incubated overnight in non-tissue culture-treated plastic ware at 37°C in serum-containing medium at 11.1 mmol/L of d-glucose. Uninfected islets had the same treatment as infected islets, but in the absence of adenovirus.
Islet Transplantation and Graft Harvesting
After overnight incubation, Ad-IGF2-infected, Ad-Luc-infected, and uninfected islets were counted into groups of 500 or 800 islets and transplanted under the left kidney capsule of the recipients (31). To harvest the graft, the kidney capsule surrounding the graft was incised and removed with the graft. The grafts were immediately immersed in 4% paraformaldehyde-PBS, were fixed overnight, and, after removal of any excess paraformaldehyde by capillary action, were weighed (31).
Intraperitoneal Glucose Tolerance Test
The intraperitoneal glucose tolerance test (IPGTT) was performed 1 day before graft harvesting. Glucose (2 g/kg) was administered intraperitoneally to conscious animals after a 2-h fast. Blood glucose was measured at 0, 30, 60, 90, and 120 min after injection. Nondiabetic nontransplanted rats (n = 6) were used as normal controls. Total area under the curve (AUC) was determined using the trapezoidal rule method.
Efficiency of Infection
The efficiency of infection was quantified in Ad-GFP-infected islets 24 h after infection, islets were dispersed into single cells and analyzed by flow cytometry, as previously described (45).
IGF2 production was determined by immunohistochemistry in islets before and after transplantation. Graft sections were incubated overnight at 4°C with a rabbit anti-human IGF2 antibody (final dilution 1:100) (Novozymes GroPep, Adelaide, Australia). Preparations were visualized with LSAB+ System-HRP (DakoCytomation, Carpinteria, CA, USA).
Insulin Secretion
To determine whether IGF2 overexpression modified β-cell function, glucose-stimulated insulin secretion was used as a functional assay. Ad-IGF2-infected and control uninfected islets were cultured for 2 and 7 days with RPMI 1640 medium supplemented with 11.1 mmol/L glucose and 10% FCS. Cultured islets were preincubated 1 h at 37°C in triplicate groups of 10 islets in 1 ml of fresh Krebs–Ringer bicarbonate buffer supplemented with Hepes and bovine serum albumin (BSA) (KRBH buffer) containing 2.8 mmol/L glucose. The medium was removed, and islets were incubated at 37°C with 1 ml of KRBH buffer containing 2.8 or 16.7 mmol/L glucose for an additional hour with continuous shaking (45). Supernatants were removed and stored at −80°C until assayed for insulin. Insulin was measured by ELISA (Mercodia Rat insulin ELISA, Mercodia AB, Uppsala, Sweden) and was corrected by DNA content. Islets were disrupted by sonication, and DNA content was determined by a fluorimetric assay, as previously described (45).
β-Cell Apoptosis
Sections were double-stained by immunoperoxidase for apoptotic nuclei using the terminal deoxynucleotidyl transferase biotin-dUTP nick end labeling (TUNEL) technique (In Situ Cell Death Detection Kit, ApopTag; Intergene, Oxford, UK) (5,28) and by alkaline phosphatase for the endocrine non-β-cells of the islets. A cocktail of antibodies (DakoCytomation) including rabbit anti-swine glucagon (final dilution 1:1,000), rabbit anti-human somatostatin (final dilution 1:1,000), and rabbit anti-human pancreatic polypeptide (final dilution 1:500) was used (5). When assessing apoptotic nuclei, we excluded necrotic regions. β-Cell apoptosis was also determined in the pancreases of normal Lewis rats. β-Cell apoptosis in islet grafts and in control pancreas was expressed as percentage of TUNEL-positive β-cells, and a minimum of 1,200 β-cells were counted per sample.
β-Cell Replication
Rats were injected with the thymidine analogue 5-bromo-2′deoxyuridine (BrdU; Sigma Immunochemicals, St. Louis, MO, USA), 100 mg/kg body weight IP, 6 h before removing the graft (27). Sections were double-stained with immunoperoxidase for BrdU using a cell proliferation kit (Amersham, Amersham, UK) with a modified protocol (5,28) and for endocrine non-β-cells of the islets using the cocktail of antibodies described above. Sections were also double-stained for BrdU and for β-cells with a rabbit anti-human insulin antibody (final dilution 1:50) (Santa Cruz Biotechnology, Santa Cruz, CA, USA). β-Cell replication was also measured in the pancreases of normal Lewis rats injected with BrdU 6 h before pancreas excision. β-Cell replication in islet grafts and in control pancreas was expressed as percentage of BrdU-positive β-cells, and a minimum of 1,200 β-cells were counted per sample.
Individual β-Cell Area
The mean cross-sectional area of individual β-cells, a measure of β-cell size, was determined using image-analytical software (AnalySIS 3.0; Soft Imaging System, Münster, Germany). The individual β-cell area on the day of transplantation was determined in sections from the five groups of 500 isolated islets that were used to measure the initially transplanted β-cell mass. For both grafts and isolated islets, the perimeter of the β-cell tissue on a random field was carefully traced on the computer's monitor to exclude any other tissue, and the total β-cell area and β-cell nuclei (392 ± 25 nuclei per sample) in that field were determined. To calculate the area of the individual β-cells, the total β-cell area in the field was divided by the number of β-cell nuclei (5,28).
β-Cell Mass
β-Cell mass was measured by point-counting morphometry (52), as previously described (28,31). β-Cell mass was obtained by multiplying the weight of the graft by the relative β-cell volume. In addition, the β-cell mass of islets at the time of transplantation was determined in five groups of 500 islets isolated on different days. Islets were pelleted and then fixed and weighed as described for islet grafts. The islets β-cell mass was obtained by multiplying the weight of the islets by the percentage of β-cell volume, determined with image-analytical software (AnalySIS 3.0; Soft Imaging System) on sections of the islet pellets.
Statistical Analysis
Statistics were performed using SPSS 14.0 (Chicago, IL, USA) for Windows and differences among groups were evaluated using the Kruskal–Wallis H test, and when determined as significant (p < 0.05), the Mann–Whitney U test was used to determine specific differences between groups. Subsequently, the Bonferroni correction was used as post hoc test to adjust the type I error (a) for multiple testing. Since the use of adjustments for multiple tests has been questioned, as it increases the likelihood of type II errors, the result of the Mann–Whitney U test is presented when it is statistically significant, and it is followed by the adjustment with the Bonferroni correction. Results are expressed as median values and interquartile range (IQR). In figures, mean ± SEM is given to describe the data as it is a better known and more widely used descriptor. The Kaplan–Meier method was used to analyze the metabolic evolution after transplantation, and the differences among the survival curves were compared by the log-rank test with subsequent Bonferroni correction.
Results
Efficiency of Adenoviral Infection
At 24 h after infection 100% of islets and 20% of the individual islet cells were infected, as indicated by GFP production. We have previously shown that this rate of infection is sufficient to double β-cell replication in vitro in Ad-IGF2 infected islets (13). The expression of the transgene was predominantly found in peripheral cells of the islets. Specific staining with IGF2 antibody showed a clear production in Ad-IGF2-infected islets, both before transplantation and at all time points after transplantation (Fig. 1). No IGF2 expression was found in noninfected islets or in the renal parenchyma surrounding the transplanted Ad-IGF2-infected islets.
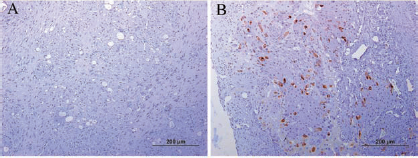
Insulin-like growth factor II (IGF2) expression (brown) in islet grafts from control uninfected group (A) and from adenovirus infected (Ad)-IGF2 group on day 10 after transplantation (B).
β-Cell Function in Infected Islets
Glucose-stimulated insulin secretion was similar in Ad-IGF2-infected islets and in uninfected islets 48 h and 7 days after infection, indicating that β-cell function was unaffected in infected islets overexpressing IGF2 (data not shown).
IGF2 Overexpression Improved the Metabolic Outcome of Islet Grafts
Blood Glucose
To explore the effects of IGF2 overexpression on the outcome of islet grafts, STZ-diabetic rats were transplanted with 800 syngeneic islets, a marginal β-cell mass to achieve normoglycemia. All animals transplanted with 800 Ad-IGF2-infected islets achieved normoglycemia on day 10 after transplantation and remained normoglycemic until graft removal on day 60 (Fig. 2A). In contrast, only 14% of animals transplanted with 800 Ad-Luc infected islets or with 800 control uninfected islets were normoglycemic on day 10 after transplantation. Normoglycemia was eventually achieved on days 21–28 after transplantation in most but not all rats (71%) transplanted with uninfected islets or with Ad-Luc-infected islets. The improved metabolic evolution of Ad-IGF2 group was confirmed by the statistically significant difference in the Kaplan–Meier survival curve (Fig. 2B). In all normoglycemic rats, graft harvesting was followed by recurrence of severe hyperglycemia.

Metabolic evolution of streptozotocin (STZ)-diabetic Lewis rats transplanted with 800 Ad-IGF2 islets (squares), with 800 Ad-luciferase (Luc) islets (triangles), or with 800 control uninfected islets (rhombus) and of nondiabetic nontransplanted animals (crosses). (A) Nonfasting blood glucose. Tx indicates transplantation day, and the arrow indicates the removal of the graft. Values are the mean. ∗p < 0.05 versus nondiabetic nontransplanted animals (Mann–Whitney U test). After adjusting for multiple comparisons, all differences except day 40, between nondiabetic nontransplanted animals and control uninfected and Ad-Luc groups remained statistically significant. (B) Kaplan–Meier survival curve showing differences in percentage of normoglycemic animals over time among Ad-IGF2 (squares), Ad-Luc (triangles), and uninfected (rhombus) groups (log-rank, χ2 = 19.39, p = 0.0001). The percentage of normoglycemic animals over time differed significantly between Ad-IGF2 group and control uninfected group (log-rank, χ2 = 11.48, p = 0.001) and between Ad-IGF2 group and Ad-Luc group (log-rank, χ2 = 11.48, p = 0.001), but not between the control uninfected and Ad-Luc groups (log-rank, χ2 = 0.17, p = 0.67). Differences remained statistically significant after adjusting for multiple comparisons (p < 0.005). (C) Intraperitoneal glucose tolerance test. Kruskal–Walis, p < 0.05 among all groups at each time point, Mann–Whitney, ∗p < 0.05 versus all other groups except Ad-IGF2 at time 120. †p < 0.05 between Ad-IGF2 group and control uninfected group. #p < 0.05 between Ad-IGF2 group and Ad-Luc group. Differences among transplanted groups were not statistically significant after multiple comparison correction.
Intraperitoneal Glucose Tolerance Test
The IPGTT was performed in all three groups transplanted with 800 islets and in control non-STZ-injected rats. All transplanted groups showed glucose intolerance with higher hyperglycemia after glucose injection than control nontransplanted non-STZ-injected animals (Fig. 2C). Nevertheless, glucose intolerance was milder in Ad-IGF2 group that was less hyperglycemic than uninfected control and Ad-Luc groups 30, 60, or 90 min after glucose injection and achieved glucose values similar to control non-STZ-injected animals after 120 min. The AUC was also lower in Ad-IGF2 group [1,635 mmol/L·min (IQR: 902)] than in control uninfected group [2,339 mmol/L·min (IQR: 1,054), p = 0.035)] or in Ad-Luc [2,697 mmol/L·min (IQR: 1,694), p = 0.064], although the differences did not remain statistically significant after adjustment for multiple comparisons.
IGF2 Overexpression Increased Transplanted β-Cell Replication and Mass
Since glucose is known to modify β-cell replication, apoptosis, and mass, the differences in blood glucose levels between Ad-IGF2 and control uninfected groups transplanted with 800 islets could preclude a valid comparison. Thus, to ensure comparable metabolic conditions among grafts from Ad-IGF2 group and control uninfected group throughout the study, STZ-diabetic recipients were transplanted with 500 islets, a β-cell mass clearly insufficient to restore normoglycemia in this model. As planned, both groups showed similar hyperglycemia throughout the study (Table 1).
Blood Glucose and Body Weight in STZ-Diabetic Rats Transplanted With 500 Islets
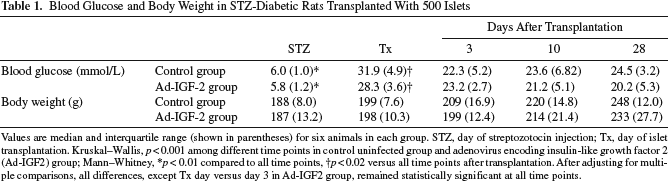
Values are median and interquartile range (shown in parentheses) for six animals in each group. STZ, day of streptozotocin injection; Tx, day of islet transplantation. Kruskal–Wallis, p < 0.001 among different time points in control uninfected group and adenovirus encoding insulin-like growth factor 2 (Ad-IGF2) group; Mann–Whitney
p < 0.01 compared to all time points
p < 0.02 versus all time points after transplantation. After adjusting for multiple comparisons, all differences, except Tx day versus day 3 in Ad-IGF2 group, remained statistically significant at all time points.
β-Cell Replication
At all time points after transplantation, β-cell replication was higher in grafts of Ad-IGF2 group than in grafts of control uninfected group [day 3: 1.45% (IQR: 0.26) vs. 0.58% (IQR: 0.18), p = 0.006; day 10: 1.58% (IQR: 1.40) vs. 0.90% (IQR: 0.61), p = 0.035; day 28: 1.35% (IQR: 0.35) vs. 0.64% (IQR: 0.28), p = 0.004] and than β-cell replication in normal pancreas [0.36% (IQR: 0.17), p < 0.02; p < 0.05 when it was adjusted for multiple comparisons] (Fig. 3B). Although glucose is a well-known stimulator of β-cell proliferation, in control uninfected group β-cell replication was significantly increased only on day 10 after transplantation, but not on days 3 and 28, compared to normal pancreas, confirming previous observations indicating that β-cell replication is impaired initially after transplantation (5) and after long-term exposure to hyperglycemia (5,46).

(A) Islet graft from Ad-IGF2 group on day 10 after transplantation. Section was double-stained for bromodeoxyuridine (BrdU) (black nuclei, clearly shown in inset) and for insulin (brown cytosolic staining). (B) β-cell replication in normal pancreas (black bar) and in 500-islet grafts from control uninfected (white bars) and Ad-IGF2 (hatched bars) groups. Values are mean ± SEM. Kruskal–Wallis, p < 0.02 among control pancreas and control uninfected groups and among control pancreas and Ad-IGF2 groups, Mann–Whitney, ∗p < 0.02 versus control pancreas. Differences remained significant after adjusting for multiple comparisons (p < 0.05). †p < 0.05 between control uninfected and Ad-IGF2 groups.
β-Cell Apoptosis
β-Cell apoptosis was significantly increased in transplanted islets from both control uninfected and Ad-IGF2 groups compared with normal pancreas at all time points (p < 0.01 and p < 0.05 when adjusted for multiple comparisons) (Fig. 4A). There were no differences between Ad-IGF2 and uninfected groups.
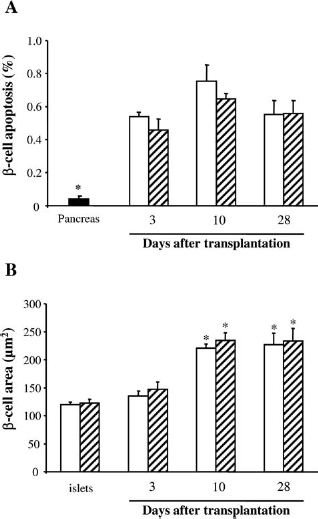
(A) β-Cell apoptosis in normal pancreas (black bar) and in 500-islet grafts from control uninfected (white bars) and Ad-IGF2 (hatched bars) groups. Values are mean ± SEM. Kruskal–Wallis, p < 0.01 among control pancreas and control uninfected groups and among control pancreas and Ad-IGF2 groups, Mann–Whitney, ∗p < 0.01 versus all other groups. Differences remained significant after adjusting for multiple comparisons (p < 0.05). (B) Cross-sectional area of individual β-cells in isolated islets cultured overnight as for transplantation, and in 500-islet grafts from control uninfected (white bars) and Ad-IGF2 (hatched bars) groups. Values are mean ± SEM. Kruskal–Wallis, p < 0.002 among control uninfected groups and among Ad-IGF2 groups, Mann–Whitney, ∗p < 0.01 versus control uninfected islets and Ad-IGF2-overexpressing islets. Differences remained significant after adjusting for multiple comparisons (p < 0.02).
Individual Cross-Sectional β-Cell Size
The individual cross-sectional area of β-cells increased significantly in islet grafts on days 10 and 28 after transplantation compared with the individual area of β-cells in isolated islets (p < 0.01 and p < 0.02 when adjusted for multiple comparisons) (Fig. 4B). Individual β-cell size was similar in Ad-IGF2 and uninfected groups.
β-Cell Mass
β-Cell mass was reduced on day 3 after transplantation in control uninfected [0.38 mg (IQR: 0.19)] and Ad-IGF2 [0.36 mg (IQR: 0.26)] groups compared with the initially transplanted β-cell mass [control uninfected: 1.60 mg (IQR: 0.22), p = 0.006; Ad-IGF2: 1.61 mg (IQR: 0.15), p = 0.006; p = 0.018 in both cases when adjusted for multiple comparisons] (Fig. 5). In Ad-IGF2 group transplanted β-cell mass increased on day 10 to a value similar to the initially transplanted β-cell mass [1.27 mg (IQR: 0.91)], and on day 28 it was higher than that of control uninfected group [0.63 mg (IQR: 0.38) vs. 0.42 mg (IQR: 0.31), p = 0.008], reflecting the higher β-cell replication of Ad-IGF2 transplanted islets. In contrast, in control uninfected group, the graft β-cell mass was always lower than the initially transplanted β-cell mass (p < 0.02 and p < 0.05 when adjusted for multiple comparisons). Although it increased transitorily on day 10 after transplantation [1.05 mg (IQR: 0.50)], in agreement with the higher β-cell replication and individual β-cell size at this time point, it was reduced again on day 28 when β-cell replication was impaired and could not compensate the increased β-cell apoptosis.
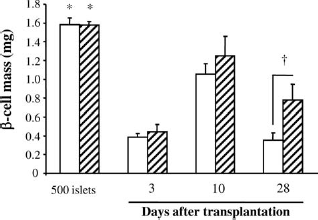
β-Cell mass in 500 isolated islets cultured overnight as for transplantation and in 500-islet grafts from control uninfected (white bars) and Ad-IGF2 (hatched bars) transplanted groups. Values are mean ± SEM. Kruskal–Wallis, p < 0.005 among control uninfected groups and among Ad-IGF2 groups, Mann–Whitney, ∗p < 0.02 versus all other groups except Ad-IGF2 on day 10 after transplantation. Differences remained significant after adjusting for multiple comparisons (p < 0.05) except between 500 Ad-IGF2 isolated islets and Ad-IGF2 group on day 28 after transplantation. †p < 0.01 between control uninfected group and Ad-IGF2 group.
Discussion
In this study, we show that transplanted islets overexpressing IGF2 maintained a sustained increment in β-cell replication that led to the regeneration of the engrafted β-cell mass and to the early achievement of sustained normoglycemia after transplantation to diabetic recipients. IGF2 overexpression did not modify insulin secretion in islets studied in vitro, did not reduce β-cell apoptosis, and did not enhance β-cell hypertrophy in transplanted islets, suggesting that the higher β-cell mass and improved metabolic control in IGF2-overexpressing grafts resulted from the increased β-cell replication.
IGF1 and IGF2 are β-cell mitogens, and they also promote β-cell survival. The biological effects of IGF2 are mediated primarily by type 1 IGF receptor, which is abundant on β-cells (14,49). IGF2 can also act through the insulin receptor (39). IGF2 expression is higher in fetal life, but it declines and in rodents is extinguished in most tissues after birth. In contrast, IGF1 expression in fetal life is low and does not rise to adult levels until weaning. In early neonatal life, there is a wave of apoptotic β-cell death (42) that has been linked to the concomitant reduction in IGF2 expression (17,34). In adult islets, incubation of islets with IGF2 improves islet cell viability (13,20,37), and IGF2 survival action has been also shown in rats fed with a low protein diet (36). However, we did not detect a reduction in β-cell apoptosis in islet grafts that overexpressed IGF2. The difficulties of preventing the massive loss of islet tissue, up to 70% of the transplanted β-cell mass, that takes place in the early posttransplant period were recently reviewed (11). β-Cell apoptosis peaks around day 3 after transplantation and plays a major role in this early β-cell death (5,11). The cause of the initial β-cell apoptosis is multifactorial (12), with hypoxia playing a fundamental role (10), and other factors such as anoikis (47), nonspecific inflammation (29,38,46), and hyperglycemia (5,15,26,33) also contributing to islet damage. Several strategies that have successfully increased the long-term outcome of islet transplantation have been, however, unable to increase the initial survival of islets (11). We have also met this difficulty in previous experiments in which we found that the beneficial effects of normoglycemia (25) and IL-1 receptor antagonist protein (46) in the outcome of islet transplantation were not associated with a reduction of early β-cell apoptosis. Robitaille et al. (37) reported that preincubation of encapsulated islets with IGF2 before transplantation increased the percentage of diabetic mice that achieved normoglycemia, but apoptosis was not determined in the islet grafts. The lack of effect of IGF2 on transplanted β-cell survival, along with previous data showing similar negative results with other therapeutic interventions, highlights the difficulty of reducing the initial β-cell death in islet transplantation. It is unknown whether a multifactorial approach would prove more successful.
IGF2 overexpression resulted in sustained β-cell hyperplasia of transplanted islets that showed throughout the study higher proliferation than uninfected transplanted islets or than endogenous pancreatic islets of normal rats. Although glucose is a very potent stimulator of β-cell proliferation (6), on day 3 after transplantation β-cell replication was not significantly higher in islet grafts of control uninfected group than in islets of normal, non-STZ-injected nontransplanted, control rats, confirming previous data indicating that β-cell replication is impaired in the early days after transplantation (5). The limitation in β-cell replication in the initial days after transplantation is mediated, at least partly, by IL-1β-mediated nonspecific inflammation (29,45). The increased replication on day 3 after transplantation in IGF2-overexpressing grafts suggests that IGF2 is able to partly prevent the suppressive effects of IL-1β on β-cell replication in vivo and is in agreement with recently published in vitro data (13). On the other hand, glucose-induced β-cell replication is not maintained when islets are exposed to chronic hyperglycemia (5,21,30), as shown in the transplanted islets of control uninfected group exposed to 28 days of sustained hyperglycemia. The increased β-cell replication in Ad-IGF2 group on day 28 after transplantation indicates that IGF2 overexpression was able to overcome this limitation. The mechanism by which IGF2 increased β-cell replication in islets exposed to chronic hyperglycemia remains unknown. As described for IGF1 (53), overexpression of IGF2 in islet cells could increase the expression of insulin receptor substrate 2 (IRS-2), a protein playing a crucial role in β-cell growth.
IGF2 has been reported to inhibit, increase, or have no effect on insulin secretion (9,16,24,55). We found that glucose-stimulated insulin secretion was not modified in Ad-IGF2 islets after 2 and 7 days in culture. This normal insulin secretion in Ad-IGF2 islets suggests that the improved metabolic outcome in rats transplanted with IGF2 overexpressing islets was due to the increased β-cell mass. IGF2 overexpression did not increase the size of individual β-cells in agreement with previous data indicating that IGF2 overexpression in transgenic mice did not induce α- or β-cell hypertrophy (9). Thus, since IGF2 overexpression did not modify insulin secretion, did no reduce transplanted β-cell death, and did not induce β-cell hypertrophy, we postulate that the increased β-cell mass in islet grafts overexpressing IGF2 and improved the metabolic outcome resulted from the sustained increment of β-cell replication throughout the first month after transplantation.
In clinical islet transplantation, a high islet mass must be transplanted to achieve normoglycemia, typically requiring two or three islet transplant procedures (23,40,44). The transplantation process results in a dramatic loss of islets leading to the successful engraftment of a β-cell mass that is marginally sufficient to maintain normoglycemia and any subsequent reduction, even from mild insults, may result in the recurrence of hyperglycemia (23). Since the preservation of the initially transplanted β-cell mass has proven to be particularly challenging, an alternative or complementary strategy to increase the grafted β-cell mass would be to enhance the replication of transplanted β-cells. In this study, we have shown that a therapeutic intervention targeting β-cell replication was able to regenerate the transplanted β-cell mass and to improve the outcome of islet transplantation. The use of adenoviral vectors in clinical islet transplantation is unlikely, but it is a useful model to investigate the benefits of local delivery of treatments to islet grafts. The results indicate that overexpression of IGF2 to the islet grafts was beneficial. IGF2 overexpression induced a sustained increment in β-cell replication in transplanted islets, with no changes in β-cell apoptosis or individual cell size, and increased the β-cell mass in the islet graft. The improved metabolic outcome of the diabetic recipients indicates that strategies that increase β-cell replication can increase the grafted β-cell mass and reduce the β-cell number required to achieve normoglycemia.
Footnotes
Acknowledgments
This work was supported by grants from the Juvenile Diabetes Foundation International (1-2002-687) and Instituto de Salud Carlos III (PI06/0891 and PI10/00636). CIBER of Diabetes and Associated Metabolic Diseases (CIBERDEM) is an initiative from ISCIII. Elisabet Estil·les was supported by a grant from Fundació Privada IDIBELL. The authors declare no conflicts of interest.