
Abstract
Background:
Spatial navigation is the ability to estimate one’s position on the basis of environmental and self-motion cues. Spatial memory is the cognitive substrate underlying navigation and relies on two different reference frames: egocentric and allocentric. These spatial frames are prone to decline with aging and impairment is even more pronounced in Alzheimer’s disease (AD) or in mild cognitive impairment (MCI).
Objective:
To conduct a systematic review of experimental studies investigating which MCI population and tasks are used to evaluate spatial memory and how allocentric and egocentric deficits are impaired in MCI after navigation.
Methods:
PRISMA and PICO guidelines were applied to carry out the systematic search. Down and Black checklist was used to assess methodological quality.
Results:
Our results showed that amnestic MCI and AD pathology are the most investigated typologies; both egocentric and allocentric memory are impaired in MCI individuals, and MCI due to AD biomarkers has specific encoding and retrieval impairments; secondly, spatial navigation is principally investigated with the hidden goal task (virtual and real-world version), and among studies involving virtual reality, the privileged setting consists of non-immersive technology; thirdly, despite subtle differences, real-world and virtual versions showed good overlap for the assessment of MCI spatial memory.
Conclusion:
Considering that MCI is a subclinical entity with potential risk for conversion to dementia, investigating spatial memory deficits with navigation tasks might be crucial to make accurate diagnosis and rehabilitation.
Keywords
INTRODUCTION
Exploring, orienting, and navigating would not be possible without a spatial representation of the environment. Spatial navigation is the ability to estimate one’s position on the basis of both static environmental (allothetic) and dynamic (idiothetic) self-motion cues [1]. While the former is based on visual perception and comprises stable objects, such as landmarks, the latter is based on vestibular, proprioceptive, interoceptive, and motor commands and comprises body-based (i.e., path integration and spatial updating) self-motion [1–3]. Both cues are integrated to build up spatial representations that determine one’s position and orientation within the environment [3, 4]. In this regard, both cognitive and bodily inputs yield to a successful navigation [4, 5]. Navigation relies on spatial memory that encompasses processes such as encoding, storing, recognizing and recalling spatial information about the environment, the objects and agents within it [6–8]. Considering that spatial memory is the cognitive substrate underlying navigation, its study is crucial to comprehend and investigate mechanisms and processing that determine our behavior within the environment. Spatial memory depends on two different reference representations that organize the information coming from the environment: egocentric and allocentric frames [9, 10]. On one hand, the egocentric frame refers to the individual location in the environment and it is based on subject-to-object relations. In fact, this memory subsystem employs the self as the reference frame in order to create a body-centered representation. On the other hand, the allocentric frame refers to object-to-object relations independently of individual’s location in the environment. This memory subsystem employs the object and/or the environment as the reference frame in order to create a world-centered representation. To successfully navigate within the environment, it is necessary to flexibly switch and combine these two reference frames according to environmental needs [1]. Studies underlying neural mechanism of spatial memory date back to the second half of the 19th century. Egocentric and allocentric frames exhibit separate neural circuits mediating different spatial strategies [11–13]. Concerning allocentric frame, O’Keefe and Dostrovsky [14] found that specific hippocampal cells, called place cells, create a cognitive map of the space, firing when a rat occupies a particular location in the environment. These findings were consistent also in humans [15, 16]. Other important cells in the medial temporal lobe, grid cells [17], head direction cells [18], and boundary cells [19], accomplish a successful cognitive map and navigation, by providing information to the hippocampus. Other neural substrates which are typically responsible for allocentric processing are parahippocampal and retrosplenial cortex (RSC) [20, 21]. With respect to egocentric frame, parietal area and dorsal striatum are responsible of the egocentric visual representations of the space, which depend both on visuospatial processing and on individual’s position, actions and decisions in proximity to discrete landmarks [10, 23]. The ability to switch from one frame to the other involves the posterior cingulate cortex (PCC) and the RSC instead [13, 25]. Furthermore, egocentric and allocentric frames share some common networks like the bilateral fronto-temporal regions [26–28]. Hence, spatial memory encoding is a bottom-up process that starts in the parietal ‘window’ (a visual egocentric representation of the space) and goes through medial parietal and temporal regions for storage and vice versa for spatial memory recall [13], where frontal regions play a broader role in memory processing, spatial attention and egocentric/allocentric strategy selection [28, 29].
Spatial memory decline in mild cognitive impairment
Navigation and spatial memory are prone to deterioration with normal aging [30, 31]. Frames switching abilities and allocentric difficulties have been reported in healthy older people [32]. Indeed, older individuals tend to prefer egocentric compared to allocentric navigation strategies [33]. In addition, self-motion appears to be compromised in aging [31, 34]. These impairments reflect neurophysiological alternations that occur in regions which are crucial for spatial cognition such as medial temporal areas, basal ganglia, and prefrontal cortex [35]. Deterioration is even more pronounced in neurodegenerative diseases like Alzheimer’s disease (AD). In fact, due to the gradual degeneration of specific brain areas (i.e., entorhinal, medial temporal, medial parietal cortices) critical in AD diagnosis [36–38], egocentric and allocentric memory are impaired [39]. Specifically, Serino and colleagues [40] reviewed egocentric and allocentric deficits in AD patients reporting a prevalence of allocentric abilities impairments. Similar deficits [39] have been found also in prodromal AD (amnestic mild cognitive impairment; aMCI) with hippocampal amnesia [41] or in MCI due to AD biomarkers [42].
MCI is considered a transitional period between normal and pathological cognitive aging [43, 44]. It is characterized by a subtle general decline in cognitive functions, usually affecting first memory domain [45]. The objective decline essentially preserves daily activities and is insufficient to constitute dementia [46]. It has been mainly linked to AD progression [45]. Indeed, prodromic signs of AD can be detected in aMCI by means of: neuropsychological tests, such as the Free and Cued Selective Reminding Test (FCSRT) [47], which can detect hippocampal (H) and non-hippocampal (NH) amnesia according to Dubois and Albert criteria [41]; biomarkers of the National Institute on Aging and Alzheimer’s Association Research Framework [38] or Albert’s criteria for MCI due to AD [42] to identify neuropathological signs of AD in MCI; genetic profiling according to APOE ɛ4 [48] non-carriers (defined with - or homozygous –/–in the papers included in the Results section) or ɛ4 carriers (+), more specifically with heterozygous (+/–) and homozygous (+/+) carriers, or according to TOMM40 poly-T length polymorphisms [49] distinguishing individuals with short (S), very long (VL) homozygous and heterozygous (S/VL) profiles at risk for AD. Finally, mixed forms of AD pathology and cerebrovascular disease (i.e., striatal lacunar lesions/white matter hyperintensities [WMH]) can be evaluated according to Fazekas’s criteria [50].
However, the heterogeneity of this clinical condition has prevented to reach a general consensus about its diagnosis. In fact MCI could differ for type of cognitive domain impaired (aMCI versus non amnestic; naMCI), the quantity of domains compromised (single versus multiple domains; abbreviated sd and md hereafter) and the etiology (e.g., neurodegenerative, cerebrovascular, depression/neuropsychiatric) [45, 52]. With respect to spatial navigation, several studies have reported impairments in both allocentric and egocentric frame and in the ability to switch between them [40, 53–55] in aMCI. These findings have been supported by neuropathological evidence of neurodegeneration of hippocampal areas and RSC, regions involved in spatial representations [39, 57]. Preliminary findings have been found also in other neurodegenerative diseases, such as dementia with Lewy body [58], frontotemporal dementia [59], Parkinson’s disease [60], and in vascular cognitive impairment [61, 62]. Additionally, subjective navigation impairment has been reported also in naMCI and in individuals with subjective cognitive decline [63].
Virtual reality for assessment of spatial memory
Virtual reality (VR) is a new technology that has been increasingly employed to study human spatial memory since it allows to provide navigation mechanisms analogous to those activated in real-life navigation [64, 65]. Its potential relies on the capability to adapt and tailor environments according to patient’s and task’s needs, for example by changing or eliminating landmarks or pathways in spatial memory tasks [29, 66]. Further, via VR it is possible to manipulate user’s spatial frame from egocentric to allocentric or vice versa [64, 67–69]. Indeed, several studies have employed VR to investigate spatial memory, reproducing classical paradigms used to study navigation such as the hidden goal task (HGT), a human analogue of the Morris water maze [70]. In the VR HGT participants, by using the mouse, are asked to retrieve goal position in a circular map (the arena) by using both starting position (egocentric) and distal cues (allocentric) or just one of these. The relation between goal, starting point and distal cues was showed before each trial but then according to the subtest (allocentric, egocentric or mixed) cues were removed/left. Additionally, after 30 min allocentric (delayed) memory was tested. The procedure was the same for the Blue Velvet Arena (the real-world version; BVA) but this time participants walked in a 2.9 m dark arena with luminous goal locations and egocentric/allocentric cues. Other alternative tasks achieved promising diagnostic results with VR [29, 71–74], and with real-space versus virtual versions [75–77] or only real-world tasks [53, 78–80]. Moreover, VR is one of the preferred tools to study spatial memory since it allows the user to actively navigate within the virtual environments, enhancing spatial and episodic performances [74, 81]. In fact, active navigation in virtual environments involves both bodily and cognitive activities [5] by including motor commands, proprioceptive information, vestibular information, decision-making, allocation of attentional resources, and manipulation of spatial information [4]. Conversely, passive navigation implies only visual information [4].
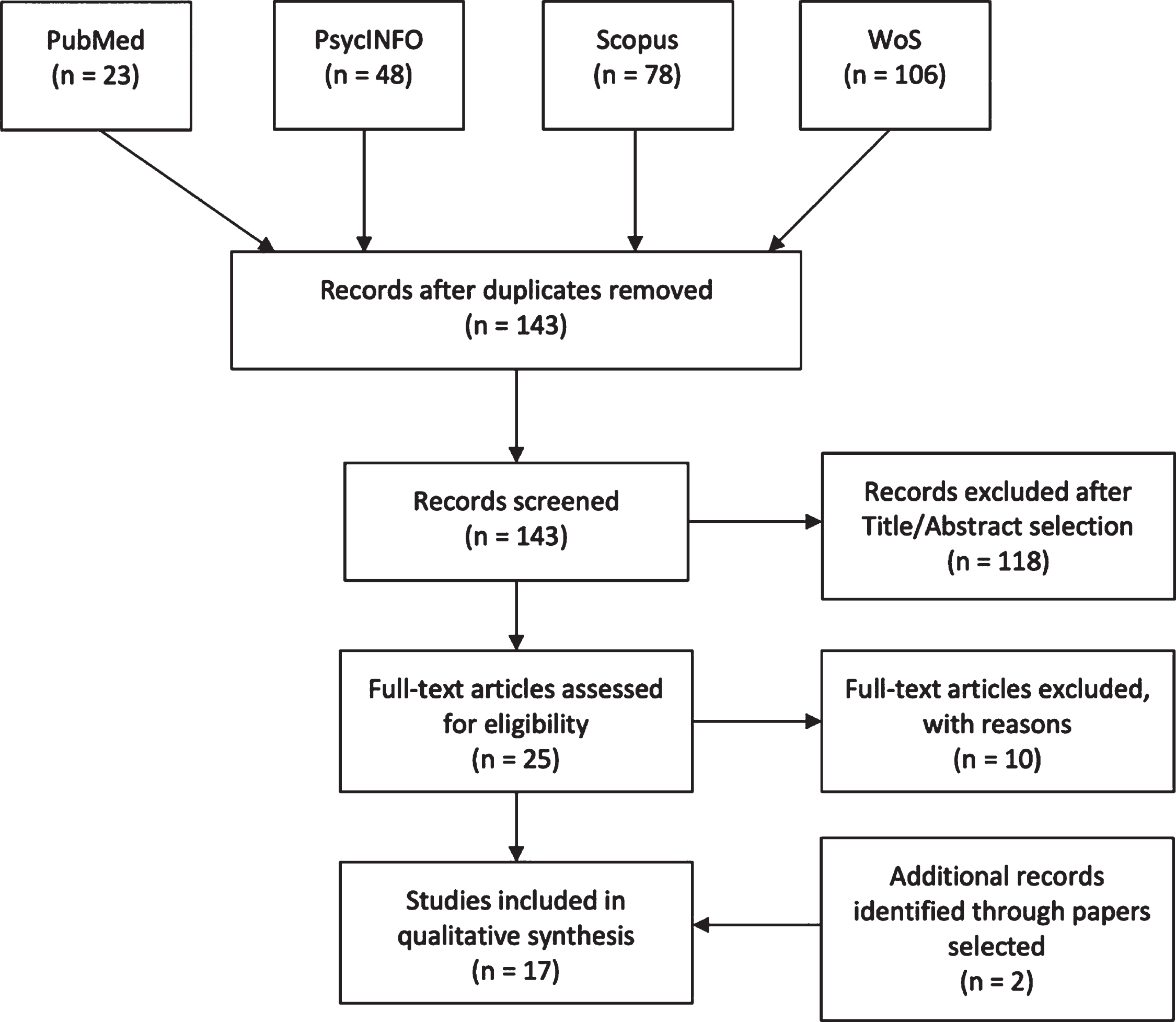
PRISMA flowchart
In this regard, VR is a simulative technology which offers ecological, controlled and standardized environments with different degrees of immersion (i.e., immersive, semi and non-immersive) and interaction (e.g., controllers, treadmill, joypad) [65, 82–84]. Indeed, VR can also be considered embodied thank to its potential to provide interactive and multisensory information [85]. VR mimics the neurocognitive process known as embodiment. During our daily experience, the body matrix, a supramodal multisensory body representation, allows us to correctly predict bodily experiences through top-down paths (predictive coding) and to feel located within our body and in the space around (sense of presence) [86]. In the same way, VR tries to predict simulations by generating the sense of presence in the virtual body and environment [87]. In this sense, virtual embodiment is the result of these two key elements and arises as the sense of controlling, owning and feeling located within a virtual body and in the space [88]. In particular, during navigation, effective virtual embodiment is provided by idiothetic and multisensory integration of information coming from internal and external cues [2, 89–91].
To our knowledge there is no systematic review summarizing the deficit of egocentric and allocentric spatial memory in real-world and virtual navigation tasks in MCI. We conducted this review to provide an overview of: 1) egocentric and allocentric spatial memory deficits in MCI profiles; 2) real-world versus virtual navigation performances in MCI.
METHODS
Preferred Reporting Items for Systematic reviews and Meta-Analysis (PRISMA) guidelines were followed [92].
Search strategy
Four high-quality databases (PubMed, PsycINFO, Web of Science, Elsevier-Scopus) were used to perform the search on 13 March 2020. Keywords used for each separate string were (“mild cognitive impairment”) AND (“egocentric” OR “allocentric” OR “spatial memory”) AND (“virtual” OR “real”) and were searched through Title/Abstract, Abstract, Topic and Title/Abstract/Keywords respectively for each database. Figure 1 shows the paper selection procedure and number of selected/excluded articles. Two papers [93, 94] did not appear in the databases and were added while reading the papers during full-text selection.
Selection criteria
Studies that uniquely investigated egocentric and allocentric spatial memory in MCI with real or virtual navigation tasks were selected. We excluded articles that did not specifically mention egocentric and allocentric memory by means of navigation tasks; that is where locations/paths where not encoded and retrieved through an act of active navigation [4, 74] for egocentric and allocentric memory. Additionally, studies where the full text was not available or where the abstract lacked basic information for review were removed. Non-English papers, reviews, meeting abstracts, conference proceedings, notes, case reports, letters to the editor, research protocols, patents, editorials, and other editorial materials were also excluded.
Quality assessment and data abstraction
PRISMA guidelines were strictly followed; search results found by the first author (CT) were shared with the review co-authors (VM, CS-B) for individual selection of papers in order to reduce bias, and disagreements concerning the type of navigation and spatial memory tested were resolved through consensus. PICO (Population, Intervention, Comparison and Outcome) guidelines were preferred as they provide more inclusive search results [95]. Hence, research question was “How are egocentric and allocentric memory (Outcome) affected in MCI (Population) with real-world and virtual (Intervention) navigation tasks?”. Comparison term was set as not applicable because, as emerged in an exploratory search, currently only few studies [75, 96] assess real-world versus virtual navigation in MCI for egocentric and allocentric spatial memory. The risk of bias for each single study was assessed following the Cochrane guidelines [97] by CT, VM, and CS-B (see Supplementary Figure 1); not admitted (N/A) answers were recorded as Cochrane risk of bias is suitable only for experimental designs. Despite we acknowledge that this systematic review does not concern a clinical intervention but the presence-absence of a deficit, Table 1 shows studies design according to definitions of experiment, quasi-experiment (QE) and non-experiment in systematic reviews [98]. Furthermore, all the papers included did not meet the criteria (e.g., randomization) of experimental studies and were considered as QE (see also risk of bias in Supplementary Figure 1); despite control over QE is less rigorous than experiments, external validity and strength of causal inference is still possible, especially when design-specific assumptions are met (e.g., independence assumption) [98]. In this regard researchers should bear in mind that results of included studies concerning the “effect estimates ... are valid at or near the locus on the ‘natural’ process that produced the exogenous variation in exposure” [98]. In order to assess methodological quality (reporting, internal and external validity and power), two authors (VM and CS-B) used the Down and Black checklist for randomized and non-randomized studies [99]. Quality scores are interpreted as excellent (26–28), good (20–25), fair (15–19), and poor (<14) [100] (see Supplementary Figure 2). As some items were strictly applicable for clinical intervention trial N/A items were also recorder: 34 for internal validity (bias), 52 for internal validity (confounding), and 24 for reporting. Average Down and Black scale score for the included studies was 14.17 (SD = 2.14) without N/A. Data extracted from each included study were as follows: reference, populations included, type of navigation task, study design, neuropsychological/behavioral correlates, neurological correlates and spatial memory findings in MCI.
Summary of the studies analyzed
A–, amyloid negative; A+, amyloid positive; AD, Alzheimer’s disease; aMCI, amnestic MCI; aMCImd, aMCI multiple domain; aMCIcom, aMCI with neuropsychiatric comorbidity; aMCIsd, aMCI single domain; APOE, apolipoprotein E; ApoE4+, positive APOE isoform E4; ApoE4 -, negative APOE isoform E4; BF1, basal forebrain - posterior part of the Nucleus basalis of Meynert; BF6, basal forebrain - the medial septal nuclei and vertical limb of the diagonal band of Broca; BVA, Blue Velvet Arena; edu, education; FCSRT, Free and Cued Selective Reminding Test; H, hippocampal; HC, healthy controls; HGT, Hidden Goal Task; Hp, hippocampus; IPP, inferior posterior parietal; MCI, mild cognitive impairment; MMSE, Mini-Mental State Examination; MWM, Morris Water Maze; naMCI, non-amnestic MCI; NH, non-hippocampal; Pc, precuneus; PCC, posterior cingulate cortex; PL, parietal lobe; QE, quasi-experiment; rCGM, regional cerebral glucose metabolism; RSC, retrosplenial cortex; S/S, homozygous for Short poly-T homopolymer; S/VL, heterozygous Short/Very long poly-T homopolymer; SCD, subjective cognitive decline; SMC, subjective memory complaints; SPL, superior parietal lobe; TMT, Trial Making Test; VRNT, Virtual Reality Navigation Task; VL/VL, homozygous for Very Long poly-T homopolymer; VR, virtual reality; vYSA, virtual Y-maze strategy assessment; ɛ3, allele 3; ɛ4 + /–, APOE genotype-ɛ4 heterozygous carriers; ɛ4–/–, APOE genotype-ɛ4 non-carriers; ɛ4 + /+, APOE genotype-ɛ4 homozygous carriers
RESULTS
Our search identified 17 studies that specifically assessed egocentric and allocentric memory with real-world and virtual navigation tasks in MCI. Findings are shown in Table 1 according to reference, year, sample(s), navigation task, neuropsychological correlates, neural correlates and main spatial memory findings. In particular, we will focus our result section on: 1) egocentric and allocentric memory deficits in MCI subtypes; 2) real-world versus virtual navigation performances of MCI.
Egocentric and allocentric spatial memory in MCI after navigation tasks
Egocentric and allocentric spatial memory in aMCI
Some studies investigated aMCI without distinguishing sd and md types by using Petersen’s criteria [44, 102]. In this cluster, egocentric memory recall was more impaired in aMCI than in healthy controls (HC) [72, 109]. Allocentric retrieval was similar among aMCI and HC in one study [109] but more impaired in other findings [54, 107]. aMCI performed similarly to HC in the mixed allocentric-egocentric recall in one study [109] but were more impaired in another [107]. aMCI performed better on allocentric recall than AD in Plancher and colleagues [106]. Interestingly, AD, aMCI, and HC were less impaired in allocentric but not egocentric recall in the active VR exploration than in the passive VR exploration [106]. Similar to HC, egocentric, allocentric, and mixed egocentric-allocentric recall performance was more impaired in aMCI than in subjective cognitive decline group [107].
Learning across egocentric task was similar among aMCI and HC across trials but impaired in the allo-centric learning [72]. Interestingly, Weniger and colleagues [72] found that larger volumes of the right-sided precuneus significantly predicted better performance on the egocentric memory task in both aMCI and HC. The authors also found that participants in both groups who had striatal vascular lesions committed more errors than participants without striatal lesions in egocentric memory task, while WMH did not affect egocentric or allocentric memory.
Interestingly, Keynejad and co-authors [104] showed that aMCI individuals with neuropsychiatric comorbidity (com-aMCI) were more impaired on egocentric memory recall compared to aMCI and HC. Both aMCI groups were equally impaired and more impaired than HC for the allocentric and delayed retrieval. In Weniger and colleagues [72], egocentric and allocentric recall performances between aMCI and HC were not accounted by spatial cognition test and depressive symptoms. However, egocentric learning (trials) by group interaction did not survive with these variables as covariates.
Conversely for neuropsychiatric symptoms, Key-nejad and co-authors [104] found that com-aMCI subgroups (anxiety disorder versus depression, dep-ression and anxiety or alcohol misuse) were equally impaired for all the subtasks of the HGT.
Subjective memory complaints and naMCI groups performed as well as the control group in all the subtests of the versions [96, 108]. Finally, aMCI versus naMCI spatial memory performances were not reported in one study [108].
Egocentric and allocentric spatial memory in aMCI with single and multiple domain
Another cluster of studies investigated the deficits according to sd and md typologies [44, 102]. In the real-world and VR version Hort and colleagues [96] found that AD and aMCImd were more impaired than the HC in all the recall subtests. aMCIsd performed worse than the HC in the allocentric and delayed recall of the real and virtual versions but as HC in the other subtests [94, 96]. The aMCIsd performed as the AD group in the delayed recall of the real and computer version but in all the other recall subtests they performed better [96]. Similarly, Mohammadi and co-authors [55] showed that egocentric and allocentric retrieval was more impaired in mild AD and aMCImd patients than in aMCIsd and HC [55]; moreover, egocentric recall was more impaired in aMCIsd, aMCImd, and mild AD than allocentric memory retrieval.
Learning abilities of the aMCImd was similar to AD group, expect for the virtual mixed allocentric-egocentric task [96]. aMCIsd had similar learning effect to HC for the allocentric VR (last trials but not first trial), egocentric real/VR, and allocentric-egocentric real task [96].
Egocentric and allocentric spatial memory in aMCI with hippocampal and non-hippocampal amnesia
Here we discuss the findings of navigation performances of individuals with H and NH amnesia detected by means of the FCSRT [41]. Consistently, egocentric, allocentric, and delayed allocentric recall were more impaired in HaMCI than NHaMCI [75, 108]. Mixed egocentric-allocentric memory retrieval was more impaired in HaMCI than NHaMCI in two studies [75, 94] but not in one research [108]. HaMCI was as impaired as mild probable AD for the egocentric, allocentric, and delayed recall but outperformed AD for the mixed recall [75], but in two studies [94, 108] HaMCI performed equally to AD in all the tasks. NHaMCI outperformed mild AD patients in all the spatial recall tasks in one work [75] but in other findings [94, 108] delayed test was as impaired as mild AD patients. Both HaMCI and NHaMCI were more impaired than controls for all the four HGT recall measures, with NHaMCI having less severe deficits [75, 108].
Learning across allocentric task was more im-paired in HaMCI than NHaMCI [75, 108], whereas contrast results were found for egocentric learning [75, 108]. Mixed learning performance was better for NHaMCI than HaMCI [108]. In particular, learning HaMCI ability was worse than control group and similar to mild AD individuals [108].
Egocentric and allocentric spatial memory in aMCI with AD biomarkers
Concerning biomarkers of AD in MCI, studies mainly used genetic markers [48, 49]. aMCI ɛ4 + had lower spatial recall performances in all the tests except for the delayed task compared to ɛ4–group [93, 94]. aMCI ɛ4 + group had lower spatial recall performance in all the tests compared to HC [93]. aMCI ɛ4–group showed similar retrieval performances to controls for the egocentric and mixed recall but were outperformed in the allocentric and delayed recall [93]. aMCI ɛ4 + showed similar deficits to AD individuals [93, 94], whereas aMCI ɛ4–outperformed AD group in all the subtests in one study [93] but not in allocentric and delayed in another [94].
More specifically, in ɛ4 profiles, egocentric, allocentric, and delayed recall were more impaired in aMCI ɛ4 + /+, +/–than in the aMCI ɛ4 –/–groups [71]. Further, egocentric retrieval was more impaired in the aMCI ɛ4 + /+ than in the aMCI ɛ4 + /–groups. In allocentric and delayed recall, aMCI ɛ4 + /+ and aMCI ɛ4 + /–were equally impaired [71].
According to TOMM40 gene profiling, Laczó and colleagues [53] found that allocentric and delayed recall were more impaired in the aMCI S/VL, VL/VL than in the aMCI S/S groups. Egocentric and mixed egocentric-allocentric memory retrieval were equally impaired in aMCI S/VL, VL/VL, S/S groups. Interestingly, there were not any differences in the ɛ4 and TOMM40 profiles in the learning performances in all the subtests [53, 93]. Only one used amyloid pathology [38], and in this study [78], both amyloid positive (A+) and negative (A–) aMCI groups showed poorer navigation compared to controls (few items found); in A + group, errors were associated with both egocentric and allocentric recall deficits, whereas in A–group, only with allocentric deficit. Interestingly, aMCI due to AD [42] group preferred egocentric strategy but, overall, strategy preference did not affect allocentric navigation [76]. Those individuals who preferred the egocentric strategy had less accurate performance in the allocentric navigation task than those who preferred the allocentric strategy [76].
Egocentric and allocentric spatial memory in MCI in real-world versus virtual navigation tasks
In this section we will report MCI performances in the two navigation types. Only few studies compared real-world versus virtual navigation task [75, 96]. In Hort and colleagues [96], recall subtests performances of aMCIsd and aMCImd (aMCImd more impaired than aMCIsd) were equal in both real-world and virtual versions. For learning performances in the first trial of the virtual mixed egocentric-allocentric was similar to control in aMCImd reflecting short-term visual memory rather than long-term spatial memory. The aMCIsd group was equally impaired in the virtual and real-world allocentric in the first and last trials of task reflecting hippocampal involvement in both versions. The pattern of allocentric memory deficit in aMCImd and aMCIsd was evident with the two navigation tasks. Similarly, in the study by Laczó and collaborators [94], aMCIsd and aMCImd performed in the same way in the two navigation tasks and with the same allocentric impaired pattern. They found also that ApoE4 non-carriers had better egocentric memory performances than ApoE4 carriers, which performed as non-carriers in all the other subtests. However, learning trials were not analyzed for the groups. The aMCImd group did better than AD group only in the mixed allocentric-egocentric VR version and in the egocentric and allocentric real-world recall [96], but in Laczó and collagues [94], aMCImd had similar performances to AD individuals in all the recall tasks. Finally, naMCI performed as HC in both versions in some studies [94, 96]. Interestingly, in Laczó and collaborators [75], correlations between real-world and virtual versions were reported and showed good correlations (mixed egocentric-allocentric: r = 0.76, p < 0.001; egocentric: r = 0.82, p < 0.001; allocentric: r = 0.83, p < 0.001; delayed: r = 0.65, p < 0.001). They found the same pattern in recall performances between HaMCI and NHaMCI (equal allocentric and delayed performances but HaMCI showed greater impairment for egocentric and mixed task) and between H/NH versus HC in the two versions. This pattern was also confirmed in Laczó and collaborators [94]. Learning performances were more impaired in the real-version for HaMCI in all subtests [108] but in the virtual was equal for egocentric memory in HaMCI and NHaMCI [75]. Parizkova and co-authors [76] used a virtual version of the Y-maze to detect MCI due to AD with allocentric or egocentric strategy and study the impact on real-world navigation. They found that aMCI due to AD individuals who preferred egocentric strategy (67%) compared to those who preferred an allocentric (33%) navigation in the virtual task had less accurate real-world allocentric memory. This pattern detected with VR reflects AD pathology that affects medial temporal lobes and leads individuals to adopt compensatory strategies to overcome allocentric deficits. The rest of the studies did not compare real versus virtual versions; hence we provided a qualitative overview of intra-MCI subtypes differences in Table 2.
Qualitative comparison of real-world versus virtual navigation studies in aMCI
-, non-carriers; +, carriers; <, significantly worse than; =, not significantly worse than; A, amyloid pathology; aMCI, amnestic mild cognitive impairment; aMCIcom, aMCI with neuropsychiatric comorbidity; aMCImd, aMCI multiple domain; aMCIsd, aMCI single domain; S/S, homozygous for Short poly-T homopolymer; S/VL, heterozygous Short/Very long poly-T homopolymer; VL/VL, homozygous for Very Long poly-T homopolymer; ɛ4 + /–, APOE genotype-ɛ4 heterozygous carriers; ɛ4, allele 4; ɛ4–/–, APOE genotype-ɛ4 non-carriers; ɛ4 + /+, APOE genotype-ɛ4 homozygous carriers
DISCUSSION
In the present paper we reviewed studies that assessed egocentric and allocentric spatial memory in MCI, by means of real-world and virtual navigation tasks. According to the sections, our work can be summarized into the following points: 1) aMCI was the most studied subtype as critical prognostic entity for AD; in particular, aMCI is characterized by spatial memory impairments and, regardless the aMCI subtype, deficits are not only confined to allocentric memory, but extend also to egocentric abilities; moreover, neuropsychological and genetic profiles show specific spatial memory patterns potentially useful to improve diagnosis; 2) the preferred navigation task is the 2D VR HGT or the HGT in the BVA; 3) real or virtual navigation tasks can be equally sensitive to detect navigation deficits in MCI. To our best knowledge this is the first systematic review that evaluates egocentric and allocentric memory impairments in MCI sample. Our findings could be used to plan future studies to improve diagnosis and prognosis of MCI. More, the review could be used as a starting point to design adequate and innovative rehabilitative solutions for MCI by means of VR thanks to its multisensory, interactive, ecological, standardized, adaptable, and safe potential for active navigation tasks in aging. Findings should be taken with caution as we found poor methodological quality due to the use of QE; however, as the included studies are not concerned with clinical interventions bur rather are interested in detecting a deficit in clinical populations, we must acknowledge that part of this bias is not due to lack of experimental rigor but rather in the design its self of the studies, which could be hardly carried out with a different method.
MCI is an heterogenous diagnostic entity in its neuropsychological and neuropathological profiles and has been initially observed in patients with memory impairments at higher risk of developing AD [51]. Indeed, in our review the most studied subtype was aMCI. This group was categorized by the use of neuropsychological marker (i.e., FCSRT) [41] or with AD biomarker, such as the presence of positive amyloid pathology or individuals at genetic risk of AD [38, 49]. Despite the crucial role of aMCI in AD and the consequent scientific and social impact, current spatial memory research did not focus on other MCI types and etiologies. Recent researches showed that egocentric and allocentric memory can be impaired in other neurodegenerative diseases, such as vascular [61, 62], Lewy bodies [58], frontotemporal [59], and even Parkinson’s disease [60, 111]. Additionally, subjective navigation complaints have been reported in individuals with naMCI and subjective cognitive decline [63]. Hence, future research should strengthen/expand aMCI findings but also focus on MCI with other underlining diseases (e.g., [112–115]). Defining the underlining etiology is crucial for differential diagnosis among MCI subtypes, for instance striatal lesions detected with Fazekas’s criteria impacted egocentric memory of both HC and aMCI. This is consistent with initial findings of spatial memory deficits in vascular cognitive impairment [61, 62]; moreover, vascular risk factors for AD were also found to affect egocentric memory as well [107].
Learning and recalling spatial information is affected in MCI and the severity of these deficits can be located between the performance of HC/subjective cognitive decline and AD individuals. This is consistent with the notion of MCI as a transitional stage between aging and dementia [51], where also spatial memory and navigation should be considered as affected domains of this subclinical entity [116]. Impairment in allocentric memory is in line with previous neuropathological and neuropsychological alterations in MCI and AD [40, 116] Consistently, the studies included that investigated neural underpinnings showed hippocampal (particularly on the right side), and more general medial temporal, involvement. This is consistent with the consolidated role of these regions in spatial memory [117–119]. Interestingly, we found also egocentric deficits that could be related in aMCI to reduced activation of the caudate and the precuneus during spatial navigation task [120]. Indeed, these regions are crucial for egocentric navigation [22, 121] and spatial visual imagery (the so called “parietal window”) [13]. The egocentric deficit could be exacerbated by neuropsychiatric comorbidities in aMCI due to fronto-striatal alterations [104, 122]. Only one study [55] compared egocentric versus allocentric memory and, surprisingly, found that egocentric memory was worst in AD, aMCI, and HC compared to allocentric memory. This is in contrast with egocentric navigation preference in aging and MCI [33, 76]. However, to successfully navigate the environment a flexible combination and switching of the two navigation modes is required [29, 116]. Importantly, only one study assess allocentric dependent-independent view-point abilities after navigation [54], a critical point as switching abilities between spatial frames is crucial for navigation and is impaired in AD individuals [24, 123]. However, aMCI preferred learning was affected only for allocentric memory, indeed aMCI due to AD pathology preferred egocentric rather than allocentric strategy.
More specifically, we found that aMCImd has greater impairment of both types of spatial memory (learning, immediate, and delayed recall), similar to AD, compared to aMCIsd but the latter should be considered as a hippocampal subtype. Indeed, spatial navigation requires a large network of brain regions and cognitive functions to successfully navigate and recall the environment (for a review, see [28]), hence aMCImd could have poorer performance as multiple neuropsychological functions are impaired. Conversely, by a neuroanatomical point of view, aMCIsd could be conceptualized as a ‘pure’ allocentric phenotype [96]. This resembles the pattern of H and NH aMCI, distinguished by means of the FCSRT [41]. We found that HaMCI had similar performances to AD on spatial learning and retrieval, whereas NHaMCI had less pronounced deficits in navigation. HaMCI and aMCIsd could be thought as prodromal ‘hippocampal’ stages of AD [41, 116] and thus it would be possible to improve diagnosis, prognosis and treatment. Indeed, AD pathology affects not only memory per se but the neural substrate of idiothetic navigation itself and spatial behavior as noted by differences in A + and A–individuals [78]. This finding is also interesting within the context of embodiment in spatial memory, where multisensory cues play a critical role for representation of the body and the space [2, 90]. Indeed, embodiment might lead to alterations in egocentric and allocentric abilities in normal and pathological (i.e., AD and Parkinson’s disease) aging [31, 124] and spatial memory can be improved by adopting active navigation as shown by a recent systematic review of Tuena and colleagues [74].
Given the crucial role in pathological aging, spatial memory could be used as a marker of AD pathology to improve differential diagnosis. Individuals with genetic markers of AD, such as ɛ4 and ɛ3 (VL) carriers, have particular deficits in recalling spatial memory rather than in spatial learning [53, 93]. Allocentric and delayed tests impairment are associated with VL compared to S poly-T homopolymer, whereas ɛ4 carriers can be distinguished from non-carriers in egocentric and allocentric memory. More, egocentric memory can tap subtle differences among heterozygous, homozygous and non-carriers. Preserved egocentric abilities can also be a marker of individual with aMCI but without A pathology. However, both types of spatial memory can successfully discriminate aMCI individuals with AD pathology from those who have only neuropsychological marker of aMCI [78].
Finally, delayed recall of allocentric memory is crucial for distinguishing aMCI of sd, md H and NH types from HC and AD, between H and NH amnesias and S-VL/VL and S/S carriers but not between ɛ4 carriers and non-carriers. Conversely, mixed performances show also this patter for ɛ4 carriers and non-carriers but not between aMCIsd and HC. It is interesting to note that in this subtask the individual is provided with both types of spatial frames and consequently could use the preferred navigation strategy. Consequently, by forcing the adoption of one of the spatial frames the results become consistent between the groups and tap specific impairments like the ‘pure’ allocentric deficit in aMCIsd.
Unfortunately, as some studies did not provide full egocentric versus allocentric performances, MCI subtypes comparisons or did not stratify the MCI population, future research should provide at least neuropsychological profiles and their statistical differences or trends of their spatial memory.
The most used navigation task real-world or virtual version of the human analogue (HGT) of the Morris water maze [70]. However, other studies used other spatial navigation tasks such as the frame matching task by Serino and colleagues [54], the large-scale real-world environment in Schöberl and co-authors [78] or the active versus passive navigation semi-immersive scenario in Plancher and collaborators [106]. Involvement of different research groups and paradigms would reduce bias and improve basic and clinical research in this topic. On the one hand, it is crucial for replicability to use consolidated tasks, on the other hand future research would benefit from more heterogenous navigational tasks. For instance, by means of discrete landmark/egocentric and boundary landmark/allocentric navigation strategies [22, 119], it is possible to tap procedural (striatal) and declarative (hippocampal) spatial memory; this paradigm could be applied to study diseases where one or both the neuroanatomical underpinnings are affected by certain diseases (e.g., Parkinson, Lewy body, WMH).
Surprisingly, our results show that real and virtual versions can provide similar results across aMCI subtypes. Vision is the most salient sensory input provided for navigation, and visual cues are indeed the most critical reference when self-motion cannot provide correct spatial orientation [11, 125]. However, some subtasks might involve other memory systems than immediate and delayed memory, such as short-term visual memory. This is consistent with findings showing that path integration and spatial memory involve medial temporal regions only when adequate number of items and delay are used for encoding and retrieval [80, 126]. Nevertheless, we think that current VR solutions still lack high degrees of virtual embodiment by means of semi-immersive/immersive apparatus. By involving embodiment and multisensory inputs [86, 87] it could be possible to improve idiothetic features (e.g., proprioception, interoception, vestibular information) for simulating real-life navigation [89, 127].
Portable neuroimaging/neuromodulation devices could be used to study real time navigation and spatial memory [128, 129]; indeed, studies reported here use ‘off-line’ paradigms. However, our results are in line with previous research concerning the neuropsychology of spatial memory [10, 118] and AD models [39, 123]. Interestingly, Schöberl and colleagues [78] showed that ‘active navigation’ has different metabolic pattern than locomotion and that navigation regions are impaired in MCI. Using regional cerebral glucose metabolism (rCGM), they compared navigation with locomotion (locomotion only without navigation task) and found that involved different brain regions (right anterior hippocampus, bilateral RSC, and pontine tegmentum) in healthy participants. In aMCI patients a relative increase of rCGM in the pontine tegmentum and in the right anterior hippocampus in the control group was found. A + aMCI showed lower rCGM during navigation compared to locomotion in the right anterior hippocampus, bilateral RSC cortex, precuneus, and parietal lobe compared to controls. A + compared to A–aMCI group revealed reduced rCGM in the bilateral hippocampus and right superior parietal lobule and increased metabolism in the pontomedullary tegmentum and left frontal lobe. Again, Schöberl and colleagues [78] found that both A + and A–groups had lower gait speed during navigation compared to controls. The findings are consistent with the critical role of active navigation components used to derive spatial information [4] and that the use of both bodily and cognitive factors during exploration could be included in memory assessment and rehabilitation for pathological embodiment in aging [1, 74].
Again, Schöberl and colleagues [78] explored visual exploration in aMCI individuals finding that A–patients had more lateral and total fixations and total, egocentric, allocentric, and search saccades than A + individuals. A–group fixed fewer unique landmarks during allocentric task than controls and A + patients, and A + and A–spent more time at crossings and neglected shortcuts compared to control group. Concerning memory tests, the authors showed that overall, egocentric and allocentric navigation performances differentiate A + from A–, rather than auditory-verbal learning, executive functions, and global cognition tests. In particular, the FCSRT has an important role when combined with spatial memory measures for diagnosis of MCI [41, 108]. Further studies should deepen the diagnostic, and consequently cognitive rehabilitation, peculiarities of MCI profiles, as spatial memory combined with neuropsychological tests and behavioral information might help to better differentiate this complex entity.
Behavioral measures are very useful for studying MCI and dementia. Indeed, if on the one hand neuropsychological tests of memory and visuospatial functions were associated with allocentric and egocentric memory respectively and spatial rotation with frames switching abilities [54, 110], on the other hand, egocentric and allocentric memory should be investigated and treated as separate entities (“Spatial Navigation”), which has important consequences for diagnostic and rehabilitative purposes. Indeed, Laczò and colleagues [110] conducted an exploratory factor analysis that revealed that allocentric and egocentric navigation scores loaded on their own factor, separately from the other cognitive functions (verbal and non-verbal memory, executive and visuospatial function, attention/working memory and language function).
Limitations of current spatial memory research in MCI could be summarized in the need to: 1) consider different MCI etiologies and/or neuropsychological deficits in assessing spatial memory and expand researches on aMCI/MCI due to AD with larger sample size; 2) employ other tasks besides HGT, including also the assessment of switching abilities; 3) improve the VR tasks, including idiothetic cues, full-immersive solutions with comparison with real versions; 4) assess all the differences and trends within the MCI typologies (naMCI included), frame switching abilities and the potential deficits in embodiment (virtual embodiment when using VR) in this sample and how they affect spatial memory and active navigation; and 5) improve blinding of outcome measures, state any missing data and explain how it was handled, add power analysis, improve the control over confounding variables and clarify in detail MCI subtypes/spatial memory differences in hypotheses and results.
Conclusions
Although egocentric and allocentric spatial memory decline has been deeply investigated in both healthy older people [32] and in AD individuals [40], to our knowledge a systematic review focusing on MCI patients is lacking. aMCI is the most studied subtype give its importance for AD and spatial memory with deficits in both egocentric and allocentric memory; aMCI with AD markers shows specific spatial memory impairments instead. However, future research could investigate how navigation could be impaired in other MCI subtypes. This can be done by using VR and real-world tasks. In particular, VR solutions showed good usability in older people and it could be useful to investigate how bodily information (embodiment) relates to spatial cognition, also combined with artificial intelligence methods, to improve diagnosis and treatment [130–142]. In conclusion, identifying cognitive markers typical of this transitional condition might allow also early neurorehabilitation interventions to prevent the progression to dementia.