
Abstract
Cryopreserved hepatocytes with good hepatospecific functions upon thawing are important for clinical transplantation and for in vitro drug toxicity testing. However, cryopreservation reduces viability and certain hepatospecific functions, but the most pronounced change is diminished attachment efficiency of hepatocytes. Adhesion of cells to the extracellular matrix and cell–cell contacts are crucial for many aspects of cellular function. These processes are partly mediated and controlled by cellular adhesion molecules. The mechanisms responsible for reduced attachment efficiency of cryopreserved hepatocytes are not well understood. To address this question, we investigated the effect of a new cryopreservation procedure, using wheat proteins (WPs) or mixtures of recombinant forms of wheat freezing tolerance-associated proteins, on the stability of three important adhesion molecules (β1-integrin, E-cadherin, and β-catenin). Immunoblot analyses revealed that the levels of β1-integrin, E-cadherin, and β-catenin were much lower in cryopreserved rat hepatocytes, when compared to fresh cells. Protein expression of the adhesion molecules was generally lower in cells cryopreserved with DMSO, compared to WPs. Moreover, the stability of the adhesion molecules was not affected by cryopreservation to the same degree, with more pronounced decreases occurring for β1-integrin (62–74%) > β-catenin (51–58%) > E-cadherin (21–37%). However, when hepatocytes were cryopreserved with partially purified WPs (SulWPE, AcWPE) or with mixtures of recombinant wheat proteins, there was a clear protective effect against the loss of protein expression of β1-integrin, E-cadherin, and β-catenin. Protein expression was only 10–20% lower than that observed in fresh hepatocytes. These findings clearly demonstrate that WPs, and more particularly, partially purified WPs and recombinant wheat proteins, were more efficient for cryopreservation of rat hepatocytes by maintaining good expression of these adhesion molecules. These promising results could lead to a new and improved cryopreservation technology for applications such as clinical transplantation of hepatocytes.
Introduction
Cell adhesion to the extracellular matrix (ECM) plays a central role in many complex biological processes, including cell shape, motility, growth, differentiation, and survival (1,12). In particular, epithelial cells are attached to basement membranes by a variety of adhesive ECM glycoproteins, including collagen, laminin, fibronectin, and proteoglycans. Cell–matrix attachment is crucial for three-dimensional cellular architecture, the establishment of basal–apical polarity, and the maintenance of tissue integrity (3,20). Cell–cell contacts are also crucial to many aspects of cellular function, such as growth, differentiation, homeostasis, and wound repair. These important processes are partly mediated and controlled by a number of cellular adhesion molecules that exhibit a well-regulated spatiotemporal pattern of expression during development and regeneration (24).
The most prominent family of ECM adhesion receptors is represented by the integrins. These are heterodimeric transmembrane proteins composed of α and β subunits, with extracellular domains that bind to the ECM, and cytoplasmic domains associated with the actin cytoskeleton and other affiliated proteins, including vinculin, talin, paxillin, and α-actinin (18). Integrin-mediated attachment to the ECM plays an important role in regulating cell survival and proliferation. A decrease in levels of β1-integrin is known to cause a loss of functional activities at focal adhesions (17). β1-Integrin-mediated attachment to the ECM is necessary for survival of hepatocytes (38).
Adhesion molecules that are involved in cell–cell contacts have been classified into several molecular families (20,44). The most important is the cadherin family (Ca2+-dependent adhesion molecules). This gene superfamily encodes transmembrane proteins that regulate Ca2+-dependent cell-cell adhesion (58) and play a role in maintaining the integrity of multicellular structures. α-Catenin is linked to the cytoplasmic domain of cadherins via β- or γ-catenin but does not directly bind to cadherins (35). Due to high homology with vinculin, it was suggested that α-catenin links the cadherin/catenin complex to the actin cytoskeleton (57). The loss of E-cadherin destabilizes the structural integrity of the epithelium and causes cells to dissociate from their neighbors (20). E-cadherin was originally named uvomorulin or liver cell adhesion molecule (L-CAM), thus suggesting that it is a major regulator of hepatocyte intercellular adhesion (16,42).
β-Catenin is a multifunctional protein that is tightly regulated in cells. It plays a pivotal role in cell-cell adhesion, where it has a structural role at adherens junctions, linking E-cadherin to the actin cytoskeleton (59). E-cadherin and β-catenin were colocalized at contact regions in cultured hepatic cells (30). Cytoplasmic β-catenin is an important component of the Wnt signaling cascade, which regulates cell proliferation. β-Catenin undergoes translocation to the nucleus where it forms heterodimeric complexes with the T-cell factor/lymphoid enhancement factor family and is responsible for transactivating target genes (23,60). The Wnt/β-catenin pathway has been shown to be important in liver growth and development (2,33). β-Catenin was localized at the hepatocyte membrane in a normal adult rat liver with some cytoplasmic staining (33). Furthermore, β-catenin undergoes regulation during Matrigel-induced differentiation and maturation of rat hepatocytes, which results in increased stabilization and translocation to the membrane.
Primary cultures of hepatocytes have been extensively used as a model system to investigate liver function, including drug metabolism, hepatotoxicity, protein biosynthesis, and gene expression. Freshly isolated mature hepatocytes spread out rapidly and form monolayers when they are cultured in plastic dishes but they show reduced liver-specific functions (6). However, when they are cultured in plastic dishes coated with liver-derived proteoglycan (26,55) or a synthetic substratum (54), they can aggregate into multicellular spheroids, retain their normal cell polarity, and express certain differentiated functions for prolonged periods of time.
Hepatocytes isolated from human donor livers are being used currently for clinical transplantation in patients with acute liver failure (ALF) and liver-based metabolic defects (52). This approach is being developed as a less invasive alternative to orthotic liver transplantation. Successful cryopreservation of hepatocytes with good hepatospecific function upon thawing would allow the establishment of cell banks. This would ensure that large quantities of hepatocytes are readily available for emergency use in ALF patients and for repeated treatment in patients with chronic liver disease and metabolic disorders.
Although cryopreservation of human hepatocytes has been successfully used in clinical transplantation, further optimization of cryopreservation protocols is necessary (52). Dimethyl sulfoxide (DMSO), a widely used cell-permeable cryoprotectant, is toxic to hepatocytes, particularly at temperatures such as 25°C and 37°C, and must be removed rapidly from contact with cells during postthaw processing (52). Cryopreservation with DMSO causes decreases in viability and certain hepatospecific functions but the most pronounced change is the diminished attachment efficiency of hepatocytes upon thawing (28, 29). Attachment efficiencies are even lower for cryopreserved human hepatocytes (53,61). This is a major problem during transplantation, where cells need to attach to the hepatic sinusoidal endothelium in the liver during the engraftment process (52). However, the mechanisms responsible for the decrease in attachment efficiency are not well understood. To address this question, we investigated the effect of a new cryopreservation procedure, using purified wheat proteins (WPs) and several wheat recombinant proteins, on the stability of three important adhesion molecules (β1-integrin, E-cadherin, and β-catenin) by Western blot analysis.
Materials and Methods
Chemicals
Collagenase, insulin, Williams' medium E (WME), DMSO, propidium iodide (PI), and other chemicals were from Sigma Chemical Company (St. Louis, MO). Primary antibodies (β1-integrin, E-cadherin, and β-catenin) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Leibovitz medium (L-15), gentamicin, and MEM vitamins were from Gibco/Invitrogen Technologies (Burlington, ON). Fetal bovine serum (FBS) was from Medicorp (Montreal, QC).
Plant Materials and Growth Conditions
Winter wheat [Triticum aestivum L. cv Clair, LT50 (lethal temperature that kills 50% of seedlings) −19°C] was used in this study. Wheat plants were grown as previously described (13,21). Briefly, plants were grown for 10 days under a 16-h/day photoperiod with a light intensity of 250 μmol/m2 s at 20°C.
Protein Extraction
The aerial parts of the seedlings were homogenized in cold ultrapure water. The homogenate was filtered through three layers of miracloth and centrifuged at 30,000 × g for 45 min at 4°C. The pH of the supernatant was adjusted to 7.4 with NaOH and sterilized using a 0.22-μm filter. The extract was concentrated by freeze-drying and stored at −20°C. The dried powder was resuspended in ice-cold WME medium before being added to the hepatocyte suspension, as described previously (21).
Purification of Wheat Proteins
The WPs extract was prepared as described previously (21). Subsequently, the WPs were partially fractionated by ammonium sulfate precipitation (0–20%, 21–40%, 41–60%, 61–80%, and 81–100%) (SulWPE) or with acetone (55%) (AcWPE). After 1 h, the precipitate was dialyzed against ultrapure water overnight using 12–14,000 MWCO tubing (Spectra/Por, Spectrum Laboratories, Inc., Los Angeles, CA) and freeze-dried.
Production of Recombinant Wheat Proteins
The four wheat cDNA clones (WCS120, WCS19, TaTIL, and TaIRI-2) (13,15,22,36,56) were expressed in the pTrcHis vector in E. coli. The bacteria were cultured in LB medium with ampicillin for 3 h and induced with 1 mM isopropyl β-1-thiogalactopyranoside (IPTG) for 3 h. The proteins were purified on His-tag resin, as previously described (13).
Hepatocyte Isolation
Hepatocytes were isolated from male Sprague-Dawley rats (140–180 g) (Charles River Canada, Saint-Constant, Que), using a two-step collagenase digestion technique (19,43). Animals were maintained and handled in accordance with the Canadian Council on Animal Care guidelines for care and use of experimental animals (37). Cell viability was evaluated with trypan blue by microscopy and with 2 μM PI (40) by flow cytometry (FACScan, Becton Dickinson, Oakville, ON).
Analysis of PI Fluorescence by Flow Cytometry
Mean PI fluorescence intensity of 10,000 cells was calculated for each sample and corrected for autofluorescence obtained from samples of unlabeled cells. The analyzer threshold was adjusted on the forward scatter (FSC) channel to exclude noise and subcellular debris (Fig. 1A, box R1). Photomultiplier settings were adjusted to detect PI fluorescence on the FL3 detector and autofluorescence of hepatocytes on the FL-1 detector (Fig. 1B).

Dot plot for analysis of PI fluorescence by flow cytometry. Following cryopreservation, hepatocytes were analyzed for PI fluorescence by flow cytometry. (A) Hepatocytes are shown within the R1 gate, which excludes noise and subcellular debris. (B) Autofluorescence of hepatocytes is detected using the FL-1 channel, while PI fluorescence is detected with the FL-3 channel in order to distinguish viable and nonviable (PI uptake) cells.
Cryopreservation of Hepatocytes
Immediately after isolation, the hepatocyte suspension (1.5 × 106 cells/ml for postthaw viability tests or 5 × 106 cells/ml for postthaw adherence and immunodetection) was added to ice-cold WME medium supplemented with 10% FBS and different combinations of WPs or recombinant wheat proteins in cold cryovials. Positive (15% DMSO and 50% FBS) and negative (WME) controls were also prepared. Tubes containing cells were frozen at a cooling rate of 1°C/min in a controlled freezing container (Nalgene, Rochester, NY) to −80°C for 1 day, and then transferred to liquid nitrogen (21).
DMSO is used for hepatocyte cryopreservation at concentrations between 10% and 20.5% (9,10,41,47). We tested two different concentrations of DMSO, 10% and 15%, and found that 15% DMSO gave higher viability in rat hepatocytes (data not shown).
The conditions for cryopreservation of 1.5 × 106 cells/ml were 15% DMSO and 50% FBS (DMSO), or 20 mg of WPEs (WPEs), or 20 mg of SulWPE (SulWPE; 41–80% pooled fractions), or 20 mg of AcWPE (AcWPE), or with mixtures of recombinant proteins WCS19-TaTIL (5 mg/20 mg) or WCS120-TaIRI-2 (10 mg/30 mg). For cryopreservation of 5 × 106 cells/ml, quantities of proteins were 60 mg for WPEs, SulWPE, or AcWPE, and mixtures of recombinant proteins were WCS19-TaTIL (15 mg/60 mg) or WCS120-TaIRI-2 (30 mg/90 mg).
Thawing and Culture of Cryopreserved Hepatocytes
Frozen cells were thawed quickly by gentle agitation in a 37°C water bath and viability (70–80%) was determined by flow cytometry with 2 μM PI. Hepatocyte suspensions were diluted 10-fold with cold WME medium, immediately after thawing. After centrifugation (4°C, 50 × g, 2 min), hepatocytes were suspended in 10 ml of WME medium and washed twice. Dead cells were removed by a 30% isotonic Percoll centrifugation step (recovery of 85 %) and viability was determined (90–95%). Postthaw viability was very low in negative (WME) controls (1.6%).
Hepatocytes (2.5 × 105/ml) were cultured in 12-well Corning plates in WME medium supplemented with insulin (0.2 mg/ml) and gentamicin (50 mg/ml) in a humidified atmosphere of 5% CO2 and 95% air at 37°C. After 3 h, medium was changed and cells were incubated overnight in L-15 medium supplemented with insulin and gentamicin.
Cellular Morphology
Morphology of hepatocytes after 24 h in tissue culture plates was evaluated by microscopy (Carl Zeiss Ltd., Montreal, QC) and photographs were taken by digital camera (Camera CCD, Sony DXC-950P; Empix Imaging, Inc., Mississauga, ON). Images were analyzed using Northern Eclipse software.
Viability
Lactate dehydrogenase (LDH) activity was determined in the medium of seeded, adherent hepatocytes (32). The hepatocyte culture medium was removed and activity of LDH released into the medium from adherent cells was quantified. The total amount of LDH activity was obtained by lysis of live cells with 10% Triton X-100 (31).
Attachment Efficiency
Attachment efficiency was determined by measuring LDH activity in attached cells following 3 or 24 h in culture, relative to that obtained in hepatocytes prior to seeding.
Immunodetection of Adhesion Molecules
Cells were cultured for 24 h, then washed with PBS, scraped off plates, and suspended in 100 μl of lysis buffer (20 mM Tris-HCl, 2 mM EGTA, 2 mM EDTA, 6 mM β-mercaptoethanol) and homogenized by sonication. Proteins (40 μg) were separated on 12% SDS-polyacrylamide gels (27) and then blotted onto polyvinylidene fluoride membranes. Membranes were blocked with 5% dry milk in TBS-T buffer (2 mM Tris-HCl, 13.7 mM NaCl, 0.1% Tween) and incubated with primary antibodies (1/500). Protein bands were detected using Western lightning chemiluminescence reagent plus (Perkin-Elmer Life Sciences, Boston, MA) using a horse radish peroxidase (HRP)-conjugated anti-rabbit IgG (1/1000). Protein level was analyzed by densitometry using a Molecular Dynamics scanner (Amersham, Baie d'Urfe, QC) and IP Lab gel software (Scanalytics Inc., Fairfax, VA).
Statistical Analysis
Results were expressed as mean ± SEM for a minimum of three experimental repeats using cell preparations from three separate animals. For each cell preparation, three different protein batches were used for cryopreservation and each condition was analyzed with at least three replicate dishes. Data were normalized to noncryopreserved experimental controls at each time interval in the same experiment. Comparison between groups and analysis for differences between means of control and treated groups were performed using ANOVA followed by the post hoc test Newman-Keuls (p < 0.05).
Results and Discussion
Optimization of Cryopreservation Protocol
We reported recently an improved cryopreservation technology for rat hepatocytes using WPs (21). In the present study, we optimized the cryopreservation procedure using WPs that were partially purified using ammonium sulfate precipitation (SulWPE) or acetone (AcWPE), as well as with recombinant forms of different wheat freezing tolerance-associated proteins. Optimal viability and cryopreservation potential were obtained when hepatocytes were cryopreserved with WPs from the 41–80% fractions. We subsequently evaluated the effects of purified WPs and recombinant wheat proteins on morphology, viability, attachment efficiencies, and the stability of several important cellular adhesion molecules in rat hepatocytes.
Cellular Morphology
Morphological analysis shows that fresh hepatocytes were more spread out and formed monolayers on culture substrates, when compared to cryopreserved cells (Fig. 2). Hepatocytes that were cryopreserved with WPs, purified WPs (SulWPE, AcWPE), or with mixtures of recombinant proteins had similar morphologies (Fig. 2). They were more spread out and had cell–cell contacts, compared to those cryopreserved with DMSO. However, they still had a more rounded phenotype compared to fresh cells.

Effects of WPEs, partially purified WPs, and recombinant proteins on cellular morphology of cryopreserved hepatocytes. Cellular morphology was visualized after 24 h of culture of thawed rat hepatocytes, following cryopreservation for 7 days in WME 10% FBS supplemented with either 15% DMSO and 50% FBS (DMSO), or WPEs, or AcWPE, or SulWPE, or with mixtures of recombinant proteins, WCS120-TaIRI-2 or WCS19-TaTIL. Freshly isolated hepatocytes served as reference control. Hepatocytes (250 × 103) were visualized by microscopy (magnification 10×). Photographs are from one representative experiment, which was repeated at least three times.
Viability and Attachment Efficiencies
Viabilities of hepatocytes following cryopreservation with partially purified WPs (SulWPE, AcWPE) and the recombinant proteins WCS120-TaIRI-2 ranged from 77.32% to 91.27% and were higher than those obtained with WPs (72.15%) or DMSO (66.93%) (Table 1). The highest viability value (91.27%) was obtained with the mixture of WCS120-TaIRI-2 recombinant proteins. This value was comparable to that of fresh cells (92.67%). The attachment efficiencies after 24 h in culture for hepatocytes cryopreserved with SulWPE, AcWPE or mixtures of recombinant proteins ranged from 70.3% to 74.3%, and were also higher compared to values obtained with DMSO (50.3%) or WPs (50.9%) (Table 1). The corresponding numbers of viable cells after 24 h of monolayer culture are also given in Table 1. Attachment efficiencies for DMSO-cryopreserved hepatocytes from various animal species are generally in the range of 50%, compared to fresh cells (9–11,46,48–50). For human hepatocytes, recent advances allow about 50% of batches of hepatocytes isolated from different human livers to be plated as monolayer cultures (“plateable” hepatocytes) after cryopreservation (28). However, attachment efficiencies of “plateable” cryopreserved human cells ranged from 5% to 40% on collagen-coated plates (51,52,61). Cellular adhesion is very important because cell density and cell–cell contacts have an impact on metabolic activities and functional integrity of hepatocytes. Although many investigators have attempted to restore these properties of mature hepatocytes in monolayer cultures of cryopreserved cells, most attempts have failed or gave unsatisfactory results. This could be due to the loss of polarized configuration of hepatocytes (4). The matrix architecture of cultured hepatocytes plays a crucial role in maintaining and restoring hepatic gene expression (4,14,34,39).
Viability and Attachment Efficiency of Rat Hepatocytes Following Cryopreservation With WPs, Purified WPs, and Recombinant Wheat Proteins
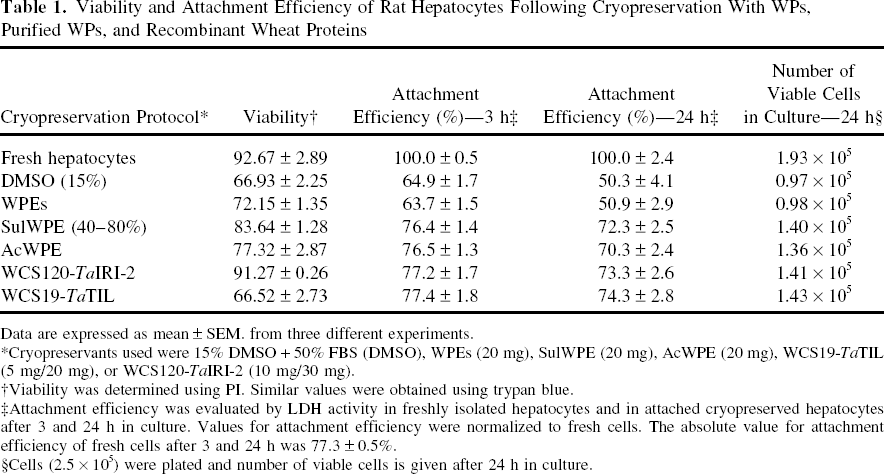
Data are expressed as mean ± SEM. from three different experiments.
Cryopreservants used were 15% DMSO + 50% FBS (DMSO), WPEs (20 mg), SulWPE (20 mg), AcWPE (20 mg), WCS19-TaTIL (5 mg/20 mg), or WCS120-TaIRI-2 (10 mg/30 mg).
Viability was determined using PI. Similar values were obtained using trypan blue.
Attachment efficiency was evaluated by LDH activity in freshly isolated hepatocytes and in attached cryopreserved hepatocytes after 3 and 24 h in culture. Values for attachment efficiency were normalized to fresh cells. The absolute value for attachment efficiency of fresh cells after 3 and 24 h was 77.3 ± 0.5%.
Cells (2.5 × 105) were plated and number of viable cells is given after 24 h in culture.
Immunodetection of Adhesion Molecules
Western blot analysis was used to compare the expression of three important adhesion molecules, β1-integrin, E-cadherin, and β-catenin, between cryopreserved and fresh hepatocytes (Fig. 3). Protein levels for β1-integrin, E-cadherin, and β-catenin were lower in cryopreserved cells, relative to fresh hepatocytes (Fig. 3A). Protein expression of these three adhesion molecules was similar in cells cryopreserved with WPs or DMSO (Fig. 3A). However, the effect of cryopreservation on the levels of adhesion molecules was different, with more pronounced decreases occurring for β1-integrin (62–74%) > β-catenin (51–58%) > E-cadherin (21–37%) (Fig. 3B–D). Hepatocytes that were cryopreserved with WPs, compared to DMSO, maintained higher protein levels of β1-integrin and E-cadherin (Fig. 3C, D). However, the loss of protein ranged from 21% to 62%. This loss of protein expression of the three adhesion molecules was markedly reversed in hepatocytes that were cryopreserved with SulWPE, AcWPE or mixtures of recombinant proteins (Fig. 3A). Proteins levels of adhesion molecules were similar and only 15–20% lower than those in fresh hepatocytes, for all four of the wheat protein samples (Fig. 3B–D). For β1-integrin, β-catenin, and E-cadherin, protein expression was about 60%, 40%, and 20% higher, respectively, for cells cryopreserved with partially purified or recombinant wheat proteins, when compared to DMSO (Figs. 3B–D). These findings clearly show that cryopreservation leads to downregulation of three important cellular adhesion molecules in rat hepatocytes. A decrease in levels of E-cadherin and β-catenin could explain the loss of cell–cell contacts, while the loss of β1-integrin is likely to be involved in the lack of adhesion to the culture substrate. These results demonstrate that partially purified WPs and recombinant wheat proteins were significantly more efficient than DMSO for cryopreservation of rat hepatocytes.

Effects of WPEs, partially purified WPs, and recombinant proteins on protein expression of adhesion molecules in cryopreserved hepatocytes. (A) Protein expression of β-catenin, E-cadherin, and β1-integrin was determined by immunoblotting after 24 h of culture of rat hepatocytes, following cryopreservation for 7 days in WME supplemented with either 15% DMSO and 50% FBS (DMSO), or WPEs, or SulWPE, or AcWPE, or with mixtures of WCS120-TaIRI-2 or WCS19-TaTIL. Freshly isolated hepatocytes (Fresh) served as reference. Protein expression of (B) β-catenin, (C) β1-integrin, and (D) E-cadherin was analyzed by densitometry, using GAPDH as loading control. Data (mean ± SEM) represent triplicate measurements from four independent experiments (n = 12). *p < 0.05, **p < 0.01, ***p < 0.001.
There is very little information about the effect of cryopreservation on attachment molecules in hepatocytes. It was reported recently that 22 genes including integrins, cadherins, and catenins were downregulated after cryopreservation of human hepatocytes with DMSO (52). Gene and protein expression of E-cadherin and β1-integrin were significantly downregulated in cryopreserved cells. There was a link between the decrease in β1-integrin and the loss of attachment. The cytoplasmic domain of E-cadherin underwent cleavage, suggesting that cryopreservation could increase degradation of adhesion molecules. Our findings confirm the downregulation of β1-integrin, E-cadherin, and β-catenin in rat hepatocytes. An important finding is that cryopreservation with partially purified WPs and recombinant wheat proteins showed promising potential by significantly reversing these effects.
The higher cryopreservation capacity of WPs, Sul WPE, and AcWPE, compared to DMSO, is probably explained by protective effects of WPs against degradation of the adhesion molecules by unknown processes. The wheat proteins may protect the plasma membrane against cryopreservation-induced damage. It is also possible that WPs enhance the association and stability of adhesion molecules at the hepatocyte membrane. WPs could protect the Ca2+-sensing receptor and thus conserve the E-cadherin-mediated cell-cell contacts. WPs could also protect Src-family tyrosine kinases that are clearly an essential part of the Ca2+-induced E-cadherin signaling pathway. The tyrosine phosphorylation of β- and γ-catenin increased the association of α-catenin with E-cadherin in differentiating keratinocytes (7). This strengthening of cell adhesion was due to the bridging ability of α-catenin between the cadherin/catenin complex and the actin cytoskeleton. The inhibition of Src-family tyrosine kinases suppressed the formation of adherens junctions, disrupted catenin/cadherin complexes, and decreased cell adhesion (7).
The higher capacity of recombinant proteins to protect the adhesion molecules could be related to their native functions in wheat. WCS120 is a dehydrin that could possibly coat essential cellular proteins and protect them against conformational changes or agglomeration during freezing or dehydration (22). WCS120 was shown to protect LDH against freezing-induced denaturation by 100-fold compared to BSA (22). Thus, WCS120 could protect the integrity of adhesion molecules and prevent their detachment from the plasma membrane. Less is known about the function of WCS 19, which appears to play a role in the enhancement of freezing tolerance (36).
TaTIL is a plant lipocalin associated with the plasma membrane in freezing-tolerant plants. Functional studies revealed that this protein is involved in protecting plant cells against oxidative stress caused by freezing. TaTIL transcripts and proteins are upregulated during stresses such as cold and heat shock, which are known to induce membrane damage (8,15). Thus, Ta TIL is likely to protect the plasma membrane from cryopreservation-induced membrane damage and prevent the loss of adhesion molecules.
TaIRI-1 and TaIRI-2 (Triticum aestivum ice recrystallization inhibition) are low-temperature-responsive genes in wheat that are upregulated during cold acclimation in freezing-tolerant species (56). TaIRI-1 and TaIRI-2 are extracellular proteins that accumulate in the apoplast of freezing-tolerant plants, such as wheat, when they cold acclimate. The primary structure of TaIRI proteins shows a series of highly conserved repeated motifs with regularly spaced serine and threonine residues that may form hydrogen bonds with the ice surface and thus inhibit the growth of ice crystals in the extracellular space (45). This enables plants to avoid damage to the membrane and survive the dehydration caused by the formation of large ice crystals outside of cells. Based on these properties, it is likely that TaIRI-1 and TaIRI-2 exert their effects as ice recrystallization inhibition proteins outside of the hepatocytes, by avoiding the formation of large ice crystals in the extracellular medium, and thus protecting the integrity of the cell membrane from any mechanical damage. Furthermore, the extracellular ice is essential for the initiation of ice formation inside cells. Intracellular ice formation is known to inflict damage during cryopreservation of cultured hepatocytes and many other cell types (25). Therefore, the ability of the TaIRI protein to inhibit the growth of ice crystals at the extracellular level would also decrease intracellular ice formation, which is likely to contribute to improved survival and protection of the plasma membrane during the cryopreservation of hepatocytes. All of these wheat proteins appear to be expressed in response to membrane stresses (5), which suggests that they play a biological role in membrane biogenesis and repair under conditions of severe stress.
LDH leakage was previously investigated during a longer in vitro period of 4 days following cryopreservation of hepatocytes with crude wheat extracts (WPEs) (21). The crude WPEs maintained viability of cells at a similar level to that obtained in freshly isolated hepatocytes during 4 days of monolayer culture following cryopreservation. After 4 days, respective viabilities of 33.1%, 35.8%, and 17.6% were obtained in fresh, WPE-cryopreserved, and DMSO-cryopreserved hepatocytes. Furthermore, hepatocytes cryopreserved with crude WPEs maintained similar levels of hepatic functions, compared to fresh cells, for secretion of albumin, ammonia metabolism, and induction of CYP1A1 and CYP2B during 4 days in culture following cryopreservation (21). Crude WPEs, compared to DMSO, caused a marked improvement in viability and hepatospecific functions throughout a 4-day in vitro culture period, following cryopreservation of hepatocytes.
The findings in this study demonstrate the capacity of partially purified WPs and recombinant wheat proteins to improve the attachment efficiency of rat hepatocytes by protecting three key adhesion molecules (β1-integrin, β-catenin, and E-cadherin), which are known to be important for the structural and metabolic integrity of hepatocytes in cell culture. The determination of the mechanisms by which WPs protect these molecules could help to further optimize the procedure and obtain cryopreserved hepatocytes with similar attachment efficiencies to fresh cells. This promising technology offers a novel strategy for hepatocyte cryopreservation that could be useful for improving clinical hepatocyte transplantation, which is currently limited by the timely availability of hepatocytes. Future work will validate this technology in human hepatocytes.
Footnotes
Acknowledgments
The authors wish to thank Michel Marion for assistance with animal surgeries to obtain hepatocytes. Financial support was provided by grants from the Natural Sciences and Engineering Research Council of Canada (NSERC) awarded to F.S. and D.A.B.