
Abstract
Recently, the concept of the brain-gut-microbiota (BGM) axis disturbances in the pathogenesis of Alzheimer’s disease (AD) has been receiving growing attention. At the same time, accumulating data revealing complex interplay between bile acids (BAs), gut microbiota, and host metabolism have shed new light on a potential impact of BAs on the BGM axis. The crosstalk between BAs and gut microbiota is based on reciprocal interactions since microbiota determines BA metabolism, while BAs affect gut microbiota composition. Secondary BAs as microbe-derived neuroactive molecules may affect each of three main routes through which interactions within the BGM axis occur including neural, immune, and neuroendocrine pathways. BAs participate in the regulation of multiple gut-derived molecule release since their receptors are expressed on various cells. The presence of BAs and their receptors in the brain implies a direct effect of BAs on the regulation of neurological functions. Experimental and clinical data confirm that disturbances in BA signaling are present in the course of AD. Disturbed ratio of primary to secondary BAs as well as alterations in BA concertation in serum and brain samples have been reported. An age-related shift in the gut microbiota composition associated with its decreased diversity and stability observed in AD patients may significantly affect BA metabolism and signaling. Given recent evidence on BA neuroprotective and anti-inflammatory effects, new therapeutic targets have been explored including gut microbiota modulation by probiotics and dietary interventions, ursodeoxycholic acid supplementation, and use of BA receptor agonists.
INTRODUCTION
Recently, the concept of the brain-gut-microbiota (BGM) axis disturbances in the pathogenesis of Alzheimer’s disease (AD) has been receiving growing attention [1, 2]. Accumulating data indicate that amyloid-β deposits constituting the characteristic feature of AD may occur at different levels of the axis including both the central and enteric nervous systems [2]. Importantly, gut microbiota is also the source of significant amount of amyloids [3]. Moreover, numerous alterations in the gut microbiota composition and function in AD patients have been reported [1–6]. The mechanisms involved in regulation of the BGM axis through which gut microbes can communicate to the brain include neural, immune, and neuroendocrine pathways [7, 8]. Additionally, microbial metabolites such as serotonin, dopamine, short-chain fatty acids, as well as secondary bile acids (BAs) may serve as direct neurotransmitters or neuromodulators [8, 9].
Over recent years there has been also growing understanding of the complex role of BAs in regulating host physiology and host-microbiota interactions [10–15]. In the past, BAs as naturally acting detergents were considered to play a cardinal role in the digestion, micelle formation, and absorption of fat and fat-soluble vitamins. Currently, they are recognized as important signaling molecules actively participating in the regulation of numerous metabolic processes and significantly affecting gastrointestinal motor, sensory and secretory functions, as well as intestinal barrier permeability and regulation of inflammatory response [16]. Given close interactions between BAs and gut microbiota, they emerge as potential modulators of the BGM axis that may have direct impact on neurodegeneration. In fact, numerous experimental and clinical studies confirm the BA involvement in the pathogenesis of AD [17–23]. This review is aimed at highlighting most recent progress in the field with respect to the BA impact on neural, immune, and neuroendocrine interactions within the BGM axis.
BILE ACID SYNTHESIS, METABOLISM, AND ENTEROHEPATIC CIRCULATION
While cholesterol metabolism is thought to play a pivotal role in the pathogenesis of AD, BAs are the products of cholesterol clearance [24]. Importantly, many cholesterol metabolism-related genes (e.g., ABCA7, ABCG1, SORLI, BINI, CLU, and PICALM) are among the main AD susceptibility loci identified by genome-wide association studies [25, 26]. Three different pathways of cholesterol clearance may be distinguished (reviewed in detail in [24]). The classic pathway, which is the main pathway of BA synthesis in the liver, begins with the conversion of cholesterol to 7α-hydroxycholesterol by CYP7A1, which is subsequently hydroxylated by sterol 12α-hydroxylase (CYP8B1) or sterol 27-hydroxylase (CYP27A1) to form the primary BAs: cholic acid (CA) and chenodeoxycholic acids (CDCA), respectively [27, 28]. The alternative BA synthesis pathway is mediated by CYP27A1, which unlike CYP7A1, is expressed in the mitochondria of most cells [28]. The second (acidic) pathway is responsible for about 5–10%of BA synthesis in humans, and about 25%in rodents [28]. The alternative pathway produces regulatory oxysterols [16]. The third pathway, called the neural one, is responsible for the majority of cholesterol turnover in the central nervous system (CNS) [29]. The brain clears excess cholesterol via CYP46A1 that catalyzes the conversion of cholesterol to 24S-hydroxycholesterol, which being more polar than cholesterol can more easily pass the blood-brain barrier (BBB) to exit the brain and pass into the bloodstream [28]. It has been shown that in AD patients, there is significantly decreased expression of cholesterol-24 hydroxylase in neurons [30].
Primary BAs (CA and CDCA) after conjugation in the liver with glycine or taurine are secreted into the bile and transported to the gut lumen when they are metabolized by bacteria. Gut microbiota initiates bile acid metabolism via a critical first step catalyzed by bile salt hydrolases (BSHs) [31]. These enzymes hydrolyze and deconjugate glycine or taurine from the sterol core of the primary BAs. Deconjugated BAs can subsequently undergo a variety of microbiota-encoded transformations (i.e., 7 α-dehydroxylation, dehydrogenation, and epimerization) that generate secondary BAs, which have widespread effects on the host and resident microbiota [31–33]. CA is converter to deoxycholic acid (DCA), while CDCA is converted to lithocholic acid (LCA) and ursodeoxycholic acids (UDCA) [31, 33].
In the terminal ileum about 95%of BAs are reabsorbed primarily through active transport by the apical sodium-dependent BA transporter (ASBT) [34]. Subsequently, they are released into the portal vein for return to the liver where they are conjugated to produce their glycine and taurine forms [35]. BAs, which are not reabsorbed in the ileum, enter the colon, and serve as substrates for microbial metabolism and undergo biotransformation to secondary BAs [36]. BAs regulate their own synthesis and transport via the nuclear farnesoid receptor (FXR). After binding to the ileal FXR, BAs induce expression of fibroblast growth factor FGF19 (FGF15 in mice), which travels via the portal bloodstream and binds to hepatocyte receptors repressing BA synthesis by inhibiting the rate-limiting enzyme CYP7A1 via negative feedback [37].
BILE ACID SIGNALING
A wide variety of BA characteristics is associated with their lipophilicity or hydrophilicity, but also with their ability to bind numerous receptors as well as activate ion channels [38]. While unconjugated BAs may be able to easily cross the plasma membrane, conjugated BAs might cross the plasma membrane using transporters. Nuclear receptors activated by BAs are the FXR, pregnane X receptor (PXR), vitamin D receptor (VDR), liver X receptor (LXR), and glucocorticoid receptor (GR). Whereas G protein-coupled receptors (GPCR) on cell surface activated by BAs include Takeda G-protein receptor 5 (TGR5), sphingosine-1-phosphate receptor 2 (S1PR2), muscarinic receptors (M2 and M3), and formyl peptide receptor (FPR). Moreover, BAs are able to activate ion channels such as epithelial Na+ channel (ENaC), and large-conductance Ca2+- and voltage-activated K+ (BK) channels. Noteworthy, all these nuclear and membrane receptors as well as bile acid-sensitive ion channels (BASIC) are expressed in the brain [38].
BAs vary in their ability to activate nuclear receptors and GPCRs. FXR is activated most potently by the primary BA –CDCA, while TGR5 receptor is mainly activated by secondary BAs (LCA and DCA) [12]. Additionally, UDCA (secondary BA in humans) and murine primary BAs (TαMCA and TβMCA) act as FXR antagonists. The ability to activate two major BA receptors occurs in the following order –for FXR: CDCA > LCA = DCA > CA and for TGR5: LCA > DCA > CDCA > CA [16, 39]. FXR is involved in BA synthesis, transport, and enterohepatic circulation as well as multiple glucose and lipid metabolic processes [40]. FXR is also a suppressor of inflammation in the liver and intestines and enhances epithelial barrier properties [41]. TGR5 being expressed on neuroendocrine cells induces glucagon-like peptide (GLP-1) release and plays an important role in glucose homeostasis [40, 42]. Moreover, TGR5 is also expressed in brown adipose tissue, muscles, and immune cells [43]. BA signaling through membrane-bound or nuclear receptors as well as neuromodulatory effects of BAs are accurately summarized in a graphic form in the review by McMillin and DeMorrow [27].
INTESTINAL CROSSTALK BETWEEN BILE ACIDS AND GUT MICROBIOTA
The crosstalk between BAs and gut microbiota is based on reciprocal interactions since microbiota determines BA metabolism, while BAs affect gut microbiota composition [12, 45]. Gut microbes directly involved in BA biotransformation control the composition of BA pool, which in turn has a profound impact on the size of BA pool [11]. Comparing germ-free rodents to rodents with typical microbiota significant differences in the BA pool size have been confirmed [46]. Gut microbiota is also able to modulate BA synthesis impacting the enzyme reactions. For example, antibiotic treatment associated with microbiota depletion inhibits the expression of CYP7A1 [47]. The expression of CYP7A1 and CYP27A1 was also found to be reduced in germ-free mice compared to normal mice [48]. Changes in the BA pool composition associated with altered ratio of primary to secondary BAs may directly affect their signaling due to different affinities to their various receptors [16, 39].
On the other hand, BAs participate in the regulation of the gut microbiota composition by direct antimicrobial effects and indirect effects through FXR-induced antimicrobial peptide synthesis in the small intestine [49, 50]. In general, a decreased BA level in the gut lumen favors gram-negative bacteria containing lipopolysaccharides (LPS) in their outer membranes, whereas an increase in BA concentration appears to favor gram-positive bacteria such as Firmicutes that are able to turn primary BAs via 7α-hydroxylation to more toxic secondary BAs [11]. In a mouse model, biliary duct ligation resulted in overgrowth of aerobic and anaerobic bacteria in the ileum and cecum, but that overgrowth could be blocked by the FXR agonist [50]. Another clinically relevant example of BA impact on the gut microbiota ecosystem is infection with Clostridioides difficile. It has been shown that in a healthy intestinal environment microbiota-derived secondary BAs inhibit the growth of this pathogen. However, antibiotic-induced dysbiosis may result in a rapid shift in the BA pool leading to decreased secondary BA level that promotes the growth of C. difficile [48].
Clostridia are among a narrow group of commensal bacteria able to perform 7α-dehydroxylation constituting a crucial reaction in secondary BA origination. Noteworthy, these spore-forming bacteria have also important impact on tryptophan metabolism in the gastrointestinal tract [51]. Golubeva et al. [52] have suggested some association between microbiota-related alterations in tryptophan metabolism and inadequate bacterial conversion of BAs that are both likely to contribute to intestinal dysmotility and impaired gut barrier function in a mouse model of autism.
Another interesting aspect of the connection between human and microbiota BA metabolism is related to probiotic BSH-containing strains like Lactobacillus and Bifidobacterium that not only participate in deconjugation of BAs but may also accumulate primary and secondary BAs in their cytoplasm either spontaneously or following intracellular BA deconjugation [53, 54]. Accumulation of primary BAs in the cytoplasm of probiotic bacteria results in their reduced conversion to secondary BAs in the colon which may have some implications in chronic inflammation [53, 55].
BILE ACIDS IN THE BRAIN
The presence of twenty BAs and their receptors has been demonstrated in the brain [38, 56]. Among them are unconjugated BAs (CA, CDC, DCA, and UDCA) and conjugated BAs (GDCA, TCA, TDCA, and TUDCA) which are related to brain physiology and pathophysiology [57]. Mano et al. [56] found that CDCA composes about 95%of total BA composition with CA and DCA being about 2-3%in the rat brain. It has been also suggested that since BA level in the brain under normal conditions is approximately 10-fold higher than circulating level, they may be actually synthesized locally [56]. It is consistent with the observation that BAs may serve as potent neuroactive steroids [58]. In fact, CDCA was found to be about 10 times more abundant than the prototypical neurosteroid pregnenolone [56]. However, BAs can be also transported into the brain from the peripheral circulation. While conjugated BAs require transporters to cross the BBB, unconjugated and lipophilic BAs can cross the BBB by passive diffusion [38, 59]. In addition, BAs in the bloodstream may affect the BBB permeability as it has been shown that phosphorylation of occludin—one of the tight junction proteins—may be increased by CDCA and DCA via ras-related C3 botulinum toxin substrate 1 (RAC1) [60]. On the other hand, UDCA and its glycine-conjugated form (GUDCA) exert a protective effect on brain endothelial cells by reducing apoptosis [61, 62].
Noteworthy, both amino acids, glycine and taurine, conjugated with BAs in the liver have neurotransmitter properties [63, 64]. Glycine is an inhibitory neurotransmitter in the CNS serving as a co-agonist along with glutamate for N-methyl-D-aspartate (NMDA) receptors [63]. Taurine demonstrates functions of a neurotransmitter, neuromodulator, and neuroprotectant, and acts as a trophic factor in the CNS development. The neurotransmitter properties of taurine are illustrated by its ability to elicit neuronal hyperpolarization [64].
The role of BAs as neurosteroids is related to their affinity to receptors for several neurotransmitters [38, 58]. TCA, GDCA, and TDCA activate muscarinic acetylcholine receptors (M2 and M3) that are known to play an important role in cognitive function, memory, and learning [65]. Moreover, CDCA, DCA, and CA block both γ-aminobutyric acid type A (GABAA) receptors and NMDA receptors [66]. The NMDA receptor activation is also important for learning and memory [67], while GABAA activation leads to the inhibition of neurotransmission [68]. All these observations imply an important role of BAs in the regulation of neurological functions. Additionally, there are accumulating data supporting neuroprotective effect of some BAs such as TUDCA [38]. Their neuroprotective action may be associated with inhibition of amyloid-β accumulation, mitochondrial damage, and apoptosis. It has been also suggested that TUDCA may induce neurogenesis in adults, but the mechanism of that action remains to be elucidated [38].
INTERACTIONS BETWEEN BILE ACIDS AND THE BRAIN-GUT-MICROBIOTA AXIS
Disturbances along the BGM axis may significantly contribute to the pathogenesis of AD [1, 5]. Alterations in the gut microbiota composition induce increased permeability of the gut barrier and immune activation leading to systemic inflammation, which, in turn, may impair the BBB and promote neuroinflammation, neural injury, and ultimately neurodegeneration associated with the formation of amyloid-β plaques and neurofibrillary tangles [1, 69–71]. Bacterial amyloids through molecular mimicry may elicit cross-seeding of misfolding and cause priming of the immune system, consequently enhancing immune response to endogenous production of neuronal amyloid in the brain [72]. Recognition of bacterial amyloid by the human TLR2 receptor (toll-like receptor 2) results in the release of pro-inflammatory cytokines such as Il-1β and Il-6 produced by activated bone marrow macrophages as well as Il-11 and Il-17A produced by activated T-lymphocytes [73]. These cytokines are able to cross the BBB and induce the production of reactive oxygen species, activation of the TLR2/1 and NF-κB signaling pathways, which are directly related to neuroinflammation and neurodegeneration [74, 75]. Also bacterial LPS can bind with microglial cell receptors (TLR2, TLR4, and/or CD14) [76, 77]. In the brain of animals exposed to curli-producing bacteria enhanced microgliosis and astrogliosis together with increased expression of TLR2, Il-6, and TNF were found [31]. In addition, microglia activation further impairs phagocytosis leading to the accumulation of Aβ42 [78, 79].
Recently, BAs possessing neuroactive properties have been emerging as a direct communication channel between the gut microbiome and the brain, since gut microbiota is directly involved in BA synthesis and metabolism affecting their pool composition and properties [80]. That, in turn, may affect BAs ability to bind and signal through their receptors.
The contribution of gut microbiota to the pathogenesis of AD is well depicted in animal models as well as clinical studies (reviewed in detail in [1]) [1, 81]. Regarding interactions between BAs and gut microbiota, an important observation in AD patients concerns a decrease in Clostridium and Bifidobacterium [81]. Both genera have BSHs which allow BAs to become deconjugated and less toxic to the microbiota. Since BSHs are the only enzymes responsible for the pivotal deconjugation reaction, their activity serves as a gatekeeper to subsequent BA transformations [82]. BA deconjugation enables also subsequent metabolism of BAs via 7α-dehydroxylation (Clostridium spp.) to secondary BAs which act as ligands for FXR [21]. The BA pool, influenced by microbial BSH activity, shapes in turn the microbiome composition and function [83]. The significant association between increased gut microbiota-derived secondary BAs both conjugated and unconjugated and changes in AD markers such as amyloid and tau proteins have been recently reported in AD patients by Nho et al. [19]. In that study serum-based BA metabolites were investigated and several BA ratios that reflect BA conversion by gut microbiota were assessed. Among numerous reported alterations in BA profiles, higher GDCA:CA levels were associated with greater global cortical amyloid deposition, while two conjugated secondary BA (GLCA and TLCA) were correlated with higher p-tau concertation in the cerebrospinal fluid (CSF) [19]. This observation supports the role of the gut microbiota transformation of BAs as a potential contributing factor in the pathogenesis and/or progression of AD.
Moreover, a growing body of evidence indicates that BAs may serve as important modulators of each of three main routes through which interactions within the BGM axis occur including neural, immune, and neuroendocrine pathways that will be discussed below (Fig. 1). A wide spectrum of the BA effects may be exerted via their receptors found throughout the body. Among BA target cells are neurons, immune cells, and enteroendocrine cells [21].
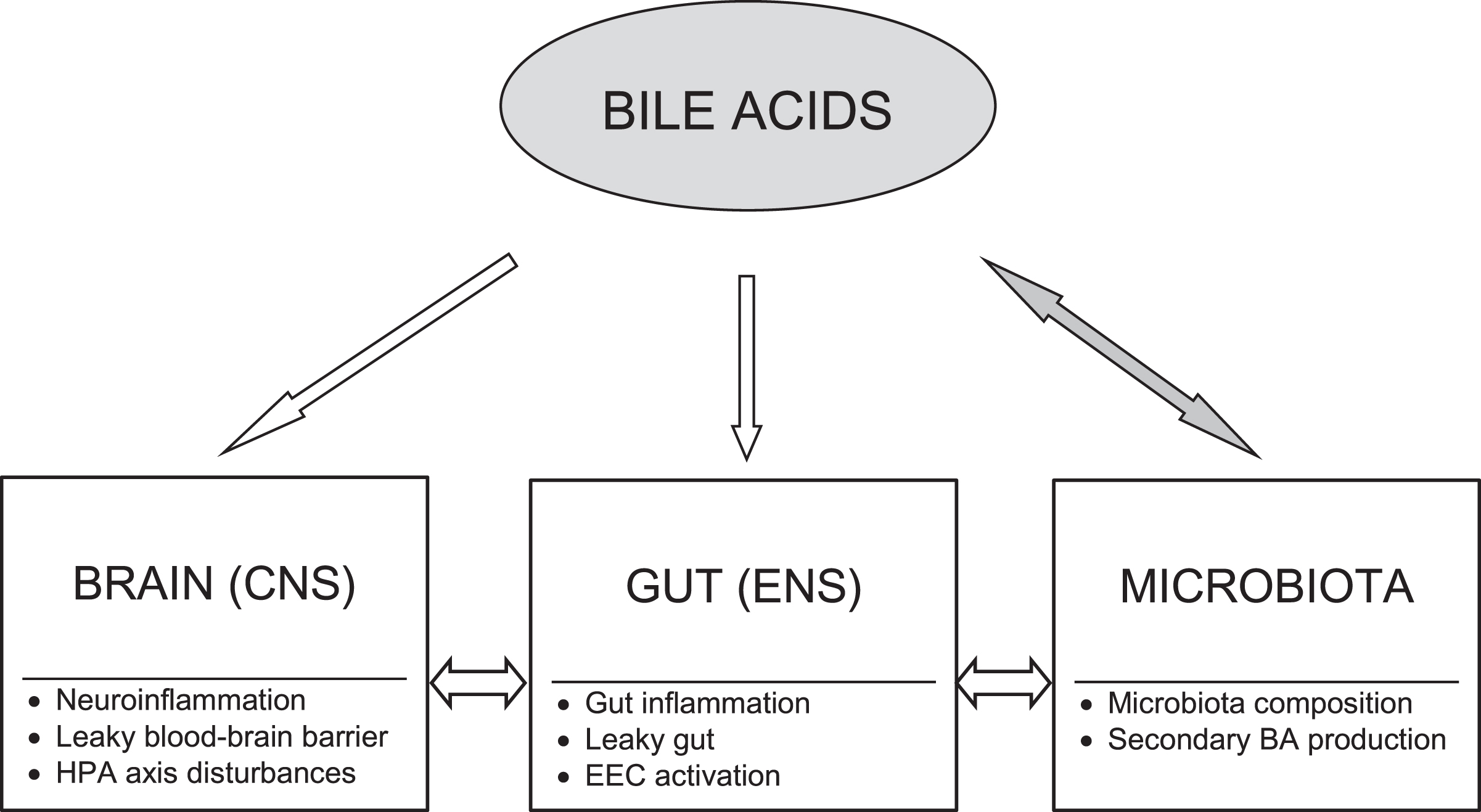
Bile acid modulatory effects on the brain-gut-microbiota axis in Alzheimer’s disease. Bile acids (BAs) may act as modulators of each level of the brain-gut-microbiota axis. There are bidirectional interactions between gut microbiota and BAs. The BA pool composition depends on gut microbiota producing secondary BAs, whereas BAs significantly influence microbiota composition. The primary to secondary BA ratio determines their signaling due to diversified binding affinities of BAs to various receptors. BAs may modify activation of both peripheral and central neural and immune cells as well as affect gut barrier and blood-brain barrier permeability. Moreover, BAs through their receptors exert neuroendocrine effects via enteroendocrine cells (EEC) and the hypothalamic-pituitary-adrenal (HPA) axis. CNS, central nervous system; ENS, enteric nervous system.
Neural pathways
BA signaling via neural pathways may occur directly (via BA receptors present in the brain and enteric nervous system) or indirectly (via the FXR-FGF15/19 axis). The most studied BA receptors are found in the brain [84]. The function of FXR and TGR5 in the brain is still unclear, but it has been reported that FXR knockout mice showed disturbed neurotransmitter system and altered neurobehavior [85]. The results from studies with FXR knockout mice confirmed changes in serotoninergic as well as glutamatergic, GABAergic, norepinephrinergic neurotransmission in either the hippocampus or cerebellum [85]. Additionally, TGR5 being expressed on neurons, glial and microglial cells can be activated by numerous neurosteroids [86, 87]. In neurodegenerative processes such as AD, BA receptors may significantly affect decline in cognitive function by affinity to muscarinic receptors as well as GABAA and NMDA receptors [66].
Among other BA receptors expressed in the brain, special attention in the context of AD is focused on pregnane X receptor (PXR), vitamin D receptor (VDR), and glucocorticoid receptor (GR) [38]. PXR as xenobiotic nuclear receptor activating cytochrome P450 can dispose toxins, for example at the BBB [88]. Vitamin D in the brain can act as neurosteroid and its beneficial effect in AD has been confirmed [89, 90]. The GR activation in the hypothalamus by BAs may suppress the hypothalamic-pituitary-adrenal (HPA) axis, which is the key arm of neuroendocrine interactions along the BGM axis [91]. Interestingly, UDCA can bind to and translocate GR [91].
Moreover, TGR5 receptors have been also identified in the enteric nervous system where their activation result in increased colonic motility, which may lead to BA-induced diarrhea [92]. Noteworthy, BA effects on the gastrointestinal tract may show regional differences between various segments of the tract, since for example activation of TGR5 on gastric smooth muscle results in delayed gastric emptying [92].
The indirect neural signaling pathway of BAs to the CNS involves FGF15/19. FGF19 can cross the BBB and bind to FGFRs that are also expressed in the brain, specifically in the hypothalamus, medial habenular nucleus and dorsal vagal complex [93]. FGF19 binds to FGFR4 but a more solid bond is created when β-Klotho, the FGF19 cofactor, is bound to FGF19-FGFR4 complex [61, 94].
Immune pathways
Although amyloid-β deposition and tau pathology have been considered as the main features of AD, accumulating data show that disturbances in immune responses resulting in neuroinflammation considerably contribute to AD pathogenesis and may be even a trigger of the pathology [95–97]. The effect of BA action is determined by various factors such as concentration of BAs, their physicochemical properties, interactions with the gut microbiota and target cells [98]. Activation of BA receptors in immune cells results in several regulatory functions that are mainly inhibitory in nature [99]. Recent findings have been pointing at a crucial role of FXR in the modulation of inflammatory response and intestinal barrier integrity [41, 100]. The results of both in vivo and in vitro studies have demonstrated that on the one hand the activation of FXR exerts anti-inflammatory effect by reducing the production of pro-inflammatory cytokines such as TNF-α, IFN-γ, interleukin-1β (IL-1β), IL-2, and IL-6 [101]. On the other hand, inflammation reduces FXR expression [102]. It has been suggested that the reduced FXR transcriptional activity upon TNF-α/IL-1β stimulation is mediated by NF-κB [41]. In fact, FXR and TGR5 knockout mice develop spontaneous intestinal inflammation [103, 104]. While in a mouse model of ulcerative colitis it has been demonstrated that experimental colitis may disturb BA synthesis by the negative feedback signaling within the FXR-FGF19 axis [105].
Additionally, both nuclear and membrane BA receptors are highly represented in immune cells such as intestinal and liver macrophages, dendritic cells, and natural killer T cells [99]. Activation of these receptors shifts the macrophage phenotype toward anti-inflammatory one (M2) with an upregulation of Il-10 and a downregulation of the pro-inflammatory cytokines including Il-6 and IFN-γ. In turn, Il-10 increases the production of Treg cells reducing the recruitment of monocytes from blood to the colon. The TGR5 signaling involves cAMP-protein kinase A (PKA)-mediated inhibition of NF-κB, and FXR signaling leads to repression of NF-κB-responsive elements. BA stimulation of both pathways blunt NF-κB-dependent gene expression. Regarding action on dendritic cells, BAs downregulate the production of TNF-α and IL-12 [99].
Another important aspect of the BGM interactions and immune response concerns the integrity of intestinal and blood-brain barriers that may also be affected by BAs [106]. As mentioned above, BAs increase phosphorylation of occludin that may result in disruption of these barriers [60]. Additionally, TGR5 activated by BAs can activate the cJun NH2-terminal kinase (JNK) pathway leading to degradation of tight junction proteins [107, 108]. Intestinal barrier integrity is also regulated by an interplay between gut symbionts and PXR through a pathway which involves luminal sensing and signaling by TLR4 [109]. Direct interactions between gut microbiota, gut immune system activation, and intestinal permeability constitute a critically important element of the BGM axis [1]. In fact, increased biomarkers of gut permeability and inflammation in AD has been recently reported [110]. An age-related shift in the gut microbiota composition associated with its decreased diversity and stability observed in AD patients may significantly affect BA metabolism and signaling [4, 5]. Additionally, dysbiosis-related intestinal inflammation and increased gut permeability are associated with an increase in proinflammatory cytokines and bacteria-derived products in the circulation resulting in systemic inflammation. Subsequently, due to concomitant BBB impairment, systemic inflammation may induce neuroinflammation observed in AD [1].
Interesting findings concern sex-dependent differences in aging-associated inflammation and dysregulation of systemic BA homeostasis [111–114]. Recently, sexual dimorphisms in human immune system aging have been reported pointing to higher innate and pro-inflammatory activity and lower adaptive activity in older men compared to older women [115]. The rate of BA synthesis and BA pool composition are also sexually dimorphic [116]. For example, in animal studies female mice have a larger total BA pool than males. Moreover, females excrete less fecal BA and catabolize less cholesterol via BA production than males [117]. Additionally, age-related hormonal changes are implicated in the differential production of BA in females [118]. Ma et al. [119] demonstrated that aging in mice has a greater impact on changing hepatic BA profiles in females, but gut BA profiles in males. Moreover, gut microbiota remodeling by co-housing with younger mice partially rescued the above changes in old mice [119]. Even more importantly, BAs and FXR as well as gut microbiota alterations are implicated in the sex-related differences in metabolic disorders, many of which constitute risk factors for AD [120]. Given the role of BA homeostasis disturbances in neurodegeneration, those sex-dependent differences in BA profiles, especially in the elderly, could potentially explain female predominance in AD. Indeed, recently, Varma et al. [23] have suggested that cholesterol catabolism and BA synthesis may impact dementia progression through sex-specific effects on signaling pathways in the brain.
Neuroendocrine pathways
BAs as neurosteroids may exert a direct impact on the hypothalamus and the HPA axis. Suppression of the HPA axis has been shown to occur during cholestatic liver injury [69]. Furthermore, McMillin et al. [91] have demonstrated that in a model of cholestasis, serum BAs gain entry into the brain via a leaky BBB and that hypothalamic BA content is increased.
Currently, another neuroendocrine pathway in which BAs may be involved is emerging as BA receptors are expressed on enteroendocrine cells (EECs) forming the diffuse neuroendocrine system [61, 122]. These interactions may have direct clinical implications in AD. A particular group of EECs, L-cells, responsible for the production of glucagon-like peptide-1 (GLP-1), are predominantly located in the terminal ileum and the colon [123]. Secondary BAs are powerful stimulants of GLP-1 release through the activation of TGR5 receptors located at the basolateral membrane of L-cells [124]. Additionally, L-cells may release GLP-1 in response to other endocrine or neural signals, not involving BA-induced TGR5 activation [125]. Although BA receptors have not been found on the vagus nerve so far, vagal afferent fibers express GLP-1 receptors and are therefore responsive to local GLP-1 level [126]. The nodose ganglion projects to the nucleus of the solitary tract (NST in the hindbrain is connected with other brain regions including the hypothalamus) [126, 127]. That points at the vagal-brainstem-hypothalamic pathway for the signal transmission of GLP-1 from the gastrointestinal tract to the CNS, which may be indirectly impacted by BAs [61].
Lately, GLP-1 and connections between its release and BA signaling are attracting attention as a possible link between metabolic and brain impairment [128, 129]. GLP-1 not only plays a key regulatory role in glucose metabolism stimulating insulin secretion, inhibiting glucagon secretion, decreasing food intake, reducing appetite, and inducing weight reduction, but affects also neurological and cognitive functions [129]. Hyperinsulinemia and insulin resistance, which are known as pathophysiological features of type 2 diabetes and obesity, have also been demonstrated as risk factors for AD [130]. Importantly, GLP-1 receptor agonists are used in the treatment for diabetes and obesity [131]. Moreover, modulation of GLP-1 activity can influence amyloid-β aggregation in AD and GLP-1 receptor agonists reduce hippocampal neurodegeneration [132]. There is also growing evidence on neuroprotective effects of these agonists in animal models of neurodegenerative diseases, regardless of diabetes [128]. Physiological consequences of BA impact on the TGR5-dependent release of GLP-1 in patients with obesity and/or on high fat diet remain to be elucidated. Nonetheless, it has been recently highlighted that BA signaling mediated by gut microbiota may contribute to obesity susceptibility [40]. Therefore, modulation of BAs could be a promising therapeutic target in obesity and obesity-related diseases including AD.
It has been also shown that secondary BAs acting through TGR5 may activate intestinal stem cells and epithelial regeneration [133] as well as induce L-cell differentiation and serotonin signaling [134]. Of note, serotonin in 95%stored in the EECs, is a key mediator of the neuroendocrine pathway within the BGM axis [135, 136].
BILE ACID DISTURBANCES IN EXPERIMENTAL AND CLINICAL SAMPLES OF ALZHEIMER’S DISEASE
Substantial BA perturbations have been reported in AD. Pan et al. [17] performed metabolomic profiling of BAs in both experimental and clinical samples of AD. Quantification of BAs in brain tissue extract and plasma was performed using a BA kit enabling simultaneous evaluation of 22 BAs. Human brain samples in that study were obtained from postmortem confirmed AD cases, while mouse brain samples from APP/PS1 male mice. Pan et al. [17] found higher plasma CA level in APP/PS1 mice compared to wild type mice. Additionally, in the brains of APP/PS1 mice higher LCA and lower tauromuricholic (TMCA), β-muricholic, and Ω-muricholic acids as well as lower TCA and TUDCA were demonstrated [17]. Intriguingly, CDCA was not detected in mouse brain [17], contrary to an earlier study performed in rats in which CDCA level in rat brain was approximately 30 times higher than those in the serum [56]. The beneficial or detrimental effects of BAs are related to their hydrophobicity or hydrophilicity. For example, hydrophobic BAs, like CDCA can damage biological membranes [137], while hydrophilic BAs, like UDCA and TUDCA, are able to inhibit apoptosis stabilizing mitochondrial membrane [138, 139].
In the above-mentioned study, Pan et al. [17] reported the following disturbances in BA profiles in clinical samples. In AD patients compared to age-matched control subjects lower plasma CA and lower brain TCA levels were found [17]. In contrast, the plasma level of LCA, a secondary BA known for its toxicity, was significantly higher in AD patients than in controls [140]. In another study the ratio of secondary to primary BAs (DCA/CA) in serum has been shown to be significantly higher in AD patients [18]. Higher level of microbiota-derived secondary conjugated BAs (GDCA, GLCA, and TLCA) were significantly associated with worse cognitive function [18]. Furthermore, in a follow-up study those changes were also correlated with alterations in CSF markers of the disease and with brain imaging changes [18]. Altogether, it suggests that the effects of gut-directed dysregulation of primary versus secondary BAs are not limited to the periphery but may also affect BA homeostasis and signaling in the human brain. Additionally, it supports the significance of the BGM axis interactions in the pathogenesis of AD in the context of BA disturbances [38]. Nevertheless, the casual relationship of these interactions needs to be further elucidated.
Interestingly, serum BA profile in AD has been shown to be associated with AD-related structural and functional neuroimaging as well as CSF biomarkers including amyloid-β, t-tau and p-tau [19]. For example, one conjugated primary BA (GCDCA) and two microbiota-derived conjugated secondary BAs (GLCA and TLCA) were associated with higher t-tau and p-tau values, reduced glucose metabolism, and larger structural brain atrophy [19]. Very recently, Baloni et al. [20], based on the brain metabolic network analysis using AD patient-derived postmortem transcriptomics and metabolomics, have revealed differential usage of alternative BA pathway for cholesterol clearance in AD. Additionally, metabolomics analysis of postmortem brain samples identified higher ratio of GCDCA/CA and secondary BAs like DCA, LCA, TDCA, CDCA, and GDCA in AD compared to controls suggesting the association of those BAs with cognitive decline in AD [20]. The presence of secondary BAs in the brain metabolomics strongly points at the role of gut microbiome in AD.
Major limitations in BA research are differences in BA profiles and metabolism among species making it challenging to translate data from animal models to the clinic [45]. Noteworthy, not only significant differences in BA profiles have been reported between human and rodents, but also between mice and rats. These differences may result in discrepancies observed in experimental vs clinical studies, like for example alteration in CA plasma level that was lower in AD patients but higher in a mouse model of AD [17]. Another example concerns UDCA which is considered to be the primary BA in bears, beavers, and mice, but the secondary, microbiota-derived BA in humans [141]. Furthermore, in mice one of the main primary BAs is muricholic acids which is not present in humans and which can act as an antagonist of FXR/FGF15 axis [142]. In humans primary BAs act as potent FXR agonists [12]. Given these discrepancies caution in interpreting translational research is required.
Other important limitations in BA research in patients with AD are related to dietary factors, concomitant disorders such as obesity and diabetes, and medications including statins commonly used in the elderly that may directly or indirectly, via alterations in microbiota composition, affect BA synthesis and metabolisms [111]. On the one hand dietary patterns have impact on BA metabolism, while on the other hand food intake is partially regulated via BA-related changes in FXR and/or TGR5 signaling modulating the secretion of gastrointestinal hormones [143]. For example, in the study comparing BA profiles in vegans versus omnivores, a higher dietary fiber and lower fat intake was correlated with lower fecal BA concentrations [144]. Significant changes in dietary patterns leading to malnutrition are common in the elderly, and the lack of fiber and healthy fat intake can induce a state of systemic low-grade chronic inflammation [145]. Malnutrition itself is a risk factor for cognitive decline [146]. Eating and swallowing problems, which are present in about 80%of AD patients [147], along with the behavioral and psychological symptoms of dementia, can strongly affect nutritional status [148]. Moreover, an association between food intake and oral health problems common in the elderly may also contribute to neurodegeneration [149].
THERAPEUTIC IMPLICATIONS AND PERSPECTIVES
Given recent evidence on BA neuroprotective and anti-inflammatory effects their therapeutic potential has been intensively studied. In particular, better understanding of a critical role of BA-microbiota interactions in the regulation of metabolism and BGM axis has directed growing interest in microbiota-based approaches to modulate BA metabolism and associated conditions including neurodegeneration [15]. Numerous reports support the beneficial effect of probiotics by enhancing intestinal epithelial integrity, protecting against barrier disruption, reducing pro-inflammatory response, and inhibiting initiation or propagation of neuroinflammation and neurodegeneration [150, 151]. Additionally, probiotic administration may increase deconjugation of BAs and their fecal exertion in association with increased BSH activity in the gut and general modification of the microbiota composition [152, 153]. While supplementation with probiotics such as Lactobacillus and Bifidobacterium has been shown to improve significantly Mini-Mental State Examination scores in AD patients [154], administration of a BSH-positive Lactobacillus strain remarkably also reduces cholesterol levels in subjects with hypercholesterolemia [155].
Dysbiosis and dysregulated BA synthesis are implicated in Western diet-induced systemic inflammation, microglial activation, and reduced neuroplasticity [156]. Therefore, alternative approaches to manage diseases linked to BA dysmetabolism include dietary interventions aimed at modulating BA metabolism through balancing the microbiota [157]. Food-based therapies affect the gut microbiota composition or may have a direct influence on neuronal functioning in both the enteric and central nervous systems [158, 159]. The MIND diet, which is a hybrid of the Mediterranean and DASH (Dietary Approaches to Stop Hypertension) diets, was designed to emphasize nutrients that have been associated with dementia prevention and to eliminate products associated with increased risk of dementia [160]. Healthy diet is characterized by a high intake of fiber, probiotics, antioxidants, and omega-3 polyunsaturated fatty acids, as well as low intake of animal-derived proteins, saturated fats, and refined sugar. Such a diet has been shown to inhibit inflammatory response, reduce insulin resistance, and decrease the risk of neurocognitive impairment and eventually the risk of AD [145, 161]. The use of prebiotics as food-compounds inducing the growth and/or activity of beneficial microorganisms is of particular interest. Among most studied prebiotics are nondigestible fibers such as inulin and fructooligosaccharides and dietary polyphenols [145]. A summary of the effects of probiotics and synbiotics (combination of pro- and prebiotics) in AD patients and animal models of AD is presented in an extensive review by Arora et al. [145]. Probiotic, prebiotic, and polyphenol-rich interventions have been reported to exert beneficial effects on fasting lipid profiles, with changes in the gut microbiota composition associated with lipid regulation [162].
Additionally, the effect of fecal microbiota transplantation (FMT) is being investigated in many animal models exploring pathogenetic mechanisms of neurodegenerative disorders [163]. Recently, the first case report on rapid improvement in AD symptoms following FMT for C. difficile infection has been published [164]. Intriguingly, the main FMT action might be related to restoring secondary BA synthesis [165] that would confirm the role of BA in the modulation of BGM interactions in AD.
Other options to modulate BA signaling include supplementation of neuroprotective BAs such as UDCA or TUDCA [166, 167]. Most central effects of TUDCA are dependent on the activation of TGR5, S1PR2, and α5β1-Integrin, which have already been identified in the brain [167]. Indeed, TUDCA has been shown to increase cell survival and inhibit neuronal apoptosis in a number of experimental models of neurodegenerative diseases such as AD, Huntington’s disease, and Parkinson’s disease [168, 169]. Furthermore, in animal model of acute neuroinflammation TUDCA exerts “triple” anti-inflammatory effect including reduction in glial cell activation, microglial cell migratory capacity, and expression of chemoattractants and vascular adhesion proteins [170]. TUDCA also reduced the accumulation of amyloid-β in the hippocampus and frontal cortex as well as rescued memory deficits in APP/PS1 transgenic mice [171, 172]. The TUDCA effects in experimental models of AD are summarized in Table 1. Although there is still no strong evidence on benefits in a clinical setting there are several ongoing registered trials with TUDCA and UDCA in AD, Parkinson’s disease, Huntington’s disease, amyotrophic lateral sclerosis, and progressive multiple sclerosis [167].
TUDCA effects in experimental models of Alzheimer’s diseases
ApoE, apolipoprotein E; AβPP, amyloid-β protein precursor; Bax, Bcl-2-associated X protein; ER, endoplasmic reticulum; JNK, c-Jun N-terminal kinase; LPS, lipopolysaccharide; MAP2, microtubule-associated protein 2; mEPSCs, miniature excitatory postsynaptic currents; PS1, presenilin 1; PSD-95, postsynaptic density protein 95.
A promising new direction of BA research would be to target BA receptors and indirect pathways of BA signaling to the brain mediated by FXR-FGF19 and TGR5-GLP-1 axes [61, 183]. FXR agonists targeting the gut-liver axis may reduce gut inflammation and preserve the intestinal barrier [41] as well as improve metabolic disorders associated with increased risk for AD [184]. Furthermore, GLP-1 receptor agonists have been reported to exert therapeutic effect in diabetes, obesity, and cognitive decline [128].
BA co-metabolism of both host and gut microbiota, and the diversified binding affinities to BA receptors among primary, secondary, free, and conjugated BAs result in the immense complexity of BA signaling [47, 119]. The advent of new analytical tools and strategies such as metabolomics and metagenomics is expected to bring a rapid progress in gaining insight in these complex BA-gut microbiota-host interactions [47]. It should enable to elucidate not only therapeutic but also diagnostic potential of BAs, which may serve as disease biomarkers. Noteworthy, intensive efforts focusing on unraveling BA signaling in AD should take into account multiple confounding factors such as gut dysbiosis, diet, sex, age, concomitant disorders, and medications. Most importantly, the causal relationship between BA disturbances and the AD pathology needs to be verified.
CONCLUSIONS
Given a direct reciprocal crosstalk between gut microbiota and BAs, they are emerging as key signaling molecules within the BGM axis. Moreover, since bacteria-derived secondary BAs can not only affect the composition and function of gut microbiota but also can modulate host physiology, they have been recognized as interkingdom signaling molecules [11, 45]. BAs have even been called a communication channel tuning the BGM interactions [80]. The observed disturbances in BA homeostasis in experimental and clinical samples of AD support the association between BA and the AD pathology as well as the significance of the BGM axis [32]. It indicates some potential therapeutic targets including gut microbiota modification, UDCA supplementation or administration of BA receptor agonists. The application of multi-omics techniques in unraveling the molecular mechanisms that underly BA-gut microbiota-host interactions should help to provide the basis for interventional studies.
DISCLOSURE STATEMENT
The author’s disclosure is available online (https://www.j-alz.com/manuscript-disclosures/21-0608r1).