
Abstract
As recent studies have shown that microglia play a key role in inflammation and immunological challenges as well as have broader roles in synaptic modulation in the brain, studies on psychiatric disorders have increasingly focused on microglia. Microglial abnormalities have consistently been observed in psychiatric postmortem brain studies, including altered microglial activation and changes in the protein and mRNA expression levels of microglial marker molecules, such as major histocompatibility complex, class II, DR (HLA-DR), complement receptor type 3 (CD11b), ionized calcium binding adaptor molecule 1 (IBA-1), macrosialin (CD68) and glucose transporter type 5 (GLUT5). Microglial abnormalities have also been observed in positron emission tomography (PET) studies. Recent advances in omics-based microglial gene expression profiling of psychiatric brains may elucidate microglial involvement in the pathogeneses of psychiatric disorders. In the present paper, we review the current status of research on expression profiling of microglia-relevant molecules in psychiatric postmortem and imaging studies and we discuss future research directions.
INTRODUCTION
Microglia are highly dynamic immune cells in the brain located within the brain parenchyma [1]. They differentiate from hematopoietic cells and then slowly turn over and are replenished by proliferation in the adult central nervous system (CNS) [2]. In the healthy brain, resting ramified microglia have neuroprotective and neurotrophic roles by dynamically interacting with other glial cells and neurons [3, 4]. In some pathological conditions, microglia activation may support brain inflammation, leading to neuronal cell death, which is mediated by the microglial release of pro-inflammatory cytokines and reactive oxygen and nitrogen species [11]. However, microglia that are activated in response to pathological events also have immune capacity or facilitate tissue repair [5]. Microglial activation, which is characterized by morphological changes, migration, proliferation, phagocytosis, production and the release of cytokines and chemokines, is induced by purinergic receptor agonists via increasing microglial intracellular calcium [6–10].
Recent evidence indicates that microglia regulate neuronal functions in the absence of immune challenge or inflammation. Microglia are required for synapses to develop during embryogenesis [11] and to regulate the number of functional synapses in the neonatal brain [12, 13]. Microglia also regulate adult neurogenesis, refine neuronal circuitry and drive oligodendrocyte differentiation during CNS remyelination [13–15]. Considering abnormalities in synaptic regulation, neuroplastic changes and neurogenesis have been implicated in the pathology of psychotic disorders [16–18]. Microglia-neuron crosstalk [19] may be a focus of future studies on psychiatric disorders.
Microglial activation has been observed in postmortem brains from patients with psychiatric disorders, including schizophrenia [20], mood disorders [21, 22], substance abuse [23], and suicidality [24]. However, detailed mechanisms of microglia involvement in the pathogeneses of psychiatric disorders have not been elucidated. Recent advances in omics technology may provide useful tools for studying the mechanism of microglia involvement. In this review, we summarize evidence supporting microglial dysfunction in psychiatric postmortem brain samples as well as discuss the current status and perspectives in omics strategies for characterizing the molecular mechanisms of microglial dysfunction in psychiatric brains.
MICROGLIAL ACTIVATION IN PSYCHIATRIC POSTMORTEM BRAIN SAMPLES (FINDINGS BASED ON CONVENTIONAL MICROGLIAL MARKER MOLECULES)
Microglia play a major role in neuronal migration, synapse formation, and neurotransmission control as well as in innate immunity and inflammation regulation within the developing and mature nervous system. In terms of the pathophysiology of neuropsychiatric disorders, previous studies have identified human microglial cell activation based on conventional microglia-specific markers, including major histocompatibility complex, class II, DR (HLA-DR), complement receptor type3 (CD11b, also known as integrin, alpha-M; ITGAM), ionized calcium binding adaptor molecule 1 (IBA1, also known as allograft inflammatory factor 1; AIF1), macrophage antigen CD68, (CD68, also known as macrosialin), and glucose transporter type 5 (GLUT5, also known as solute carrier family 2, member 5; SLC2A5) [20, 24–26], in psychiatric illnesses, which are described below. Based on these observations, theories on the neuropsychoimmunological mechanisms of neuropsychiatric disorders have been proposed, including immunological alterations that have been demonstrated in peripheral blood and cerebrospinal fluid samples from patients with psychiatric illnesses.
Schizophrenia
HLA-DR-immunostaining of postmortem brain tissue has suggested that there is microglial activation or increased microglial cellular density in schizophrenia. For example, HLA-DR-positive (i.e., activated) microglia are found in patients with late-onset schizophrenia (age older than 40 years) [20]. HLA-DR-positive microglia are increased in the frontal cortex [27–30] and temporal cortex [5, 6] of patients with chronic schizophrenia, while the brain region morphologies are not drastically altered in general [28]. HLA-DR-positive microglia in the posterior hippocampus are increased in paranoid schizophrenia patients compared with patients who have residual schizophrenia or with controls [31]. A significant increase was observed in the mean protein CD11b levels in the frontal cortex of patients with schizophrenia compared with controls [27]. CD11b is one of the most important phagocyte receptors for recognizing microbial pathogens [35], and it has been found in murine [36, 37] and human microglia [38]; furthermore, it is widely used as a marker for microglial identification. A qualitative assessment of IBA-1-stained microglial morphology demonstrated numerous activated microglial cells in the dorsolateral prefrontal cortex of schizophrenia patients [32]. IBA-1 is a calcium binding protein that is exclusively expressed in the microglia of murine and human brains [48, 49].
By contrast, several studies have provided evidence against the aforementioned proposed ideas. There were no statistically significant changes in the numbers of HLA-DR-positive cells in the anterior cingulate cortex [30, 33–35], dorsolateral prefrontal cortex [28, 34], hippocampus [31, 36], mediodorsal thalamus [33, 34] and dentate gyrus [34] from patients with schizophrenia. CD68-positive microglia were not altered in the mediodorsal thalamus [37] and dorsolateral prefrontal cortex [38] in patients with schizophrenia. CD68 is a member of a growing family of hematopoietic mucin-like molecules that are present in macrophages/microglia, and CD68 is a marker of cell proliferation [44]. Additionally, electron microscopy and morphometry analysis showed microglial cells are activated in both the PFC and visual cortex of schizophrenia patients [39].
Microglia-relevant molecule expression changes in the brains of patients with psychiatric disorders
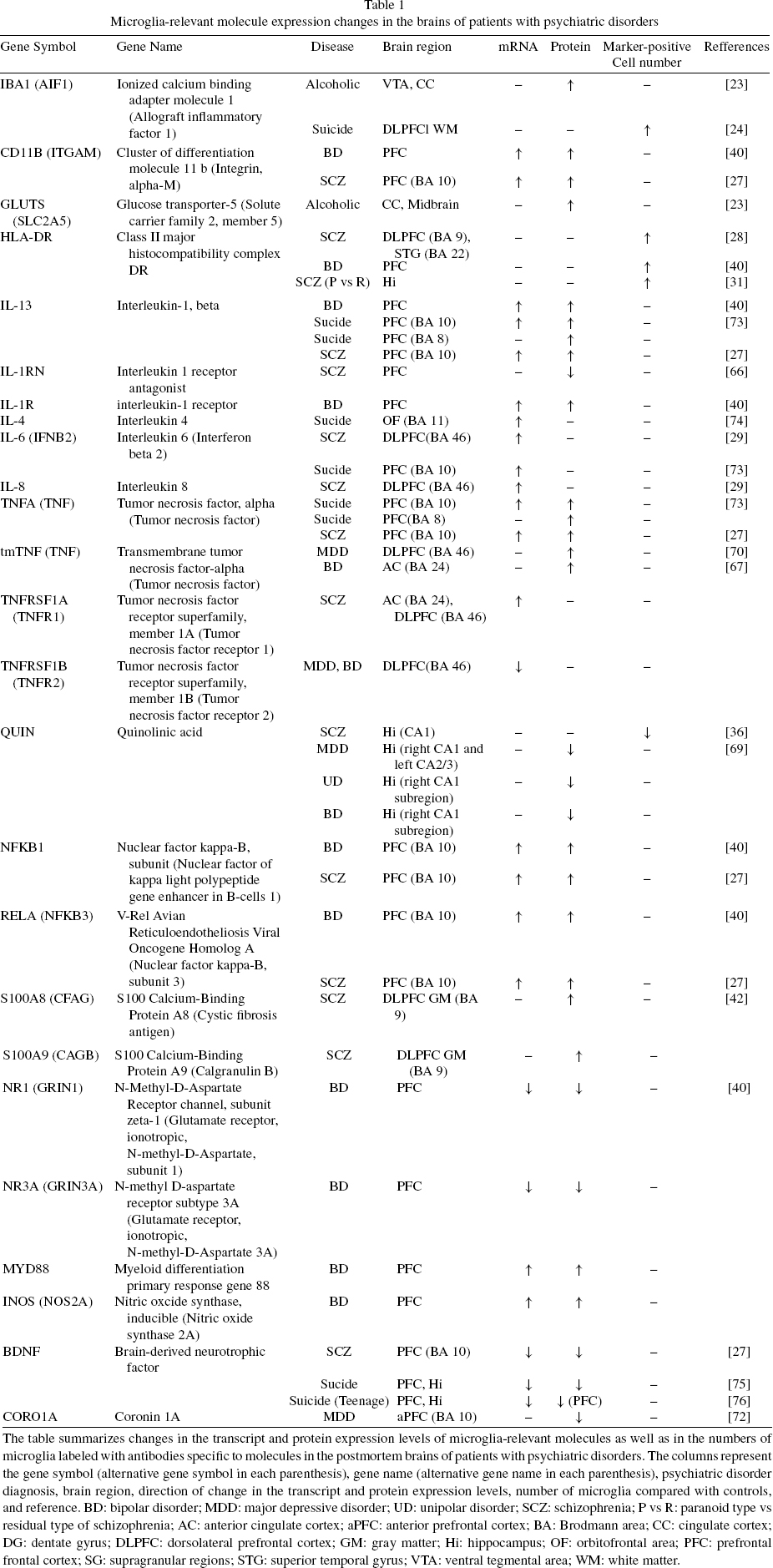
The table summarizes changes in the transcript and protein expression levels of microglia-relevantmolecules as well as in the numbers of microglia labeled with antibodies specific to molecules in the postmortem brains of patients with psychiatric disorders. The columns represent the gene symbol (alternative gene symbol in each parenthesis), gene name (alternative gene name in each parenthesis), psychiatric disorder diagnosis, brain region, direction of change in the transcript and protein expression levels, number of microglia compared with controls, and reference. BD: bipolar disorder; MDD: major depressive disorder; UD: unipolar disorder; SCZ: schizophrenia; P vs R: paranoid type vs residual type of schizophrenia; AC: anterior cingulate cortex; aPFC: anterior prefrontal cortex; BA: Brodmann area; CC: cingulate cortex; DG: dentate gyrus; DLPFC: dorsolateral prefrontal cortex; GM: gray matter; Hi: hippocampus; OF: orbitofrontal area; PFC: prefrontal frontal cortex; SG: supragranular regions; STG: superior temporal gyrus; VTA: ventral tegmental area; WM: white matter.
Bipolar disorder
Recent studies have shown that bipolar disorder involves microglial activation. Significant increases in the protein and mRNA levels of CD11b have been observed along with increased HLA-DR-positive microglia in the postmortem frontal cortices of bipolar disorder patients [40].
However, the density of IBA-1-stained microglia did not differ among bipolar disorder and control patients, which appears to contrast with the above study [32]. Another study also reported no significant differences in the HLA-DR-positive microglial density for bipolar I disorder patients compared with controls [33].
Depression
Because evidence supports microglial involvement in the pathogeneses of depression, Yirmiya, RN et al proposed that depression is a microglial disease [21]. A postmortem study demonstrated strong activation of HLA-DR-positive microglia in an old, affective disorder patient by immunohistochemistry [20]. Significant HLA-DR-positive microgliosis has been observed in the dorsolateral prefrontal cortex, anterior cingulate cortex and mediodorsal thalamus of suicidal patients with major depression, and a similar trend has been observed in the hippocampus [33]. Microgliosis is an accumulation of microglial cells as a reaction to parenchymal injury, which is accompanied with cellular morphological changes, such as the enlargement of cellular processes [41].
However, three other studies reported no differences between depressed patients and controls in IBA-1- and CD68- [24], HLA-DR- [33], and CD68- [42] positive microglial density.
Alcoholic patients
The immune system in the form of systemic and CNS inflammation has emerged in the field of alcohol abuse and alcoholism [43]; however, microglial involvement in alcoholic patients has not been well characterized. Thus far, only one research study showed that both IBA-1 and GluT5 were increased in the cingulate cortex of alcoholic brains relative to controls [23].
Suicidality
Recent postmortem studies reported that microgliosis, labeled by HLA-DR, was increased in two schizophrenic patients in the anterior cingulate cortex and mediodorsal nucleus of the thalamus; both of these patients, who had acute psychosis, committed suicide [34]. Additionally, microgliosis was observed in the dorsolateral prefrontal cortex and mediodorsal nucleus for suicide cases [33]. Additionally, one study indicated that the gene expression of IBA-1 and mRNA expression of CD45 were significantly increased in the dorsal anterior cingulate cortex for depressed patients who committed suicide [44]. However, the study reported no differences in the densities of resting or activated microglia in the dorsal or ventral prefrontal white matter, but the density of perivascular cells in dorsal white matter was higher between suicide and non-suicide cases, whereas the densities of activated microglial cells were higher ventrally than dorsally among suicide patients as well higher dorsally than ventrally among non-suicide cases [24]. Other studies also indicated that there are no significant microglial densities in the dorsolateral prefrontal cortex, anterior cingulate cortex, mediodorsal thalamus, and hippocampus of schizophrenic and depressive patients who committed suicide compared with non-suicidal cases [33]
Autism
Postmortem studies showed that the IBA-1-positive microglial somal volume in white matter and microglial cell density in gray matter were increased in a case with autism compared with control cases [45]. In a postmortem study of adult autistic patients, 2 of 8 autism brains displayed significant IBA-1-stained microglial activation in the amygdala [46]. HLA-DR-positive microglia were observed in the postmortem cerebellum of an autism patient [47]. The total densities of IBA-1-positive microglia were significantly greater in the cerebral cortex of autistic individuals, ranging from 3 to 22, compared with the control [48]. Additionally, the autism brains exhibited increased short-distance, microglia-neuron interactions (microglia-neuron spatial clustering) in the dorsolateral prefrontal cortex [49].
MICROGLIAL ACTIVATION IN PSYCHIATRIC DISORDERS BASED ON IMAGING STUDIES
Microglial activation in the brain of psychiatric patients, especially patients with schizophrenia, has been evaluated in several positron emission tomography (PET)-based neuroimaging studies utilizing microglia-specific radiotracers, such as (R)-[11C]PK11195. (R)-[11C]PK11195 binds selectively to the peripheral benzodiazepine receptor, which is also known as the mitochondrial 18 kDa translocator protein (TSPO). (R)-[11C]PK11195 binding was significantly higher in the hippocampus of schizophrenic patients (five patients were paranoid schizophrenia and 2 patients had brief psychotic disorder not otherwise specified) [50]. (R)-[11C]PK11195 was also significantly higher in the total gray matter of schizophrenic patients during acute psychotic episodes that were early in the disease course [51]. Another PET study on a novel positron emission tomography (PET) ligand, [11C]DAA1106, instead indicated that there were no significant microglial alterations in any of the brain regions between patients with chronic schizophrenia and control subjects. There was no significant difference between [11C]DAA1106 binding of the cortical regions of normal controls and patients with schizophrenia, whereas the patients had a positive correlation between cortical [11C]DAA1106 binding and positive symptom scores [52].
PET-based evaluation of microglial abnormalities has rarely been used for other psychiatric disorders. Thus far, a study on bipolar disorder reported a significantly increased [11C]-(R)-PK11195 binding potential in the right hippocampus of disease patients compared with controls. Although the same trend was observed in the left hippocampus, this difference was not statistically significant [53]. To the best of our knowledge, there have been no reports on PET-based evaluation of microglial abnormalities in patients with alcoholism. PET imaging revealed microglial activation in autistic subjects compared to control subjects. Significantly higher (R)-[11C]PK11195 levels were observed in the cerebellum, midbrain, pons, fusiform gyri, and anterior cingulate and orbitofrontal cortices from young adult subjects with autistic spectrum disorder compared with control subjects [54].
MICROGLIAL GENE EXPRESSION ABNORMALITIES IN PSYCHIATRIC POSTMORTEM BRAIN SAMPLES AND POTENTIAL MICROGLIAL INVOLVEMENT
In addition to the aforementioned conventional microglial marker molecules, microglia exert their functions via transcriptional regulations of various genes. For example, microglia play major roles in neuroimmunological and neuroinflammatory processes via transcriptional regulation of pro- or anti-inflammatory cytokines and neurotrophic factors, which also play important roles in the development of psychiatric disorders [55]. Microglia can differentiate into M1 and M2 phenotypes during these processes [56]. The M1 phenotype is associated with enhanced antigen presentation and secretion of pro-inflammatory cytokines, such as interleukin-1 (IL-1), IL-6, IL-8 and tumor necrosis factor-α (TNFα). The M1 phenotype is also associated with nuclear factor-kappa B (NF-κB), a transcription factor that plays a major role in regulating pro-inflammatory genes as well as in upregulating myeloid differentiation primary response gene 88 (MyD88), which facilitates NF-κB activities in combination with toll-like receptors [57, 58]. The M1 phenotype is also characterized by activated nitric oxide synthase (iNOS) expression [5, 60]. In contrast, the M2 phenotype secretes anti-inflammatory cytokines IL-4, IL-5 and IL-10 as well as bone morphogenetic protein 7 (BMP-7), which suppresses the expression levels of pro-inflammatory cytokines. The M2 phenotype is also characterized by the expression of trophic factors, including brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF), glial cell-derived neurotrophic factor (GDNF), insulin-like growth factor gene 1 (IGF-1), transforming growth factor β (TGF-β), and hepatocyte growth factor (HGF) [5].
In addition to the molecules related to M1 and M2 microglia, S100A8- and S100A9-positive microglia are observed in brain samples following cerebral infarction and other neurological disorders [72, 73]. S100A8 and S100A9 belong to the macrophage-related protein-100 family of calcium binding proteins, which inhibit pro-inflammatory functions [61–63]. QUIN, an endogenous modulator with agonistic properties against N-methyl-D-aspartate, which is produced by microglia, plays a role in the pathogenesis of several major neurological diseases [64, 65]. Evidence supports that microglial abnormalities are accompanied by inflammatory cytokines, which may be responsible for the abnormalities in neural development or psychiatric disorder functionality. In the brain, only microglia, not neurons or astrocytes, produce detectable QUIN levels [65].
Schizophrenia
In frontal cortex samples from SZ patients, the IL-1β protein level was significantly increased [27], whereas IL-1 receptor antagonist (IL-1RA) protein, which is mostly enriched in the prefrontal cortex, was significantly reduced in the schizophrenic brain samples [66]. The IL-6 and IL-8 mRNA expression levels were also significantly upregulated in the dorsolateral prefrontal cortices from schizophrenic patients [29]. The TNFα mRNA and protein levels were also significantly increased in frontal cortex samples from schizophrenic patients [27]. Additionally, the TNF receptor type I (TNFR1) mRNA level was significantly increased in both the anterior cingulate and dorsolateral prefrontal cortices of patients with schizophrenia [67]. The protein expression levels of M1-related nuclear transcription factor subunits, NF-κBp50 and NF-κBp65, were significantly increased in the frontal cortices of schizophrenia patients. In contrast, the protein and mRNA levels of the M2-related growth factor BDNF were significantly reduced in the frontal cortices of schizophrenia patients [27]. The protein expression levels of other microglial marker chemokines, S100A8/A9, were significantly higher in the dorsolateral prefrontal cortices of patients with schizophrenia [42]. Additionally, there were significantly more QUIN-immunoreactive microglial cells in the CA1 hippocampal subregions of schizophrenia patients. However, no significant changes in quinolinic acid (QUIN)-immunoreactive microglial cells were observed in the hippocampus CA2/3 and dentate gyrus regions of patients with schizophrenia [36].
Bipolar disorder
A postmortem study demonstrated significantly elevated mRNA and protein levels of IL-1β, IL-1R and MyD88 in the frontal cortices of bipolar disorders patients [40]. The tmTNFα levels were increased in the postmortem anterior cingulate of bipolar disorder patients [67]. The TNFR2 mRNA level was significantly decreased in the dorsolateral prefrontal cortices of bipolar and major depressive disorder patients [67]. The nuclear protein and mRNA levels of microglial NFKB1 (p50) and NF-κBp65 [40] were significantly increased in the postmortem frontal cortices of bipolar disorder patients. Recombinant N-methyl-d-aspartate receptor subtypes NR-1A and NR-3A were expressed in the microglia [68] and the corresponding mRNA and protein levels were significantly decreased in the frontal cortices of bipolar disorder patients compared to controls [40]. M1 Microglia phenotype marker iNOS mRNA and protein levels were significantly increased in the frontal cortices of bipolar disorder patients compared with controls [40]. Additionally, QUIN staining was reduced in both unipolar and bipolar disorders in the right CA1 field as well as in depressed patients in the right CA1 and left CA2/3 of the hippocampus [69].
Depression
Several postmortem studies reported that the transmembrane TNF (tmTNF) protein level was significantly increased in the dorsolateral prefrontal cortex but not increased in the anterior cingulate cortex of major depression disorder patients. There was no change in the soluble TNF (sTNF) levels in either brain region [70]. Patients with bipolar disorder and major depressive disorder had significant reductions in the TNFR2 mRNA expression in the dorsolateral prefrontal cortex without a reduction in the anterior cingulate cortex levels [67]. Additionally, microglia-specific actin-binding protein coronin-1a (CORO1A) [71] was decreased in the anterior prefrontal cortices of patients with major depressive disorder [72].
Suicidality
Of the pro-inflammatory cytokines, the IL-1β, IL-6 and TNFα mRNA and protein levels were significantly higher in the prefrontal cortices of suicide victims compared with healthy control brains [73]. Analysis of covariance revealed no significant difference in the IL-1β, IL-6 and TNFα expression levels in the orbitofrontal cortex between controls and suicidal patients, whereas there were significant differences in the orbitofrontal area in the expression levels of an anti-inflammatory cytokine, IL-4, between suicide victims and controls. Higher IL-4 expression levels were observed in women [74]. The BDNF protein and mRNA levels were significantly reduced in both the prefrontal cortex and hippocampus of suicide subjects [75]. The BDNF protein expression level was significantly decreased in the prefrontal cortex of teenage suicide victims, whereas no change was observed in the hippocampus [76]. Additionally, the mRNA expression of both BDNF and its receptor, tropomyosin receptor kinase B (TrkB), were significantly decreased in the prefrontal cortex and hippocampus of teenage suicide victims [76].
COMPREHENSIVE GENE EXPRESSION PROFILES OF MICROGLIA AND GENE FUNCTION
Thus far, a number of gene expression profiles of postmortem brain tissues from patients with psychiatric disorders have been reported for schizophrenia, bipolar disorder, depression, and autism. Among them, the cell type specificities for neurons [77–81] and oligodendrocytes [82, 83] have thoroughly been discussed. However, microglia-related gene expression profiles have received little attention. In addition to the aforementioned conventional microglial markers, cytokines and growth factors, various other microglia-specific molecules may be involved in neuroinflammatory aspects of the pathogeneses of psychiatric disorders. Evaluation of the comprehensive gene expression profiles of microglia may help clarify the role of microglia in psychiatric disorders.
To better understand how microglial gene expression profiles differ between schizophrenic patients and controls, we selected tentative microglial gene expression profiles based on the gene expression profile of isolated microglia from mice and then compared them with gene expression profiles that are specific to the major neuropsychiatric diseases. Using an Illumina BeadChip-based gene expression profile of microglia from C57BL/6 murine brains [84], 6,319 transcripts with signal intensities greater than 500 were considered to have high levels of detectable transcripts in microglia. However, based on Illumina BeadChip-based C57BL/6 prefrontal cortex microarray data [85] from the PubMed GEO DataSet, 22,024 transcripts had signal intensities of less than 500, which is considered low in the prefrontal cortex. The overlapped 2,024 transcripts (1,814 genes) were defined as microglia-dominant transcripts (genes), which are highly expressed in microglia and relatively less expressed in prefrontal cortex samples.
The list of microglia-dominant transcripts was compared with the gene expression profiles of postmortem prefrontal cortex samples from schizophrenia, bipolar disorder, and major depression patients provided by Stanley Foundation Brain Collection to estimate how microglia-related transcripts are related to the pathogeneses of psychiatric disorders. The gene expression profiles of each psychiatric disorder were compared with controls, and altered genes were considered increased or suppressed with a fold change of >1.2 or <0.833, respectively. Among whole detectable genes (8,583), 1,319, 1,636 and 1,312 genes were increased in schizophrenia, bipolar disorder and depression, respectively, while 1,263, 1,253 and 1,206 genes were suppressed in each psychiatric disorder, respectively. Of these, 391 and 278 genes were commonly increased and suppressed, respectively, in all three diseases. Among the above increased genes in schizophrenia, bipolar disorder and depression, 134 (10.2%), 177 (10.9%) and 158 (12.0%) genes overlapped with the 1,814 microglia-dominant genes. Likewise, among the aforementioned increased genes in schizophrenia, bipolar disorder and depression, 138 (10.9%), 113 (9.0%) and 102 (8.5%) genes overlapped with the microglia-dominant genes. Among 391 commonly increased genes and 278 commonly suppressed genes in all three diseases, 39 (10.0%) and 23 (8.3%) genes overlapped with the microglia-dominant genes. The proportions of altered microglia-specific genes in each psychiatric disorder were 8–12%, which is reasonably comparable to the microglia (10–15%) composition of all brain cells [86]. These preliminary data suggest that evaluating postmortem brain omics data, including gene expression profiles, while focusing on microglia may provide useful insights that have otherwise been masked by the cellular heterogeneity of the tissue characteristics.
PERSPECTIVES
As summarized in previous descriptions, multiple studies based on the microglial marker-based evaluation of postmortem brains have indicated that microglia are generally activated in the brains of patients with schizophrenia, bipolar disorder, and major depression, especially in suicidal cases. These findings were supported by multiple PET studies. Further, multiple studies have indicated that M1-type marker expression levels in microglia were increased whereas M2-type markers were decreased in general in the postmortem brains of patients with psychiatric disorders. Although some studies do not support microglial activation in psychiatric postmortem brains, few studies demonstrate the opposite direction of change. Considering the heterogeneity even in a single psychiatric disorder, such as schizophrenia, bipolar disorder, and major depressive disorder, the previous findings may indicate that microglia are activated in certain types and processes of psychiatric disorders.
In addition to the heterogeneity of the disorder, several features need to be considered in postmortem brain studies of psychiatric disorders. For example, a previous study indicated that the postmortem interval was significantly correlated with the ramified cell numbers in the anterior cingulate cortex and dorsolateral prefrontal cortices as well as with the ameboid cell density in the hippocampus [34]. Notably, the effect of postmortem interval on the gene expression profile in a postmortem brain is generally low if the interval is shorter than 48 hours. However, the agonal condition and brain tissue pH more drastically impact the gene expression profile [87]. Evaluations of the postmortem interval, agonal factor and tissue pH need to be considered when assessing the molecular expression profiles of microglia in postmortem brain tissue samples from psychiatric patients.
Autoimmune processes and inflammatory component alterations have been implicated in the pathogenesis of psychiatric disorders based on postmortem brain analyses. Detailed molecular mechanisms of microglial involvement in the pathogeneses of psychiatric disorders have yet to be elucidated. On the other hand, substantial gene expression profiling of psychiatric postmortem brains has been conducted, and some expression profiling studies have focused on the cellular specificities of neurons and oligodendrocytes. However, microglia-specific gene expression profiles have received little attention. Application of omics technologies to microglial gene expression profiling of psychiatric brains may elucidate distinct mechanisms of microglial involvement in the pathogeneses of psychiatric disorders.
Footnotes
ACKNOWLEDGMENTS
This work was partially supported by a grant-in-aid for scientific research on innovative areas (No. 24116007) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan, Health and Labour Sciences Research Grants research on psychiatric and neurological diseases and mental health (H19-kokoro-ippan-001), and an Intramural Research Grant (No. 21-9) for Neurological and Psychiatric Disorders from the National Center of Neurology and Psychiatry. All authors declare no biomedical financial interests or potential conflicts of interest.