
Abstract
The amyloid-β oligomer (AβO) hypothesis was introduced in 1998. It proposed that the brain damage leading to Alzheimer’s disease (AD) was instigated by soluble, ligand-like AβOs. This hypothesis was based on the discovery that fibril-free synthetic preparations of AβOs were potent CNS neurotoxins that rapidly inhibited long-term potentiation and, with time, caused selective nerve cell death (Lambert et al., 1998). The mechanism was attributed to disrupted signaling involving the tyrosine-protein kinase Fyn, mediated by an unknown toxin receptor. Over 4,000 articles concerning AβOs have been published since then, including more than 400 reviews. AβOs have been shown to accumulate in an AD-dependent manner in human and animal model brain tissue and, experimentally, to impair learning and memory and instigate major facets of AD neuropathology, including tau pathology, synapse deterioration and loss, inflammation, and oxidative damage. As reviewed by Hayden and Teplow in 2013, the AβO hypothesis “has all but supplanted the amyloid cascade.” Despite the emerging understanding of the role played by AβOs in AD pathogenesis, AβOs have not yet received the clinical attention given to amyloid plaques, which have been at the core of major attempts at therapeutics and diagnostics but are no longer regarded as the most pathogenic form of Aβ. However, if the momentum of AβO research continues, particularly efforts to elucidate key aspects of structure, a clear path to a successful disease modifying therapy can be envisioned. Ensuring that lessons learned from recent, late-stage clinical failures are applied appropriately throughout therapeutic development will further enable the likelihood of a successful therapy in the near-term.
Keywords
INTRODUCTION TO THE AβO HYPOTHESIS
The transition from the amyloid cascade to the AβO hypothesis
The detection of amyloid-β oligomers (AβOs) in human brain parenchyma and vasculature was first reported while the original amyloid cascade hypothesis was being introduced and developed [1–3]. At the time, AβOs were regarded as intermediates en route to generation of amyloid plaques, which were believed to be the pathogenic form of Aβ.
Today, AβOs are widely regarded as the most toxic and pathogenic form of Aβ (Fig.1) [4, 5]. AβOs show an Alzheimer’s disease (AD)-dependent presence in humans and animal models [1, 6–13], and their buildup occurs early, before plaques, evidenced by both immunochemistry [14] and immunohistochemistry [15, 16]. In support of a toxic role for AβOs and not plaques, the Osaka familial AD mutation of Aβ (APP E693Δ) shows extremely low levels of senile plaques [17–21] despite severe cognitive impairment [17, 20], while cerebrospinal fluid (CSF) manifests low levels of overall Aβ, but elevated levels of AβOs [22]. Transgenic (Tg) mice carrying this mutation [19], or a closely related one [23], likewise manifest AβOs and other major forms of AD neuropathology but not plaques. Although historically AD has been defined as dementia with plaques and tangles, replacing plaques with AβOs in this definition may be closer to the pathogenic mechanism.
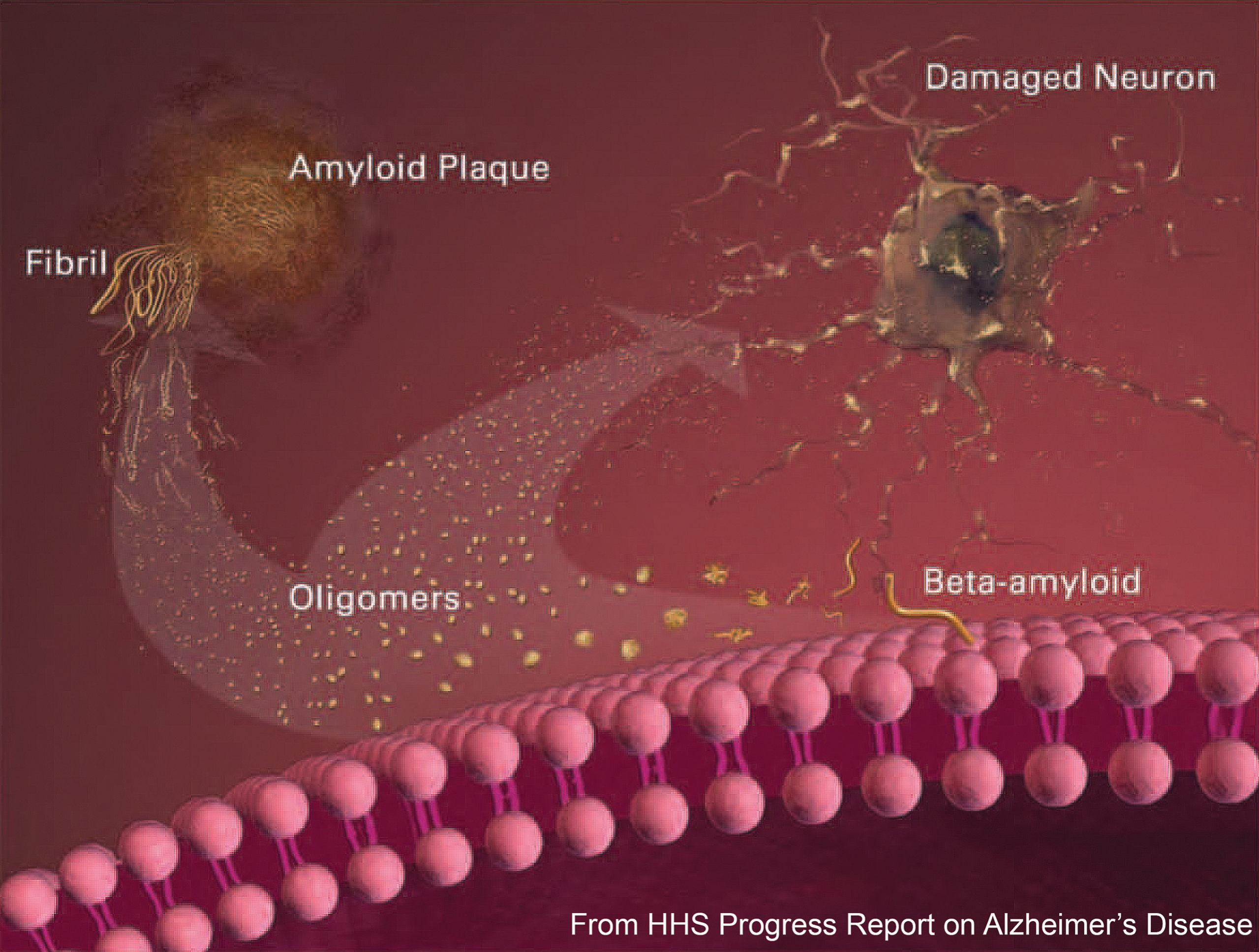
Synthetic AβOs can assemble at very low concentrations of Aβ42 monomer, in harmony with pre-plaque buildup in brain tissue [24, 25]. In vitro, AβOs form within minutes from low nanomolar concentrations of monomeric Aβ42 [26, 27]. Because Aβ has been found to aggregate in sodium dodecyl sulfate (SDS) [28], some investigators have concluded that the quickly forming AβOs seen in SDS-PAGE are experimental artifacts [29, 30]. However, under full denaturing conditions, SDS-PAGE experiments show monomeric Aβ in the complete absence of AβOs [31]. AβOs can be observed, moreover, in the absence of SDS by atomic force microscopy (AFM) and by size exclusion chromatography [26, 31]. Evidence for structural homology between certain forms of synthetic and brain-derived AβOs has been presented [6]; this is discussed further below.
Besides their presence in brain, AβOs show an AD-dependent buildup in human CSF. An ultrasensitive assay, known as the BioBarcode, which is capable of attomolar measurements, showed median levels of AβOs in CSF from AD patients to be 30-fold higher than from non-demented individuals [32]. This elevation is opposite to the AD-dependent change measured in monomeric Aβ levels, which decrease rather than increase [33]. Levels are so low, however, that for most assays, comparisons of CSF AβO levels are not feasible [12, 35]. Ultrasensitive assays for AβOs in CSF, however, are extremely lengthy and difficult, and their lack of precision requires multiple runs to provide a reliable measurement. These are all factors that preclude their adaptation for the clinic.
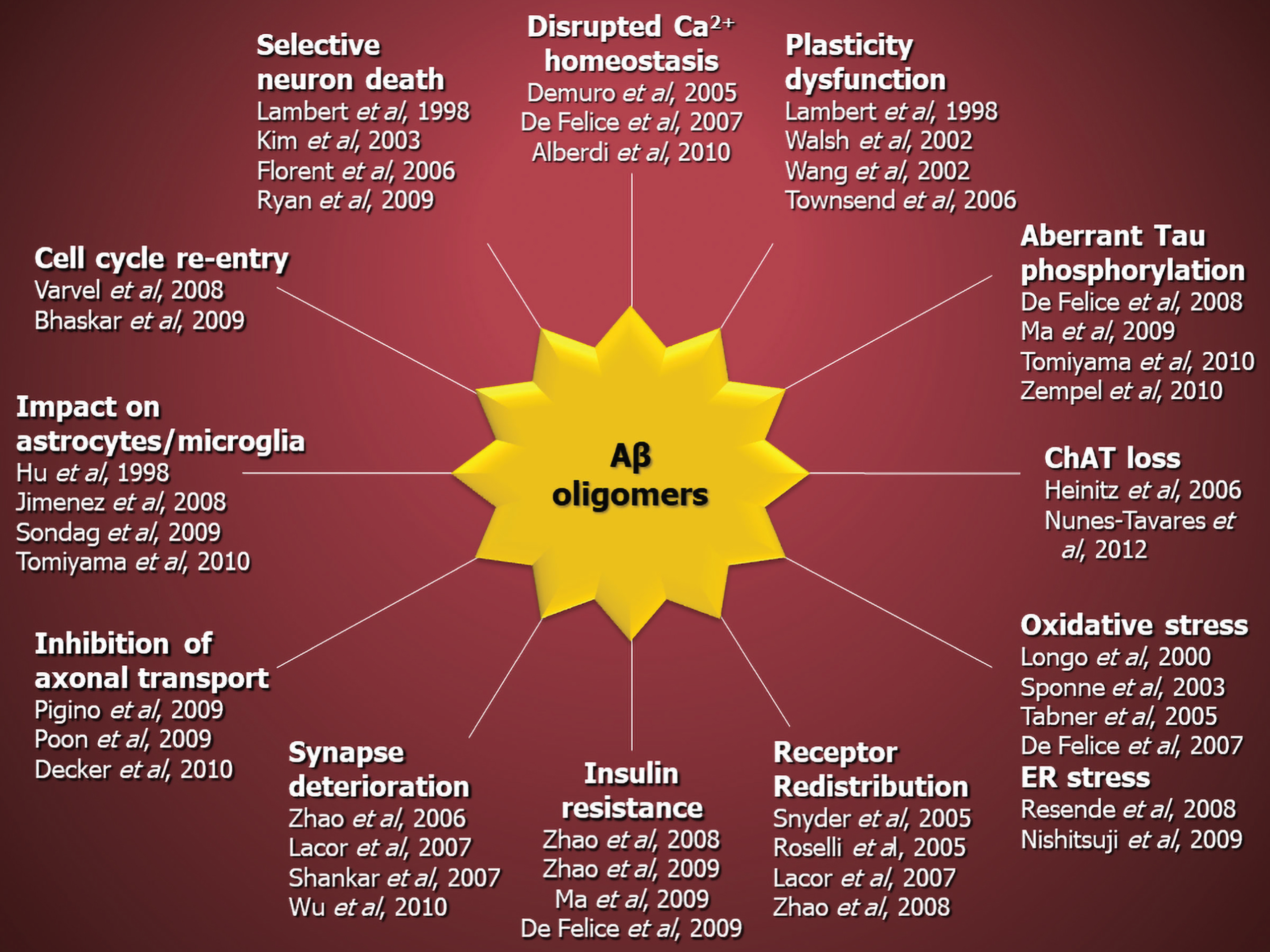
There is extensive evidence that elevated AβO levels in the brain has pathogenic consequences, with results coming from behavioral, neuropathological, and cell biological studies, as discussed below. Memory performance is lost when small quantities of AβOs are injected into the intracerebral ventricle (i.c.v.) of non-Tg animals [36–39]. Long-term potentiation (LTP) and long-term depression, the electrophysiological underpinnings of memory formation, are disrupted by AβOs both ex vivo and in vivo [26, 41]. Synthetic and brain-derived AβOs both exhibit these characteristics. In addition to their cognitive impact, exogenous AβOs instigate multiple facets of AD-neuropathology in culture and animal models, including non-human primates (NHPs) [42–46]. If one assumes an AβO molecular weight in aqueous solution of ∼100 kDa (see below), these effects are elicited at sub-nanomolar AβO concentrations [26, 47–50]. Overall, AβOs have been found to instigate tau pathology [19, 52], loss of neuronal polarity [53–55], impairment of axonal transport [56–58], deterioration of synapses [47, 55], oxidative stress [59–62], endoplasmic reticulum (ER) stress [18, 64], insulin resistance [48, 65–67], neuroinflammation [19, 69], cholinergic impairment [70, 71], loss of trophic factors [45, 72–75], epigenetic changes [74, 76–80], ectopic mitosis [81–83], and selective nerve cell death [26, 84]. A complicating factor is that these various responses were obtained under widely divergent conditions, with different disease models, time-scales, doses, and AβO preparations. Nonetheless, the collective body of evidence offers strong support for a mechanism in which AD neuropathology and cognitive loss are the consequences of the cellular damage instigated by AβOs (Fig.2).
The evidence is strong that AβOs are manifested in AD brain. Experiments in animal models strongly suggest, furthermore, that AβOs are both necessary and sufficient for dementia. Sufficiency, at least vis-á-vis amyloid plaques, is indicated by instances of AD without senile plaque pathology. Highly demented individuals with the Osaka mutation (APP E693Δ) manifest AβOs (and other facets of AD pathology) in the absence of senile plaques [17–21]. This has been experimentally recapitulated in a Tg mouse model harboring this mutation [19]. In addition, Tg mice expressing a different mutation in the same APP residue (Dutch APP E693Q) also exhibit AβO accumulation and altered synaptic structure without plaques [23, 85]. The sufficiency of AβOs for pathogenesis was first indicated in an APP mouse (Indiana APP mutation V717F; outside of Aβ42 sequence) that showed synapse loss despite absence of plaques [86]. In addition, a Tg rat expressing the Indiana mutation also shows pre-plaque AβO-associated cognitive impairment [87]. A later study comparing Tg strains indicated in fact that elevated levels of amyloid plaques likely protected against pathogenic AβO buildup [88].
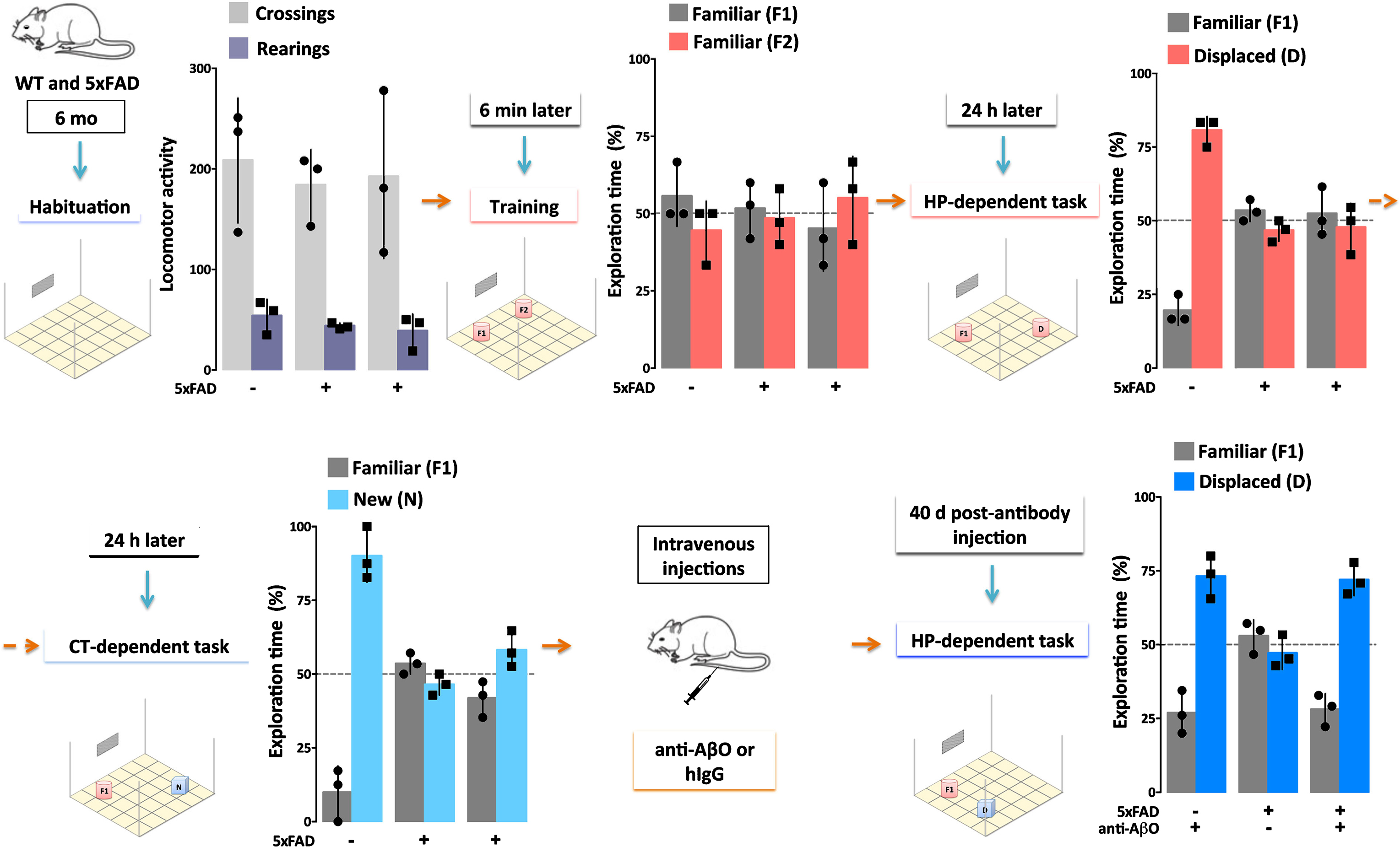
Direct evidence that AβOs are necessary for dementia comes from experiments using AβO-selective antibodies. Such antibodies were first shown to protect cell models against the damage caused by exogenous AβOs [51, 90]. When administered to various Tg AD mice, the antibodies prevent AD-like pathology and rescue memory performance [89–94]. New data from our group indicates that a single injection of an AβO-selective antibody (30μg) can suffice to rescue memory performance in 6-7-month-old Tg 5xFAD mice for at least 40 days (Fig.3; Bicca and Klein, unpublished data). AβOs and plaques in these mice begin to accumulate extensively around 2 months of age [11, 95]. The new data are in harmony with previous evidence that an AβO-selective antibody can reach the parenchyma and engage AβOs [96], but not Thioflavin S (ThioS)-positive amyloid plaques, when injected into 5xFAD mice.
The large body of evidence that AβOs are both necessary and sufficient to trigger AD-associated memory malfunction and neurodegeneration, coupled with the extensive portfolio of documented AβO-triggered cellular and behavioral effects, sets the stage for new AD therapeutic approaches targeting AβOs. As the third decade of the AβO hypothesis begins, the biggest challenge is to mobilize a clinical trial that will validate or invalidate the hypothesis. While “Aβ dyshomeostasis has emerged as the most extensively validated and compelling therapeutic target” [5], the past development of Aβ-based therapeutics has largely concerned plaque elimination, ignoring AβOs. However, the link between plaques and cognitive dysfunction has been tenuous for decades [97–99], and no Aβ-directed therapeutic has yet reached a clinical efficacy endpoint [100–103]. In a potential turning point, an antibody that can engage AβOs, Aducanumab, has recently shown modest therapeutic benefit in early clinical trials [104, 105]. A potential limitation of Aducanumab is that it lacks stringent selectivity for AβOs. Off-target engagement with senile plaques likely accounts for the high dosage-requirement found in trials. Antibodies are needed that target only the most pathogenic configurations of Aβ, i.e., AβOs. Such antibodies will be optimized by a better understanding of AβO structure-toxicity relationships [101, 106–108],
Besides development of AβO-specific antibodies [89, 109], other tactics are likely to improve the prospects of Aβ-directed therapies. Such tactics may be earlier intervention within the disease continuum and better criteria for patient selection [5, 108] and better biomarkers for monitoring of investigational new drugs [103], including inflammation markers to better predict complications [106]. Furthermore, multi-factorial therapies may be needed [106]. Although it has been suggested that Aβ-targeting therapies may only be beneficial in prodromal individuals [110], if AβOs play a role in disease progression, e.g., through promoting propagation of tau pathology (below), there may be a meaningful chance that AβO-immunotherapy would be beneficial even after AD onset. Overall, there is an important call for more rigor in preclinical development. At each phase of the drug discovery process for Aβ-targeting therapies, it has been possible to find significant gaps in data [102]. Target engagement, e.g., was not established for the majority of therapeutic agents analyzed [102, 107]. Furthermore, compounds have been moved into phase III trials on the basis of very limited data [111], premature moves that have had a tendency to poison the well. Consequently, the discouraging track record of Aβ-directed drugs has provided significant impetus to point new drug discovery efforts toward non-Aβ targets, despite the preponderance of evidence that AβOs are the culpable AD neurotoxins [112–114].
STATUS OF THE FIELD
In lieu of clinical efforts based on the AβO hypothesis, there nonetheless have been substantial developments in the last five years regarding more fundamental issues. Of the more than 4,000 publications on Aβ oligomers or oligomeric Aβ, about half were published in the last five years. These fundamental developments regarding AβO pathogenicity are just beginning to be tested clinically and we predict that they will set the stage for therapeutic success. This section will consider major developments regarding: 1) species of AβOs, their assembly, and relation to amyloid plaques, and emerging insights into how to approach molecular structure; 2) mechanisms of how AβOs initiate their impact on neuronal function and structure; 3) downstream pathways resulting in neural damage, and 4) multicellular interactions contributing to AβO pathogenicity.
A multitude of AβO species or just two?
One of the biggest knowledge gaps currently facing the field is the precise identity of the most toxic AβO structures [5, 115–117]. Without this knowledge, it is impossible to know if Aβ-directed therapeutics are engaging the correct target. Characterization of AβO structure has been hindered by AβO metastability and heterogeneity [116, 117]. Consequently, a multitude of AβO species have been identified in the literature [117]. It is not clear which of these AβO species are AD-relevant and which are experimental artifacts. One possibility is that there exist a multitude of pathogenically-relevant AβO species in the AD brain and that their high number correlates with the myriad Aβ-associated toxic pathways identified in the literature [29, 115]. Another possibility is that there are only a few discrete AD-relevant species, and the majority identified in the literature are merely artifacts induced by non-physiological experimental conditions [107, 117]. As stated by Benilova and colleagues, “The lack of a common, agreed-upon experimental description of the toxic Aβ oligomer makes interpretation and direct comparison of data between different research groups impossible [117].”
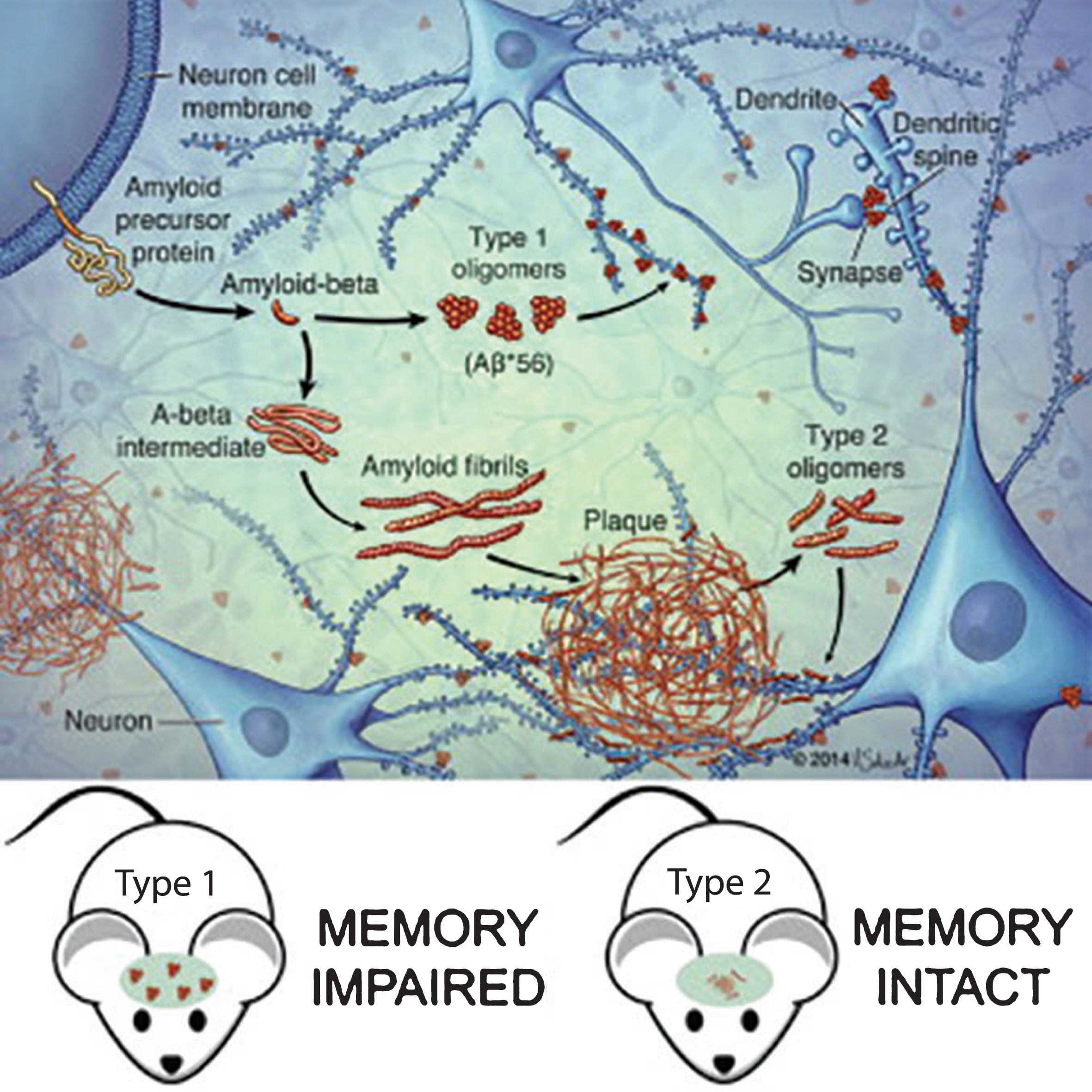
Some patterns regarding AβO structure-toxicity relationships are, however, already emerging in the literature. For instance, it appears as if AβOs, whether produced in vitro or present in the brain of animal models or AD patients, can be divided into toxic and non-toxic sub-populations based on simple aspects of their quaternary structure, molecular weight and antibody reactivity, as well as their relationship to amyloid plaques. The toxic AβO species appear to be greater than 50 kDa [16, 118], reactive with the anti-amyloid oligomer antibody A11 [119] and the anti-AβO antibody NU4 [120], and unrelated to amyloid plaques (Fig.4) [118, 119]. On the other hand, the non-toxic AβO species appear to be less than 50 kDa [16, 118], reactive with the anti-fibril antibody OC [119], and related to amyloid plaques temporally, spatially, and structurally [118, 119]. In addition to their convenient immuno-identification, they also can be separated in vitro by size exclusion chromatography [31] or ultrafiltration with a 50 kDa molecular weight cutoff [16, 118]. These populations have been referred to in the literature, respectively, as “peak 1” and “peak 2” [31], high molecular weight (HMW) and low molecular weight (LMW) [16, 118], and “type 1” and “type 2” [119]. Myriad evidence supports a toxic role for type 1 AβOs. In vitro, they bind cultured synapses (Fig.5) [16, 118], inducing production of reactive oxygen species (ROS) [39], while type 2 AβOs cannot. Both species have been implicated in binding PrPc [121–123]. In vivo, type 1 AβOs disrupt memory function [39, 120]. Type 2 AβOs have been found not to be associated with memory dysfunction [119, 120], although in one study, they were [39]. HMW, type 1 AβOs appear to be most prominent AβOs in the AD brain under native conditions [124–126]. The differential toxic impacts of LMW and HMW AβO species has been recently reviewed by Ferreira and colleagues [115].
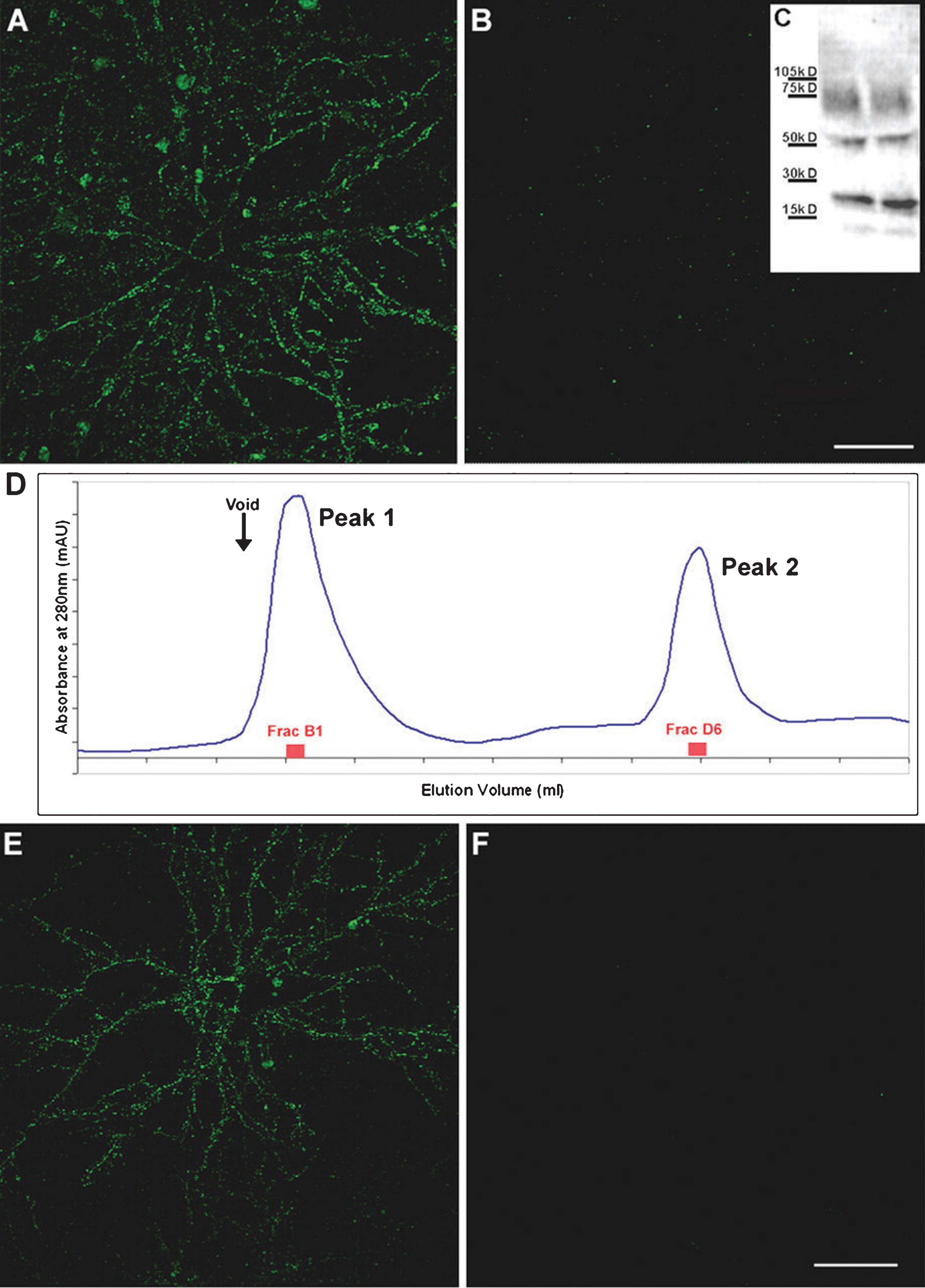
One specific type 1 AβO has been identified, the 56 kDa SDS-stable species sometimes referred to as Aβ*56 [44]. Aβ*56 was first identified as a prominent AβO in AD brain [6] and Tg2576 mice [44] and has been observed more recently in CSF [14]. A recent study by Lesné and colleagues compared the in vitro toxicity of Aβ*56 to two LMW species, dimers and trimers [127]. They found that Aβ*56 interacted with N-methyl-D-aspartate receptors (NMDARs), increased NMDAR-dependent Ca++ influx, and increased the activation of Ca++/calmodulin-dependent kinase IIα (CAMKIIα). The latter was associated with increased site-specific phosphorylation and missorting of tau. Dimers and trimers did not induce any of these effects. On the other hand, trimers were able to induce pathological conformational changes in tau, which was associated with a selective decrease in proteins governing axonal transport [128]. The lack of dimer toxicity is consistent with earlier observations from O’Malley and colleagues utilizing crosslinked dimers [129]. According to their results, they proposed that the contribution of dimers to AD is through their ability to further assemble into larger, more stable synaptotoxic assemblies. It is possible that the toxic response observed with trimers above was similarly due to their ability to assemble into large, more stable synaptotoxic assemblies [106] (i.e., HMW type 1 AβOs). This possibility cannot be discounted since the trimers were not conformationally stabilized in this study.
Many studies of AβO structure and function have been conducted with synthetic AβOs or AβOs derived from Tg mouse brain. Some researchers, however, are calling for analysis to be restricted to AD brain-derived AβOs [107, 130]. Yet, it seems as if there is structural homology between synthetic and brain-derived AβOs. For example, under non-denaturing conditions, synthetic and brain-derived AβOs show structural equivalence in terms of mass, isoelectric point, and immunoreactivity with conformation-sensitive antibodies [6]. Furthermore, as suggested above, at least three identical AβO species can be found in AD brain and the brain of multiple Tg mouse models: Aβ*56, dimers, and trimers [14, 128].
However, one important justification for restricting analysis to AD-derived AβOs is the increasingly apparent presence of Aβ proteoforms in the AD brain as well as the contribution of Aβ proteoforms to Aβ aggregation and toxicity (reviewed in [107, 131]). These proteoforms are not present in synthetic or cell-derived AβOs. It is well known that Aβ40 and Aβ42 are the most abundant Aβ peptides found in AD. However, in addition to these peptides, myriad truncated Aβ peptides also have been found in the AD brain [132] and CSF [133]. Using mass spectrometry, one study identified 26 unique Aβ proteoforms in the AD brain. 73% of these were N-terminal truncations and 30% were C-terminal truncations. The N-terminally truncated peptides were predominately found in the insoluble fraction of the brain, while the C-terminally truncated were predominately found in the soluble fraction. Canonical Aβ42 was a minority of the proteoforms identified and was equally distributed between the insoluble and soluble fractions [132].
Truncated Aβ peptides likely play a role in AD pathogenesis as they can form stable oligomeric complexes with the full-length Aβ42 peptide [133]. In fact, N-terminally truncated Aβ peptides formed through pyroglutamylation of glutamic acid residues are increasingly recognized as very toxic proteoforms of Aβ. Pyroglutamylated (pE) Aβ has been found to drive misfolding of Aβ into more toxic aggregates when present at 5–33% of the total Aβ concentration [134]. Pyroglutamylation also increases the aggregation speed of Aβ [135]. Anti-pE Aβ antibodies have been developed and successfully utilized in Fab form for co-crystallization with pE Aβ [136]. These studies revealed that the pE modification confers a pronounced bulky hydrophobic nature to the N-terminus of Aβ that might explain its enhanced aggregation properties. Interestingly, one group finds that pE AβOs may be the most abundant AβO species in AD brain [137]. Other Aβ proteoforms reported in the past 5 years to increase Aβ toxicity include C-terminally extended Aβ43 [138–140], Aβ peptides with N-terminal extensions up to 40 residues [141, 142], aspartic acid isomerization [143], and phosphorylation [144, 145].
AβO assembly pathways and their relation to amyloid plaques
A preponderance of data now supports the hypothesis that some AβO species are “on-pathway” to fibril formation, while others are “off-pathway”. It is these “off-pathway” oligomeric species that may be the most toxic [146]. This on/off-pathway classification appears to correlate with the HMW/LMW and type 1/2 AβO classifications discussed above. Most data show that LMW, type 2 AβOs are on-pathway to fibril formation, while HMW, type 1 AβOs are off-pathway [118, 148]. High-speed AFM imaging demonstrates that LMW AβOs quickly form fibrils, whereas HMW do not [147]. These aggregation differences between LMW and HMW AβOs are consistent with earlier findings using SDS-PAGE analysis [118]. In fact, it seems as if the only contribution of HMW AβOs to fibril formation may be through their dissociation into LMW AβOs, which then seed fibril formation [147]. Interestingly, differences in the aggregation pathways of these two AβO structures occur as early as the dimer stage [149]. But contrary to the hypothesis of HMW AβOs being more toxic than LMW AβOs, one study has found that HMW AβOs are not as toxic as the LMW AβOs into which they dissociate [150]. And another study utilizing all-atom molecular dynamics simulations observed that compact AβO structures, with an oligomeric order up to 18 (81 kDa), are off-pathway to fibril formation, while larger, elongated AβO structures are on-pathway to fibril formation [149]. Therefore, although there is general agreement in the literature regarding toxicity of AβO species, there is not complete consensus.
An alternate hypothesis to the on/off-pathway model of AβO formation, is the fibril-seeded model [151]. In this model, toxic AβOs are predominantly formed from monomers that dissociate from fibrils only after a small, but critical concentration of fibrils has formed. This model is supported by kinetic experiments, selective radiolabeling experiments, and cell viability assays. Further support for secondary nucleation of AβOs comes from molecular dynamics simulations. These simulations predict that a hydrophobic fibril region causes the structural changes required for catalyzing the formation of AβOs on the fibril surface [152]. However, AβOs can form within minutes in vitro, even at low nanomolar concentrations [26]. Recent AFM studies confirm that AβOs can form within minutes [153]. This quickly forming AβO population is specifically dominated by hexamers and dodecamers and quickly followed by AβO-seeded fibril formation. Therefore, one factor that has led to contrasting conclusions regarding the timing of AβO primary versus secondary nucleation pathways may be the differing time resolutions of the different experimental techniques.
Another hypothesis, the amyloid plaque buffering hypothesis, supports this notion of co-existing primary and secondary nucleation pathways for AβOs. This hypothesis predicts that plaques act as a reservoir or sink for toxic AβOs [107]. AβOs gradually deposit as diffuse plaques, which cause inflammation, but AβOs also can directly cause damage leading to dementia via altered signaling [5, 110]. Evidence for AβOs existing in these diffuse plaques comes from immunofluorescent imaging with anti-AβO antibodies. This has been observed in the AD brain and in the brains of multiple animal models [7, 155]. Over time, this plaque reservoir is saturated or loses capacity and toxic AβOs become free to diffuse and exert toxicity [107, 157]. Overall, it seems as if evidence in the literature converges into the hypothesis that Aβ aggregates into distinct AβO species, with differing toxicities and relationships to fibrils, that can interconvert.
Emerging insights into how to approach molecular structure
A major hurdle to AβO structural characterization is AβO metastability and heterogeneity. One major approach to stabilize and isolate distinct AβO species has been crosslinking. One widely applied crosslinking method for AβO stabilization has been photo-induced crosslinking (PICUP), developed by the Teplow group. Initially, this method was successful in stabilizing only LMW oligomers of the Aβ40 peptide [158]. Recently, PICUP has been improved through use of the mutated Aβ42 peptide [F10, Y42]Aβ42, enabling stabilization of Aβ42 oligomers up to dodecamers [159]. Another crosslinking strategy used for AβO stabilization is dityrosine crosslinking. This method is thought to be AD-relevant as it occurs under conditions of elevated copper and oxidative stress [160]. Copper is relevant to AD as there is some evidence that dyshomeostasis of metals, including copper, may contribute to AD pathogenesis [161]. Furthermore, dityrosine crosslinked proteins are found to be prevalent in AD brain and CSF [160]. Molecular dynamics simulations predict that dityrosine crosslinking promotes Aβ self-assembly, at least up to tetramers [162]. In one study, copper was found to stabilize Aβ in an oligomeric conformation sufficiently to enable 3D structural characterization by small-angle x-ray scattering [163]. The putative mechanism of this copper-mediated stabilization was through copper-induced dityrosine linkage of Aβ peptides [164]. Different copper ratios had different effects on Aβ aggregation, with supra-equimolar ratios favoring formation of ellipsoid oligomers and sub-equimolar ratios favoring formation of fibrils [163]. These ellipsoid AβOs were predicted to contain 38 copies of the Aβ peptide and are therefore consistent with the converging classifications of off-pathway, HMW, type 1 AβOs. Given published findings, it is essential that AD-relevant stabilization techniques continue to evolve to enable direct structure-function comparisons of distinct AβO species under AD-relevant experimental conditions. This will make it possible to properly interpret Aβ-directed clinical findings and make the most informed efforts at rational design of AβO-targeting therapeutics.
What makes AβOs toxic to neurons?
AβOs can be extracellular in vivo, existing in CSF [32, 165] and in interstitial fluid [166]. Some brain cells when exposed experimentally to extracellular AβOs become dysfunctional and deteriorate, as reviewed above. How AβOs instigate pathological changes, and why only some cells are affected, are fundamental questions to which we do not yet have satisfying answers.
The simplest possibility is that cell damage is initiated by physiochemical interactions between AβOs and neuronal membranes. It has been reported that AβOs can insert directly into lipid bilayers, causing disruption by acting as a pore, a phenomenon first observed with artificial membranes [167]. The extensive amount of literature concerning Aβ-lipid membrane interactions and molecular level membrane modeling has recently been reviewed [168], including the possible involvement of metals in the mechanism [169]. AFM was used recently to show structural damage to POPC/POPS lipid bilayers caused by Aβ40 in different aggregation states [170]. Aggregation and lipid interaction properties of Aβ peptide fragments incubated in the absence or presence of total brain lipid extract bilayers indicate that some sequences interact with and disrupt bilayers (e.g., Aβ40) but others do not (e.g., Aβ28 and Aβ12–24) [171]. Some experiments indicate that oligomers of Aβ have more membrane affinity than monomers [172]. Putative oligomers from Aβ that is pyroglutamate-modified also binds neurons and causes a loss of plasma membrane integrity [173]. Ion channel formation in cell membranes [174] has been reported for oligomers of Aβ42, but not Aβ40, and attributed to a pore-forming beta-barrel AβO structure [175]. Individual AβOs larger than trimers reportedly induce Ca++ entry as they cross the cell membrane [176]. Cholesterol enhances formation of an annular octameric channel of Aβ22–35, which induces a zinc-sensitive Ca++ influx [177], suggested as a possible lipid raft association. Recruitment of AβOs to rafts is consistent with findings that depletion of the ganglioside GM1 blocks AβO interactions toxicity [178]. On the other hand, data suggest that a moderate increase in membrane cholesterol content may be protective against AβO toxicity [179]. Protection also is conferred by a pentapeptide from the glycine zipper region of the C-terminal of Aβ, which blocks apparent membrane insertion and abolishes synaptotoxicity [180].
One significant difficulty encountered by the bilayer insertion hypothesis is its inability to account for the specificity of AβO attachment. Two neurons side-by-side can exhibit completely different ability to accumulate AβOs, one showing robust synaptic accumulation and the other showing virtually none [16]. Cell-specific toxicity, measured by tau hyperphosphorylation, correlates with this binding [51]. There also is a difficulty in accounting for binding saturability [16, 96], although it might be argued that AβO insertion into lipid rafts specific to particular synapses could be saturable. It has been hypothesized that AβOs may act through both lipids and proteins, forming pores within membranes while also binding to receptors to induce specific intracellular responses [181].
The receptor hypothesis regards AβOs as gain-of-function pathogenic ligands that bind adventitiously to specific proteins acting as toxin receptors. Overall, the receptor hypothesis provides a mechanism that fits well with salient facets of the cell-based evidence. The hypothesis was introduced to explain toxicity that was cell-specific and dependent on expression of Fyn, and to explain why AβO binding was virtually eliminated by treating cell surfaces with low doses of trypsin [26]. Consistent with the receptor hypothesis, AβO binding shows (A) saturation and high-affinity for cultured neurons and synaptosome preparations; (B) specificity for particular neurons and particular brain regions; (C) targeting of synapses; (D) accumulation at dendritic spines; (E) pathological impact, such as tau hyperphosphorylation, specific to neurons with bound AβOs; (F) sensitivity to low doses of antagonist; (G) binding to trypsin-sensitive proteins; (H) association with small patches of isolatable membranes; and (I) specificity in Far-Western immunoblots [6, 182]. These findings apply generally to brain-derived and synthetic AβOs and support the hypothesis that binding of AβOs is ligand-like and mediated adventitiously by proteins acting as toxin receptors.
Perhaps the most intriguing and well-studied AβO toxin receptor candidate is the cellular prion protein (PrPc). Strittmatter and colleagues in a series of papers have provided strong evidence that PrPc is capable of mediating AβO binding [122, 183–185], starting with their unbiased screening of a cDNA expression library that identified PrPc as a potential high-affinity AβO receptor [186]. Extensive experiments with multiple models support this possibility and connect binding to neural damage [187–190]. It has been reported that binding of AβOs to PrPc is dependent on integrity of cholesterol-rich lipid rafts and that AβOs bound to PrPc accumulate in endosomes after which they are trafficked to lysosomes [191]. Investigations of how externally-oriented PrPc might bring about intracellular damage through transmembrane coupling to Fyn are discussed further below. Coupling of AβO-bound PrPc to Fyn is consistent with the original studies showing that Fyn expression is required for AβO-induced toxicity [26, 192] and evidence showing involvement of Fyn in physiological PrPc signaling [193]. It should be noted that the PrPc hypothesis is still somewhat controversial, and some reports are not in agreement with the role of PrPc as an AβO toxin receptor [29, 194–197].
Another promising candidate receptor is the Na+K+ ATPase alpha3 subunit (NKAα3), recently identified independently by two groups using disparate preparations and identification strategies. It was shown first by Ohnishi and colleagues that NKAα3 can bind both brain-derived and synthetic AβOs [198], each resembling type 1 AβOs with respect to their relatively large size. Verification of NKAα3 as an AβO receptor subsequently was provided by DiChiara et al. [199]. This group used solubilized synaptic membrane proteins reconstituted in nanoscale artificial membranes and an AβO-specific antibody to isolate AβO-bound NKAα3. Co-localization of AβO binding sites with NKAα3 was confirmed in hippocampal cell cultures. As discussed later, down-regulation of NKAα3 could play a significant role in converting AβO binding into cell pathology.
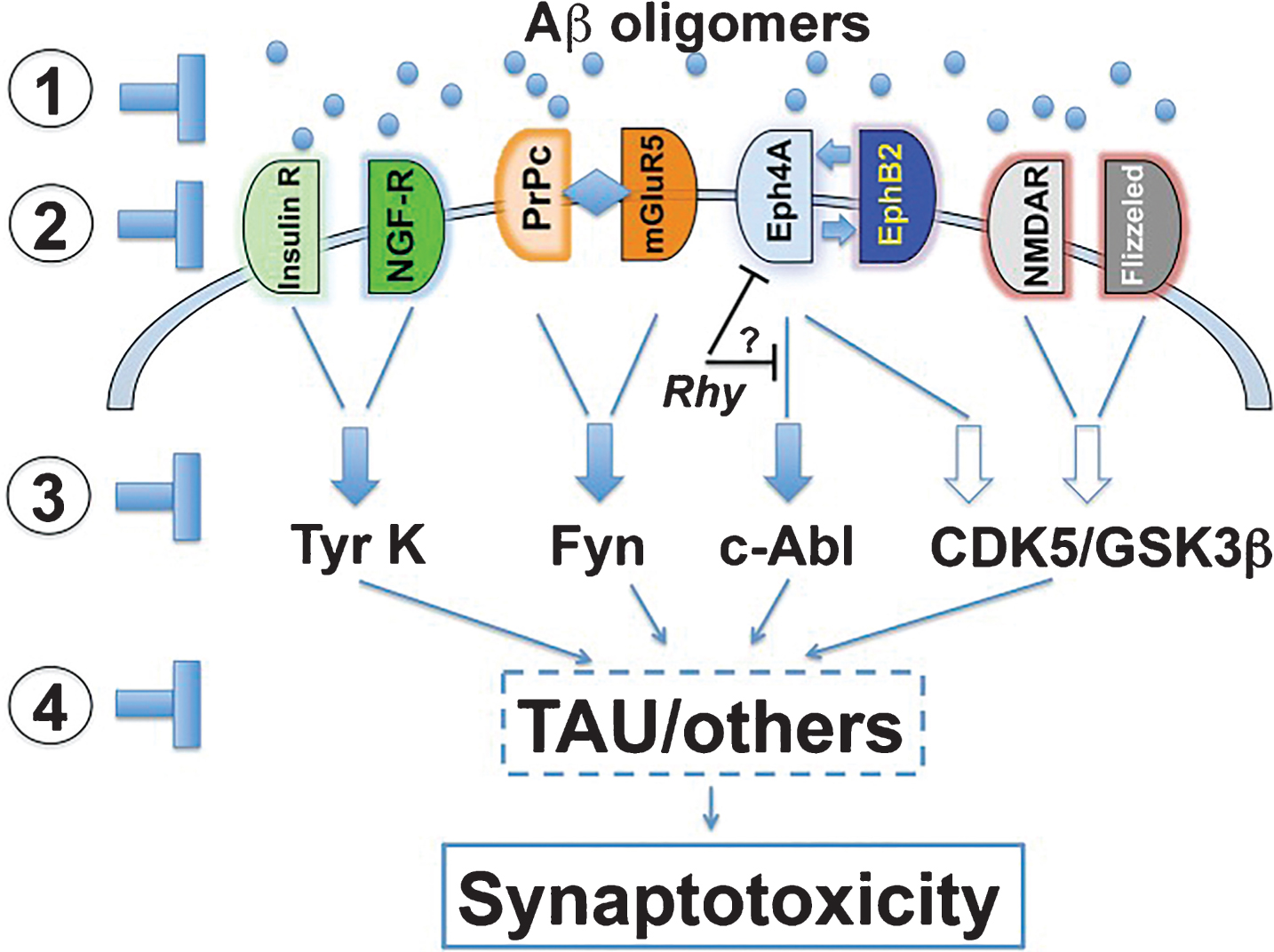
Overall, and rather remarkably, the current list of candidate toxin receptors for AβOs comprises a very large number of membrane proteins besides PrPc and NKAα3. As has been reviewed [115, 200–202], these include the metabotropic glutamate receptor 5 (mGluR5) [182, 184], NMDARs [58, 62], Sigma-2 receptor/progesterone receptor membrane component 1 (PGRMC1) [203, 204], frizzled receptor [205], neuroligin [206], Ephrin type-B receptor 2 (EphB2) [207], Ephrin type-A receptor A (EphA4) [208, 209], p75 neurotrophin receptor (p75NTR) [210], alpha7-nicotinic acetylcholine receptor (α7nAChR) [211, 212], adrenergic receptors [213], the receptor for advanced glycation endproducts (RAGE) [214], calcium channels [215–217], leukocyte immunoglobulin-like receptor subfamily B member 2 (LILRB2)/paired Ig-like receptor B (PirB) [64, 219], N-formyl peptide receptor 2 (FPR2) [220], immunoglobulin gamma Fc region receptor II-b (FcγRIIb) [221], transient receptor potential melastatin 2 (TRPM2) [222], insulin receptor (IR) [48], and the AMPA receptor [223].
It is not known why there are so many candidate receptors. There certainly are different forms of AβOs, which could interact with different membrane proteins. AβO ligands in aqueous buffer are between 100 and 300 kDa (Cline and Klein, unpublished); in this mass range AβOs would comprise 22–66 Aβ monomers. To interact with a binding domain of a toxin receptor, only particular regions of the oligomer surface would be needed. Ligand-like regions could assume multiple configurations influenced by buffer composition. For example, monomeric Aβ in Ham’s F12 media assembles into structures quite different than Aβ in phosphate buffered saline (the former has a high type 1 to type 2 ratio [118], while the latter has a low type 1 to type 2 ratio) (unpublished data). Even within a population of synthetic type 1 AβOs, there is a small subpopulation of synapse-binding AβOs that can be targeted uniquely by a selective single-chain variable fragment antibody [118]. Different targeted binding proteins might nonetheless mediate similar changes downstream. It is known, for instance, that AD-type phosphorylated tau can be induced by oligomers of different proteins such as Aβ [51], α-synuclein [224], and even lysozyme [225].
Receptor transduction mechanisms → how does the initial receptor-AβO interaction on neurons trigger a change that leads to intracellular damage?
The mechanism by which PrPc mediates AβO impact intracellularly has been carefully worked through (Fig.6). It incorporates a number of molecular players previously implicated by multiple laboratories: mGluR5 [182, 184], Fyn [26], tau [51], NDMARs [58, 62] and protein tyrosine kinase 2 (Pyk2) [226]. Both high and low molecular weight AβOs have been implicated in this pathway [121–123, 191]. Data are consistent with a mechanism in which AβOs first bind to PrPc on cell surfaces and stimulate Fyn via mGluR5 activation (reviewed by Nygaard, et al. [227]). Consistent with activation of mGluR5 by AβOs, the ability of glutamate to activate the prion-mGluR5 complex is occluded [228]. Downstream, stimulated Fyn is known to phosphorylate tau [229] and cause tyrosine phosphorylation of the NR2B subunit of NMDARs [183]. It is thought that AβO binding to neurons and AβO neurotoxicity depends on a pre-existing PrPc-mGluR5 complex [230]. However, since PrPc can be removed with full retention of AβO binding [194], it may be that the critical membrane-organizing function of PrPc precedes the ligand binding step.
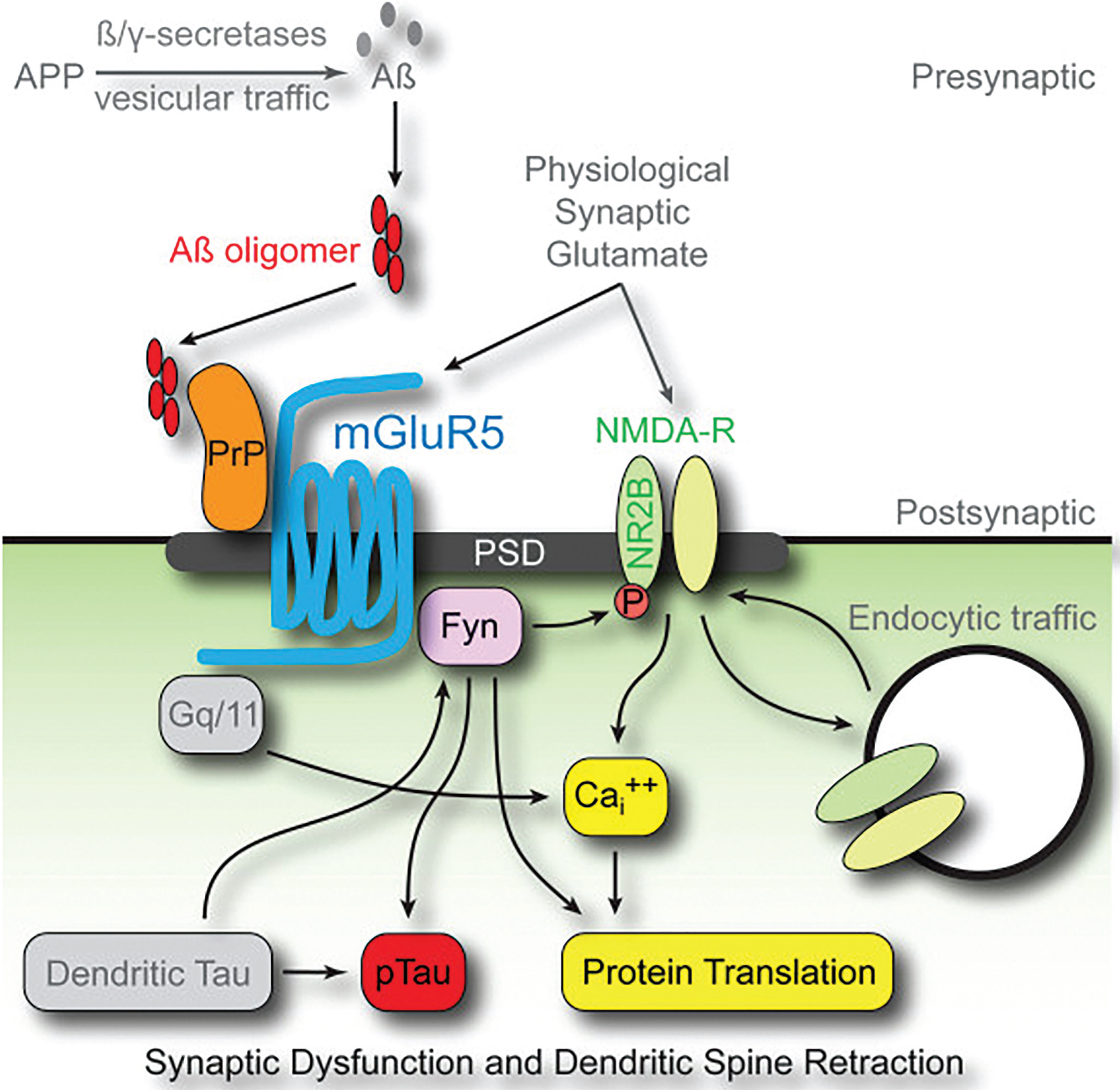
An interesting potential connection exists between this synaptopathic mechanism and Pyk2. Pyk2 has a single nucleotide polymorphism identified as increasing the likelihood of late-onset AD [231]. Functionally, Pyk2 is a focal adhesion kinase (FAK), an enzyme that previously was shown to be stimulated by toxic Aβ preparations and to form complexes with Fyn [226]. Pyk2 normally helps regulate synaptic plasticity [232–235]. It is activated by increased intracellular calcium and by Fyn [236–241]. Ectopic activation of Pyk2 potentially could be an early event in this AβO pathogenic pathway, which would provide a molecular basis for its risk factor status.
Insights also have been obtained into how AβO binding mediated by NKAα3 could be transduced into neuronal damage. As described by Ohnishi et al. [198], AβO binding leads to a slow, time-dependent decrease in ATPase activity. The consequence is Ca++ buildup via N-type voltage-sensitive calcium channels (N-VSCC) and mitochondrial channels and ultimately apoptosis. Decrease in activity was suggested as linked to AβO binding to the ouabain binding site of the NKAα3 [198]. This observation suggests the new hypothesis that, under some circumstances, AβOs could be endogenous ouabain-like physiological regulators of ATPase. The slow, time-dependent decrease in activity, however, could be linked to the observed impact of AβOs on NKAα3 distribution. Following exposure to AβOs, neuronal NKAα3 accumulates in dense clusters along dendrites. These clusters of NKAα3 increase in size and then decrease in abundance (Fig.7) [199]. This presumably occurs at dendritic spines, where AβOs also cluster [16, 55]. Like the NKAα3 redistribution, spines undergo time-dependent changes in morphology and abundance due to AβO exposure. Ultimately, there is a large down-regulation of NKAα3, which could account for decreased ATPase activity (Fig.7).
The issue of distribution is a salient one given that NKAα3 acts not only in cation transport, but also as a membrane protein docking station that functions to control signaling pathways [242]. These docking stations organize multiple membrane proteins [8], including neurotransmitter receptors linked to AβO-induced neuron damage [243]. The clustering of NKAα3 is in harmony with the earlier observation that AβOs induce the clustering of mGluR5 [182, 184]. As discussed above, mGluR5 is a Ca++ mobilizing receptor, and it is regarded as a key mediator of AβO-elevated Ca++ buildup and the damage that ensues [184]. The time-dependence of AβO-induced clustering of mGluR5 has been imaged using quantum dots and single-particle tracking in experiments with live neurons [182]. It has been hypothesized that mGluR5 clustering itself is a seminal step for the transduction mechanism [182]. Supporting this possibility, clustering of mGluR5 molecules induced by receptor antibodies mimics the toxic impact of AβOs [182]. Because mGluR5 and NKAα3 each co-localize with cell-surface bound AβOs, they likely are part of the same ectopic clusters. Recently, single-particle tracking experiments have shown that NKAα3 becomes immobilized during exposure of hippocampal neurons to toxic assemblies of synuclein [244]. These results support the hypothesis that NKAα3 has a central role as an immobilizing docking station for toxic oligomers found in multiple proteinopathies. With respect to generation of these clusters, the role of the NKAα3 docking station relative to roles played by mGluR5, or other membrane domain-organizing proteins such as PrPc [245], is not yet clear.
The seminal interactions between AβOs and NKAα3 molecules at the cell surface may be suitable targets for new drug discovery strategies, as suggested by Ohnishi et al. [198]. Attachment of AβOs to NKAα3 is amenable to high-throughput screening for antagonists using Nanodiscs [194, 246]. Results from a preliminary screen showed that AβO binding to spines can be blocked by low doses of a small organic molecule, albeit one with promiscuous binding, precluding its use for therapeutics [194]. Nonetheless, Lee and colleagues have shown that behavior in a Tg AD model could be safely rescued using this same compound [247]. Future investigations of the docking station hypothesis are expected to open the door to therapeutics targeting the first step of a complex pathway that leads to neural damage and dementia.
Whether the NKAα3 acts, as has been suggested [198], as a “death target” for AβOs is not confirmed yet. Most AD-like pathology is evident in cultures containing almost exclusively neurons, but cell death is minimal; neuron death likely requires the presence of factors released by glia [248]. It is possible that the impact of AβOs on NKAα3 may render them more vulnerable to inflammatory cytokines.
TRENDING TOPICS IN AβO RESEARCH
With about 2,000 AβO papers published since 2013, there has been a great deal of progress on many issues. Some of the salient directions are considered briefly in this section.
Toxic effector pathways after initial transduction
Downstream, after the initial transduction steps, the impact of AβOs has been tracked to mitochondrial effects, ER stress, and autophagy/lysosomal dysfunction. These may be the consequences of surface events discussed above, but some studies show that AβOs may themselves enter cells and act directly on these organelles, as discussed below.
AβO-associated NMDAR activation [62] promotes Ca++ release from the ER, which leads to ER stress [249] with subsequent mitochondrial dysfunction [250], astrogliosis [115, 251], and apoptosis [18]. AβOs also have been found to trigger the unfolded protein response, a collection of signaling pathways that respond to ER stress [252]. AβOs further decrease resistance of brain mitochondria to Ca++-induced opening of mitochondrial permeability transition pores [253]. Cytochrome C is released by AβO-activated BAK pores [254]. Voltage-dependent anion channel 1 also interacts with Aβ monomers and oligomers, and the block of mitochondrial pores leads to mitochondrial dysfunction [255]. Morphological effects on mitochondrial fusion and fission dynamics, essential for neuronal function, have been reviewed [256]. AβO targeting of mitochondria promotes mitochondrial fission, disruption of mitochondrial membrane potential, increase of intracellular ROS and activation of mitophagy [257]. AβOs decrease mitochondrial volume [258], and AβO-induced oxidative stress is associated with mitochondrial fission [259]. AβOs activate fragmentation through the GTPase dynamin-related protein 1 (Drp-1) [260] and extracellular signal-regulated kinase (ERK) [259]. Fragmentation also has been associated with AβO-induced mitochondrial transport defects, with histone deacetylase (HDAC6) activation part of the mechanism [260]. AβO-induced mitochondrial damage appears to be restricted to neurons and not microglia or astrocytes [261].
With respect to autophagy, the sole catabolic mechanism for degrading protein aggregates, there is increasing evidence for autophagic dysfunction in AD and other neurodegenerative diseases [262, 263]. The endosomal-lysosomal (autophagy) system is a prominent site of AβPP processing, Aβ uptake, and Aβ production [262]. One study has found that AβOs associate with autophagic vacuoles in AD axons, starting a pathway that impairs retrograde transport, which contributes to autophagic stress [264]. On the other hand, another study found that it is Aβ monomers, and not AβOs, that contribute to autophagy [265]. AD and lysosomal storage disorders share many overlapping pathologies, including neuronal accumulation of lysosomal vesicles, dystrophic axons, ectopic dendrites, cognitive deficits, and neurodegeneration [262]. Lysosomal storage disorder gene variants also have been found to be associated significantly with Parkinson’s disease [266]. Restoration of autophagy function may represent a promising therapeutic target as rifampamycin, a candidate preventative therapeutic thought to restore autophagy function, has been found to inhibit oligomerization of Aβ and tau, tau phosphorylation, synapse loss, and microglial activation in AD mouse models [267].
Intracellular effects of AβOs may be instigated by surface mechanisms but also could be a result of direct interactions between organelles and internalized AβOs. In NHPs, i.c.v.-injected AβOs were observed on the surface and inside neurons [46]. Internalization may involve signaling pathways that affect regulation of receptor-mediated endocytosis. In the human neuroblastoma SH-SY5Y line, AβOs activate p38 mitogen-activated protein kinase (p38 MAPK) and ERK1/2 signaling pathways via the α7nAChR, which in turn results in AβO internalization [268]. Internalized Aβ (monomers and AβOs) localized to all organelles (ER, Golgi complexes, multivesicular bodies/late endosomes, lysosomes, exocytotic vesicles, and mitochondria) and non-membrane-bound cytosolic structures [269, 270]. The uptake of Aβ via endocytosis is rapid and spontaneous. It is retained in lysosomes, where accumulation leads to aggregation [271].
The relationship between neurons, astrocytes, microglia, and AβOs
De Strooper and Karran propose that AD pathogenesis is not simply a neuron-centric, linear cascade initiated by Aβ and leading to dementia, but rather a long, complex cellular phase consisting of feedback and feedforward responses of astrocytes, microglia, and vasculature [202].
Many lines of evidence support this hypothesis. For instance, AβOs have been found to induce astrogliosis [272] and trigger ROS generation in activated astrocytes [273]. AβOs reportedly cause disturbances in the signaling of insulin, protein kinase B (Akt), and excitatory amino acid transporters 1 and 2 [274]. Decreases in the activation and expression of astrocytic glutamate transporters has been linked to impaired synaptic plasticity [275]. AβOs at picomolar levels, within minutes, can increase levels of intracellular Ca++ in astrocytes but not neurons [276]. An increase in ROS production by nicotidamine adenine dinucleotide phosphate (NADPH) oxidase in both neurons and astrocytes has been found to activate caspase-3, also linked to LTP inhibition. In these experiments, only a small fraction of AβOs were impactful and their damage was blocked by clusterin [276]. AβOs and fibrils bind and activate Ca++-sensing receptors, which drives both neurons and astrocytes to secrete Aβ42. While the Aβ-exposed neurons start dying, astrocytes survive, and they keep over-secreting Aβ42, nitric oxide (NO), and vascular endothelial growth factor A (VEGF-A), apparently contributing to the demise of neurons [277]. On the other hand, astrocytes, before they are affected by AβOs, appear to release insulin and insulin-like growth factor (IGF1), whose trophic effects serve to protect neurons from AβO toxicity [73].
Microglia in AD are involved in phagocytosis of Aβ plaques [278–281], a process that is regulated by astrocytes [278]. It is possible that AβOs play a role in attracting microglia to plaques (Bicca and Klein, unpublished data). Besides chemotactic effects, AβOs induce a switch in microglial phenotype to a pro-inflammatory phenotype, leading, e.g., to aberrant tumor necrosis factor (TNF) signaling [282]. Aberrant TNF signaling causes decreased brain serotonin levels and subsequent depression [283]. It also causes insulin receptor substrate (IRS-1) and PKR (dsRNA-dependent protein kinase)-dependent synaptic dysfunction and memory loss [221]. There thus is a link between AβOs, neuroinflammation, mood alterations, metabolic disorders, and memory loss. Microglia also may contribute to AβO formation, by releasing particles that can bind rapidly to Aβ and cross-seed its aggregation, including oligomerization [284]. It should be noted that there also is evidence that microglia may contribute to neuronal loss and memory impairment in a manner independent of Aβ [281]. One potential mechanism is through microglia engulfment of synapses [285].
Tau progression and prion-like action may be instigated and potentiated by AβOs: PART (primary age-related tauopathy)
Most evidence in the literature converges on the hypothesis that AβOs are upstream of tau in AD pathogenesis and not the other way around, as reviewed by Bloom [286]. However, there is currently no consensus in the field, with some studies demonstrating crosstalk between AβOs and tau and some demonstrating that each acts separately [110, 286–290]. In support of the hypothesis that AβOs trigger tau pathology, it was demonstrated in 2008 that AβOs were capable of inducing tau hyperphosphorylation in cultured neurons in the absence of fibrils [51]. Tau distribution in AβO-exposed neurons ectopically redistributes to dendrites [52]. AβOs also have been shown to induce tau-dependent microtubule severing [291], to disrupt tau translocation to excitatory synapses [292], and to stabilize microtubules, the latter leading to tau-dependent loss of spines and tau hyperphosphorylation [293]. AβOs even can seed the formation of tau oligomers, which are thought to be the most toxic form of tau [294]. In the AD brain, synaptic AβOs have been found to precede synaptic phosphorylated tau (pTau), even perhaps driving the synaptic spread of pTau [295]. It is known that spread of tau pathology in a Tg mouse tauopathy model is accelerated by crossing with an APP Tg mouse [296]. Recent data suggests that AβOs may induce neurons to release pTau within exosomes, thereby suggesting a potential mechanism for AβO-induced spread of tau pathology [73]. Interestingly, this release of tau was increased by the presence of insulin. The idea that tau is secreted by neurons is supported by numerous other studies, as reviewed by Pooler et al. [297]. Furthermore, i.c.v. injection of AβOs into NHPs induces tau hyperphosphorylation and formation of neurofibrillary tangles throughout the NHP brain [46].
Many recent studies have attempted to give Aβ and tau an even playing field on which to determine their pathological relationships by crossbreeding mice expressing human tau (wild-type or mutant) and APP/PS1 mutants. However, these studies show inconsistencies in data leading to contrasting conclusions. Co-expression of mutant tau and mutant Aβ appears to support a synergistic action, showing dramatically increased pTau aggregation and spread, inflammation, and synapse loss [296, 299]. Multiple studies utilizing a wild-type human tau instead of mutant tau also support a synergistic model, for example [300, 301]. Although it may be that some aspects of AD pathology are cooperatively affected by Aβ and tau, while others are independently affected [155, 302]. Contrary to these studies, one report found no evidence for pathological interaction between Aβ and tau [303]. These apparently disparate findings may be the result of utilizing different transgenes and/or pathological readouts.
AβOs themselves as prions?
The idea that oligomers of amyloid proteins, including AβOs, may spread from neuron-to-neuron in a prion-like manner has been widely considered. Although there is currently no clear clinical evidence that AD is a transmissible prion-like disease [5], experimental data support the idea that AβOs may spread from cell-to-cell and brain region-to-region in a prion-like manner. A recent review of this hypothesis states that Aβ aggregates have all of the key characteristics of canonical mammalian prions, including a β-sheet rich architecture, the tendency to polymerize into amyloid, templated corruption of like protein molecules, the ability to form structurally and functionally variant strains, the systematic spread by neuronal transport, and resistance to inactivation by heat and formaldehyde [304]. Another review of this concept predicts that small, extracellular oligomers of amyloid proteins would have a high propensity for prion-like spread, while large intracellular oligomers would have a lower propensity for prion-like spread [305]. In support of AβOs acting as prions, one study has found that AβOs can transfer from cell to cell [306]. This transfer was shown to be dependent on insufficient cellular clearance of Aβ peptides and oligomers. The remaining un-degraded Aβ was able to cause seeding and pathology in the receiving cells. Cell transfer was an early event seemingly independent of later toxicity. Aβ can seed its own aggregation in vitro [307, 308] and brain extracts from AD patients and animal models can seed Aβ aggregation in vivo [309, 310]. Furthermore, i.c.v. injections of AβOs to NHPs induced accumulation of AβOs in specific brain regions far from the injection site, suggesting spreading [46]. Hypotheses for the mechanisms of Aβ spread include exosome transfer [311] and spread directed by the limbic connectome [312, 313].
Mechanisms of buildup
Three intriguing new hypotheses for the mechanism of AβO accumulation that have emerged in the literature in the past 5 years are saturated proteostasis, shear-induced amyloid formation, and slowed clearance of AβOs from interstitial fluid. These hypotheses are briefly reviewed below.
Saturated proteostasis
One theory to explain the accumulation of Aβ aggregates is saturated proteostasis [314]. This theory, based on a large body of evidence, states that there may be nothing particularly unique about the Aβ42 peptide that causes it to aggregate into toxic oligomers and amyloid fibrils. In fact, this may be an ability inherent in all proteins if they are placed in the right conditions. There is increasing evidence that many proteins are kinetically, but not thermodynamically, stable in their native states and that they become metastable when their cellular concentrations exceed their critical values. Considering the specific example of Aβ42, one study found that changing the propensity of this peptide to aggregate by only 15% through site-directed mutagenesis resulted in large changes in toxicity [315]. The authors interpret this finding to mean that Aβ42, and other amyloid proteins, may be extremely close to their solubility limit under physiological conditions. Thus, they hypothesize that in AD, or other neurodegenerative diseases associated with misfolded proteins and aging, age-related stress makes the entire proteome susceptible to aggregation, which in turn saturates the protein quality-control system of the cell [314]. Indeed, the majority of proteins implicated in AD were found to be present at supersaturated concentrations in the cell [316]. Therefore, proponents of this saturated proteostasis theory suggest that more effective therapeutics may target the driving forces for whole-proteome aggregation and protein quality-control mechanisms instead of individual disease-related proteins like Aβ [314]. More specifically, the systems that were found to be of importance in maintaining proteostasis in AD were involved in trafficking and clearance mechanisms, including specific branches of the endosomal-lysosomal and ubiquitin-proteasome systems [317].
Shear-induced amyloid formation
Another new hypothesis to explain the etiology of AβO buildup is the shear-induced amyloid formation hypothesis [318]. This hypothesis predicts that Aβ within the slow-flowing interstitial fluid can gain significant shear energy at, or near, the wall of the narrow extracellular spaces of the brain parenchyma. This could cause Aβ to absorb to the brain membrane and form oligomers on the membrane and/or form plaques within the flow pathways of the brain extracellular spaces.
Slowed clearance of AβOs from interstitial fluid
Microdialysis experiments have shown the presence of AβOs in interstitial fluid [166]. These findings are an extension of earlier studies showing a circadian rhythm in interstitial Aβ levels [319]. Clearance through the glymphatic system is inversely correlated with AβO size [166]. Impaired glymphatic functioning is considered to be a likely factor in AβO accumulation [320, 321] (see discussion below).
Etiological factors that trigger AβO buildup in sporadic AD
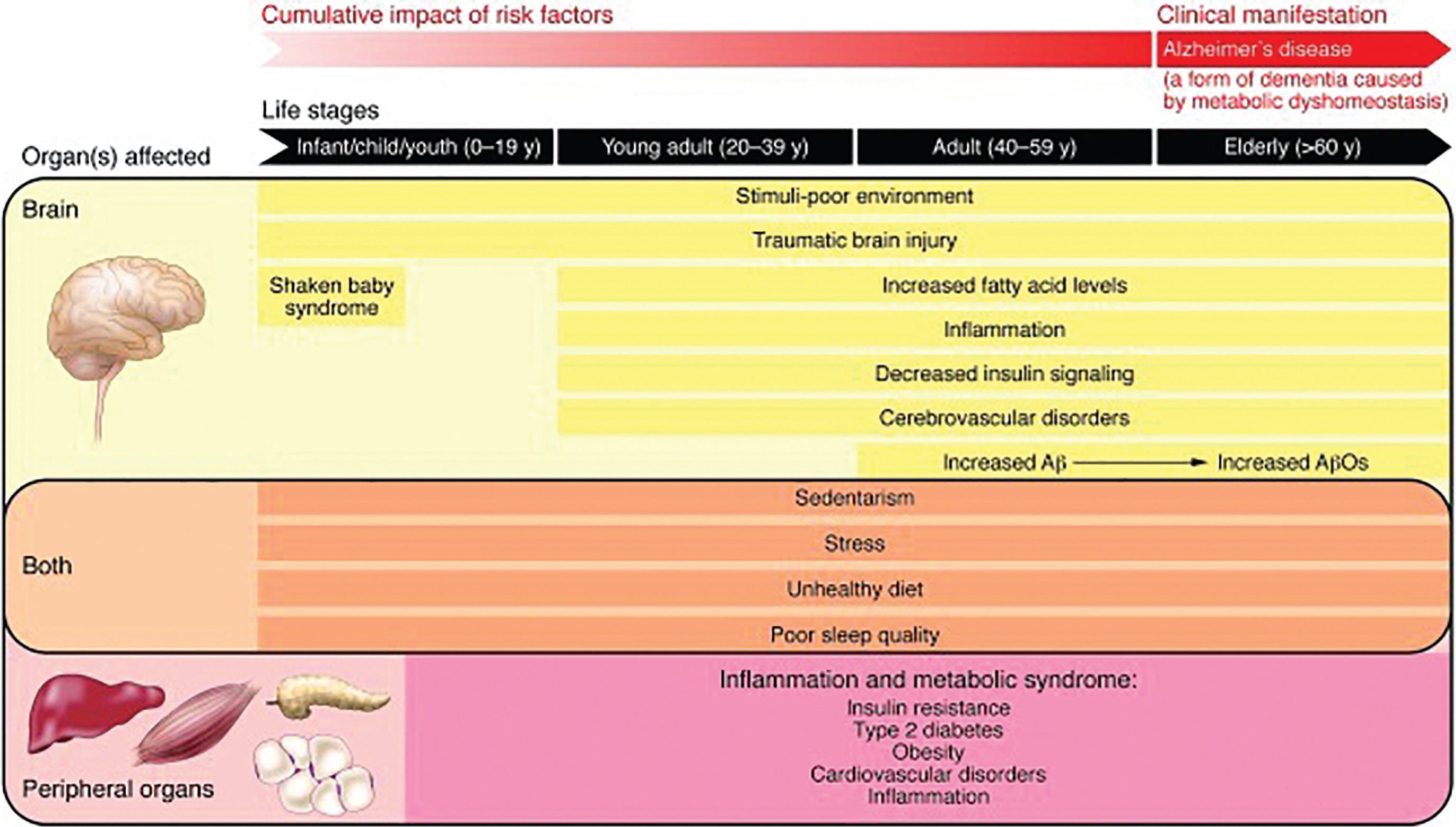
There is evidence that traumatic brain injury (TBI), atmospheric pollutants, poor quality of diet and sleep, and metabolic diseases (e.g., type 2 diabetes and hypercholesterolemia), may trigger AβO buildup, eventually leading to non-inherited forms of AD (sporadic) (see, e.g., [322]). A hypothesis from De Felice for the contributions of these etiological factors to AβO buildup and AD is illustrated (Fig.8) [322]. Evidence implicating each of these factors in AβO buildup is briefly reviewed below.
TBI
TBI is a risk factor for AD [323] with AD developing in 55.5% of TBI patients [324]. Aβ pathology has been found to accumulate in the brain and CSF following TBI, including amyloid plaques [325] and AβOs [199, 326–328]. Soluble Aβ levels, including AβOs, increase with TBI severity [327] and declining patient prognosis [328]. Results are consistent with indications that AβPP expression is injury-related, e.g., in shaken-baby syndrome [329–331]. These observations are supported in TBI animal models, wherein Aβ levels rise within 1 hour after a single mild cortical impact and continue to rise for at least 24 hours [332, 333] and are associated with increased memory impairment [334]. AβO-associated proteins, PrPc and pTau, also are increased in TBI mouse models [335].
Atmospheric pollutants
Recent studies in mice have demonstrated that air pollutants, specifically vehicular-derived airborne nano-sized particulate matter, induce AD-like neuronal damage, including reduced synaptic function [336], altered neuronal differentiation and depression-like responses [337], and reduced neurite outgrowth [338]. Two AD risk factors, age [339] and gender (female) [337], appear to increase susceptibility to these detrimental effects.
Poor quality of diet and sleep
Diets high in sugar, salt, and fat and low in fruits and vegetables are associated with a higher risk of AD [340]. In animal models, diets high in fat increase soluble Aβ without changing plaque burden [341] and diet-induced insulin resistance impairs cognition [342]. In humans, such diets have been shown to perturb the circadian modulation of cortisol secretion, which is associated with poor sleep quality. Poor sleep quality also is associated with dementia and can negatively affect glymphatic system activity, which leads to Aβ accumulation via impaired clearance (see discussion below) [340]. Furthermore, sleep restriction in mice promotes neuroinflammation and synapse loss and potentiates AβO-induced memory deficits [343].
Diabetes
Sporadic AD has been called type 3 diabetes for its molecular and biochemical similarities with type 1 and 2 diabetes [322, 344]. An increasing body of evidence shows that AD is coupled to impaired brain insulin signaling, glucose utilization, and energy metabolism, all of which lead to increased oxidative stress, neuroinflammation, and further increased insulin resistance. Specifically considering AβO buildup, it has been found that glucose concentrations observed in diabetics facilitate Aβ oligomerization [345]. Furthermore, induction of diabetes in rabbits leads to AβO accumulation in the brain and retina [346]. Most recently, type 2 diabetes has been found to be positively associated with Aβ42 in CSF [347]. The mechanism for AβO buildup in diabetes is not known, but it has been hypothesized to be mediated by inflammation [322].
Hypercholesterolemia
Hypercholesterolemia also is an AD risk factor [348, 349]. Many studies have shown that elevated cholesterol levels may contribute to AD pathogenesis, and several cholesterol-related gene polymorphisms are associated with AD, the most well-known of which is APOE [349]. Hypercholesterolemia accelerates AβO accumulation and memory impairment in AD mouse models [350, 351].
It is important to note that even though these factors have been shown experimentally to trigger AβO buildup in sporadic AD, lifestyle and therapeutic interventions aimed at modifying these risk factors in humans have yet to show definitive success. This further highlights the difficulties and challenges associated with developing successful interventions for AD, even when the therapeutic target (AβOs) plays an established role in the disease process.
Endogenous protection and its failure
Astrocyte-mediated clearance of Aβ
A growing body of evidence indicates a role for astrocytes in clearing excess levels of Aβ from the brain [352]. Interestingly, it seems as if astrocytes have differing abilities to clear different aggregation states of Aβ, presumably due to size differences. Not surprisingly, astrocytes seem to have a harder time clearing fibrils compared to AβOs [353]. Astrocyte-mediated clearance of Aβ seems to occur by multiple mechanisms, as recently reviewed, including receptor-mediated uptake, secretion of degrading enzymes, and secretion of ApoE, which acts as a chaperone [352]. A few astrocyte receptors implicated in Aβ clearance are of note: RAGE, which is currently being targeted therapeutically in phase III clinical trials (http://clinicaltrials.gov), and matrix metalloproteinases, which are implicated in AβO-induced disruption of the blood-CSF barrier integrity (see discussion below). Overall, one possibility is that astrocyte protection of the brain from Aβ fails when Aβ accumulation reaches a certain threshold at which astrocyte-mediated clearance is saturated [352]. This hypothesis is consistent with advanced astrocyte pathology in AD brain that is detected by a monoclonal antibody developed against AβOs [89].
It may be the case that astrocytes near amyloid plaques switch from a neuro-supportive role to an inflammatory role. An opposing view is that astrocyte failure in AD is neuroprotective [354]. Another hypothesis is that Aβ, specifically AβOs, can stimulate astrocytes to produce and secrete more Aβ [355, 356]; this mechanism seems to occur through Ca++-sensing receptors expressed on the astrocytes [277]. Other evidence suggests that astrocytic metabolic dysfunction may regulate Aβ production through AβPP processing [354]. And astrocytes may also mediate Aβ clearance through induction of microglial phagocytosis [278]. Thus, more experimentation is needed to fully elucidate the role of astrocytes in Aβ production and clearance within AD. It also was shown recently that healthy astrocytes secrete insulin and IGF1 that act to protect neurons from AβO toxicity [73]. Note that these mechanisms need not be mutually exclusive.
Insulin
Extensive evidence indicates that insulin signaling and AβOs are connected in a vicious cycle of pathogenesis, as recently reviewed [200, 357]. This vicious cycle may be initiated, in some cases, by diabetes, which decreases insulin signaling in the brain. Since insulin signaling protects against AβO accumulation [346] and neurotoxicity [66], this leads to increased AβO accumulation and AβO-associated damage in the brain. AβOs themselves then further disrupt insulin signaling at many levels via pro-inflammatory mechanisms [357], e.g., by downregulating the expression of IRs on the plasma membrane [66]. Thus, a vicious pathogenic cycle is created in which AβOs upregulate their own production and aggregation by disrupting the physiological actions of insulin. Such a mechanism could account in part for AβO buildup in AD brains.
Importantly, the cellular stress and synaptic dysfunction induced by AβOs can be counteracted by stimulating brain insulin signaling [66, 322]. Therefore, either insulin or therapeutics aimed at increasing/repairing insulin signaling may be promising candidates for the treatment of AD [322]. One study exemplifying this promise demonstrated that the anti-diabetes agent exenatide protects against AβO-induced pathologies in cell culture and AβO-induced impaired insulin signaling and cognitive deficits in mice [358]. Furthermore, a recent study testing the effect in Tg mice of a therapeutic targeting multiple receptors involved in insulin signaling found a multitude of benefits including reversal of memory deficits, reduction of apoptotic factors, increase of factors promoting synaptic health, increase in neurogenesis, and reduction in Aβ, neuroinflammation, and oxidative stress [359]. Hopefully the multiple drugs targeting insulin signaling that are currently in clinical trials (see discussion below) also will have such a robust therapeutic effect.
Glymphatic system and impaired AβO clearance
The recently discovered glymphatic system functions to remove metabolic waste, including soluble proteins, from the CNS [360]. The glymphatic system involves CSF inflow to the brain, which drives interstitial fluid to clear waste out of the brain. Recent studies in mice show that glymphatic activity decreases sharply during aging, resulting in decreased Aβ clearance from the brain [320]. Studies in AD mouse models show that this decreased Aβ clearance is due to oligomer formation [321], especially HMW AβOs [166], indicating that the size of larger AβOs may make it more difficult for the glymphatic system to clear them from the brain. Poor sleep quality, which is associated with dementia, might negatively affect the activity of the glymphatic system [340]. CSF levels of Aβ have been found to be increased significantly in insomnia patients [361]. Thus, the role of the glymphatic system in AD with regards to Aβ may be that its decreased activity leads to impaired AβO clearance, which may lead to further aggregation resulting in larger AβOs or insoluble amyloid plaques, both of which the glymphatic system cannot clear. Ultimately, repairing glymphatic activity may be another option for therapeutic targeting in AD treatment.
Blood-CSF barrier
The function of the blood-CSF barrier is to keep undesirable molecules and pathogens out of the brain. Several studies have shown that the integrity of the blood-CSF barrier is disrupted in AD [362], and recently, evidence has been presented that this disruption can be induced by AβOs, seemingly through increased expression and activity of matrix metalloproteinases [363]. Based on their data, the authors of this study hypothesize that AβO-induced breakdown of the blood-CSF barrier might be an early event in AD pathogenesis that would contribute to the enhancement of neuroinflammation. Therefore, early therapeutic inhibition of matrix metalloproteinases may decrease neuroinflammation in AD.
Newer AD models
There is growing consensus in the literature that Tg mice are not ideal models of AD, partly because they are based on genetic mutations present in only <5% of AD patients. Furthermore, although many therapeutics have ameliorated cognitive deficits and/or neuropathology in Tg mouse models, these same therapeutics have failed in clinical trials. The traditional Tg mouse models that express mutant forms of human APP and/or presenilins generally do not develop tau pathology unless they also express mutant forms of tau [364–367]. However, tau mutations are associated with other tauopathies, not AD. It has been argued that Tg mouse models actually simulate the asymptomatic phase of AD and therefore are telling us how to prevent AD, not cure AD [368]. Next-generation Tg mouse models are being developed that accumulate Aβ without phenotypes related to Aβ overexpression, which may be unrelated to AD. It has been recommended that these models be used with the caveat that researchers consider the strengths and limitations of each model against the scientific and therapeutic goal of a prospective preclinical study [369]. There remains a call for more suitable models that recapitulate sporadic AD and more closely model human physiology.
Perhaps one of the most exciting AD model systems recently introduced is the NHP. NHPs have an APP sequence that is completely homologous to that of humans [370] and they develop plaques and tau pathology [370–372]. To speed up development of AD pathology, researchers introduced Aβ preparations containing fibrils into NHPs via i.c.v. injection. This resulted in microglial activation, neuronal loss, and tau phosphorylation [373, 374]. In a major advance for the AβO field, researchers from Brazil and Canada showed that i.c.v. injection of AβO preparations free of fibrils into NHPs induced fundamental features of AD pathology without development of Aβ fibrils and plaques [46]. The pathological features induced by AβOs in these NHPs included synapse loss, tau hyperphosphorylation, and activation of astrocytes and microglia. Most importantly, this research team recently reported that sustaining AβO injections for 12–18 months results in memory deficits and synapse loss [375]. This NHP model shows great promise as a superior AD model for therapeutic testing.
Another potentially powerful new AD model system comprises human induced pluripotent stem cells, or iPSCs. iPSCs derived from both familial AD and sporadic AD patients have been studied, and these AD-derived iPSCs show AβO accumulation, ER stress, oxidative stress, and tau hyperphosphorylation [376, 377]. These studies indicate that familial AD and sporadic AD iPSCs exhibit differential manifestations of ER stress [376] and Aβ40 accumulation [377], indicating that different therapeutics may be effective for patient subsets. In 2015, an organoid human iPSC-derived system was developed, also termed a “3D human neural cell culture system” [378]. This iPSC system developed key events in AD pathogenesis, including extracellular aggregation of Aβ and accumulation of pTau. The De Strooper group recently created a novel chimeric model wherein human iPSCs were studied in a more natural environment, i.e., via transplantation into the brains of APP Tg immunodeficient mice [379]. These human neurons were able to differentiate and integrate into the mouse brain, express 3R/4R tau splice forms, show abnormal phosphorylation and conformational tau changes, and undergo neurodegeneration. Remarkably, transplantation of these human iPSCs altered gene expression, upregulating genes involved in myelination and downregulating genes related to memory and cognition, synaptic transmission, and neuronal projection. Therefore, human iPSC models are attractive AD models for their human origin and ability to integrate into mouse models, which are more easily utilized than NHP models.
Efforts are also being made at developing improved rodent models for AD. Octodon degus, a small rodent endemic to Chile, needs no genetic manipulation as its Aβ sequence differs from human in only 1 amino acid (H13R). Unlike mouse and rat Aβ, this Aβ sequence does form AβOs and this rodent also develops plaques and pTau with age [380, 381]. Tg rats are also being developed as AD models. The TgF344-AD rat expresses mutant APPsw and PS1ΔE9 genes and manifests age-dependent cerebral amyloidosis that precedes tauopathy, gliosis, apoptotic loss of neurons in the cerebral cortex and hippocampus, and cognitive disturbance [367]. These rats also exhibit pathological changes in the retina, including plaques and inflammation, e.g., microglial recruitment and complement activation [382]. This is interesting considering AβOs may also be involved in the pathogenesis of macular degeneration and glaucoma (see discussion below). Another Tg rat was developed using the Tg2576 mouse protocol. These rats exhibit cognitive deficits at 8–12 months, activated astrocytes in the brain, ThioS staining in the hippocampus and cortex, and elevated levels of Aβ38, Aβ40, and Aβ42 [383]. Tg rats expressing the Swedish and Indiana APP mutations also exhibit elevated levels of these Aβ peptides in the CSF [384] as well as pre-plaque intracellular AβO-associated cognitive impairment [87]. There are indications that non-Tg rabbit, which expresses the human Aβ sequence, may also prove valuable for studies of AβO etiology [346].
The use of Drosophila as an AD model was reviewed recently [385]. Drosophila has homologues of human APP and tau. Other advantages of Drosophila for use as an AD model include low genome redundancy, which greatly simplifies the analysis of single gene disruption, short lifespan, and their low cost compared to mammalian models. Drosophila have been used to uncover or validate several pathological pathways or susceptibility genes and have been readily implemented in drug screening pipelines. Interestingly, using a Tg Drosophila model expressing the Artic mutation, it was found that AD-like pathologies affected the circadian system in an age-dependent manner [386]. These Tg flies showed a dramatic degradation of circadian rhythms in tune with their reduced longevity and impaired climbing activity.
The use of Caenorhabditis elegans as an AD model was recently reviewed [387]. C. elegans is useful as an AD model as it has homologs of AD-related genes, including APP, tau, and PSEN1. C. elegans has complex biochemical pathways just like mammals, many of which are conserved. Its neuronal connectivity has already been established, making it a suitable model for learning and memory impairments [387]. Furthermore, C. elegans has a short lifespan, thereby speeding up study of Aβ accumulation, a process that can take months or years in other model organisms. To directly study the impact of the exact human Aβ42 sequence, Tg worms also have been developed and these model organisms have been shown to accumulate AβOs and utilized to study AβO-directed therapeutics [388–391].
Therapeutics
Eliezer Masliah, the current head of the NIA’s Division of Neuroscience, proposed multiple possible strategies for targeting the AβO pathogenic cascade in a 2014 commentary in PNAS (Fig.9) [392]. He and co-author Cassia Overk proposed that therapeutics for AD might involve 1) directly clearing AβOs; 2) blocking AβO surface receptors; 3) interfering with AβO-induced signaling pathways; or 4) decreasing downstream effectors such as tau. The AD therapeutics currently in clinical trials are described in a systematic review of clinicaltrials.gov conducted in January 2017 by Cummings and colleagues [393]. Approximately half of the 105 agents currently in clinical trials are amyloid related. Figure10 illustrates the mechanistic targets of many of the amyloid-targeted therapeutics. Some of the current efforts germane to AβO pathogenic mechanisms are discussed below.
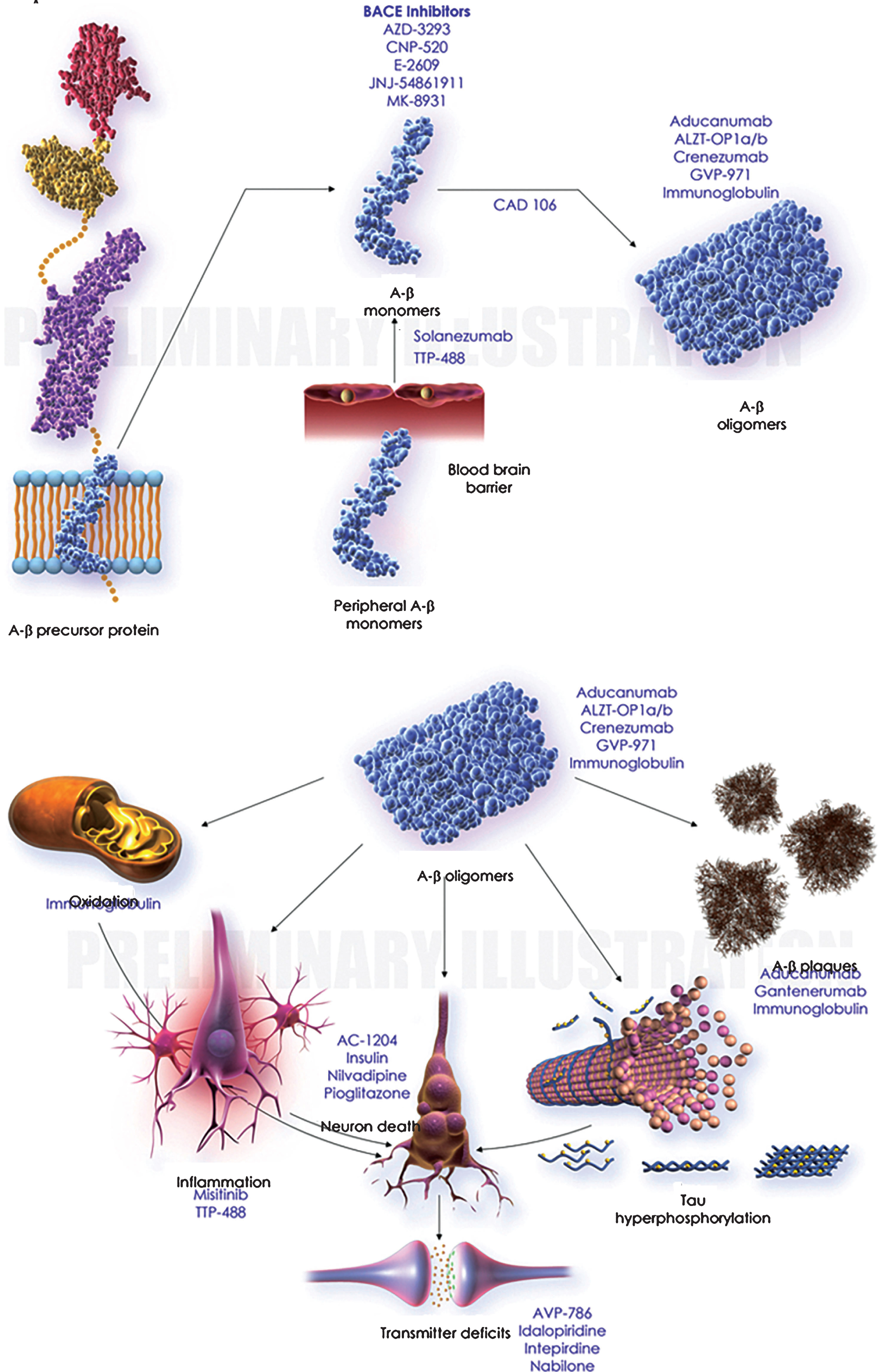
Anti-Aβ immunotherapies represent 8% of the phase II pipeline and 18% of the phase III pipeline. In 2014, Goure and colleagues of Acumen Pharmaceuticals proposed in their review of immunotherapeutics that current Aβ-directed therapies were failing due to lack of selectivity for AβOs; instead, they bind to monomeric or fibrillar Aβ, or both [101]. Monomers and fibrils are more abundant than AβOs in the AD brain, but less germane to nerve cell damage. The authors suggest that the affinity for monomers and/or fibrils by Aβ immunotherapies in clinical trials is why high doses are required for therapeutic benefit. Acumen, in partnership with Merck, has developed an antibody, known as ACU193, that is unique among Aβ immunotherapeutics in its selectivity for Aβ oligomers. ACU193 has greater than 500-fold selectivity for AβOs over fibrils [394] and greater than 2500-fold selectivity for AβOs over monomers [34]. Success of ACU193 in clinical trials would provide compelling evidence for the hypothesis that soluble AβOs are the primary toxins instigating AD pathogenesis.
What may be two examples supporting AβO-directed immunotherapies are Crenezumab and Aducanumab. Genentech reported at the 2017 Clinical Trials on Alzheimer’s Disease (CTAD) conference that their Aβ immunotherapy Crenezumab had a 10-fold higher affinity for AβOs over Aβ monomers [395]. In immunoprecipitation experiments, its main target was AβOs that are, or dissociate into, SDS-stable dimers. It is of note that dimers themselves are not thought to be toxic [129]. Genentech reported that primary efficacy endpoints were not met for Crenezumab in two phase II trials (ABBY and BLAZE), although subgroup analysis showed greater reduction in cognitive decline in patients with mild AD given the higher dose of Crenezumab. They are currently enrolling patients in two phase III studies (CREAD and CREAD2), which will dose up to 4-fold higher.
Aducanumab (Biogen), an antibody that targets Aβ oligomers and fibrils, has shown in phase Ib trials a reduction of amyloid plaques in a dose- and time-dependent manner and, most importantly, a slowing of cognitive decline. Slowing of cognitive decline required the highest doses tested. However, these doses caused amyloid related imaging abnormalities [105]. While there is optimism that higher doses of these antibodies will result in greater therapeutic efficacy, a fully AβO-selective antibody may be essential for using low doses to avoid complications while still providing disease-modifying efficacy.
BACE inhibitors represent 6% of the phase II pipeline and 18% of the phase III pipeline. Supporting evidence for the therapeutic value of BACE inhibitors comes from the protective A673T APP Icelandic mutation in humans, which reduces BACE processing of AβPP [396, 397]. On the other hand, genetic deletion of BACE in mice causes many side effects, most notably the AD symptoms of neurodegeneration and memory dysfunction [398]. Since the systematic review conducted by Cummings and colleagues in January 2017 [393], Merck has halted its phase II and III trials of the BACE inhibitor verubescestat for lack of efficacy; a leading theory for this failure is the timing of treatment [399]. A recent success in AD mouse models, demonstrating beneficial effects on cellular, long-range circuitry, and memory impairment, has motivated researchers to start another clinical trial with a modified BACE inhibitor [400].
The relationship between insulin resistance and AβOs in AD is discussed above. Therapeutically, insulin signaling may be a convenient target if the many treatments already developed for type 2 diabetes could be repurposed for AD [406, 407]. In hippocampal cell cultures, exogenous as well as astrocyte-secreted insulin and IGF1 displace AβOs bound to cell surfaces [73]. Diabetes drugs fall into five categories: intranasal insulin, incretins, dipeptidyl peptidase 4 (DPP-4) inhibitors, peroxisome proliferator activated receptor (PPAR-γ) agonists, and the common diabetes treatment metformin [406]. Intranasally delivered insulin has been shown to improve memory function in AD patients, although different studies have obtained inconsistent results as to whether this is effective in APOE E4-positive AD patients [406]. The impact of intranasally delivered insulin on AD is currently being investigated in phase I and II clinical trials sponsored by Wake Forest University. Incretins are gastrointestinal hormones that stimulate insulin secretion and inhibit glucagon release in a glucose-dependent manner [406]. Liraglutide, an incretin analog, has been shown to reverse memory impairment, synaptic loss, and reduce plaque load in aged APP/PS1 mice [408]. Liraglutide is currently in phase II clinical trials (Imperial College London). DPP-4 inhibitors block degradation of incretins and have been found to improve memory function, reduce levels of Aβ and pTau, and decrease inflammation in AD rodent models [406]. However, no DPP-4 inhibitors are currently in clinical trials [406]. Activation of PPAR-γ induces the expression of multiple genes involved in the insulin signaling cascade, which improves insulin sensitivity in patients with type 2 diabetes [406]. Metformin, a very widely prescribed drug for diabetes [409], is currently in clinical trials to examine its effect both on aging in general and AD. On the other hand, it has been recently linked to an increased dementia risk in diabetes patients [410].
Combination treatments
Given the complex neuropathology of AD and the lack of effective biomarkers for sporadic AD, it may be the case that multi-factorial, combination treatments will provide the greatest efficacy in AD treatment. This is a sentiment shared by many in the field [5, 418]. In October 2017, Amylyx Pharmaceuticals received a grant from the Alzheimer’s Association and the Alzheimer’s Drug Discovery Foundation to conduct phase II clinical trials with the combination therapy AMX0035 (Alzheimer’s Association). AMX0035 is a combination of two compounds that synergistically block mitochondrial and ER stress. Preclinical studies show that this combination protects brain cells from inflammation and oxidation in models of amyotrophic lateral sclerosis, AD, and mitochondrial diseases [419]. ALZT-OP1 (AZTherapies) is a combination of two small molecule drugs, cromolyn, an asthma drug, and ibuprofen. Cromolyn was found to inhibit Aβ aggregation in vitro and reduce soluble Aβ in the brain in vivo [420]. The intended effect of ibuprofen in the combination therapy is to reduce neuroinflammation. ALZT-OP1 is currently in phase III clinical trials. It is likely that many more combination therapies will arise in the future.
AD prevention: Diet, exercise, and mental/social engagement
One review article states that although it is difficult to make conclusions regarding diet as an AD risk-factor due to the difficulty in analyzing eating patterns, there does seem to be clear evidence that diet influences AD. Protective foods identified include fish, fruit, coffee, and wine. There is also evidence that a diet high in saturated fats may increase AD risk [421]. In a Tg mouse AD model, high cholesterol promotes earlier buildup of AβOs [350]. A systematic review and meta-analysis by researchers at the Mayo Clinic found that a higher adherence to the Mediterranean diet is associated with a reduced risk of developing mild cognitive impairment (MCI) and AD and a reduced risk of progressing from MCI to AD [422]. Vitamin B may also have a positive effect [423]. To more directly test the impact of diet on AD risk, participants are currently being recruited for a clinical trial to determine the effect of saturated fat and glycemic index on cognition in older individuals with or without an APOE E4 genotype (sponsored by the University of Washington). APOE4 is known to affect the presence and impact of AβOs [424–430]. Recruitment is underway also for clinical trials testing Genistein, a dietary supplement that has been found to have antioxidant and neuroprotective effects on AD and increases PPAR-γ levels, which results in increased APOE expression and Aβ degradation (Fundación para la Investigación del Hospital Clínico de Valencia). The omega-3 fatty acid DHA, which protects against AβO-instigated dendritic spine loss [431], shows potential to decrease AD incidence in APOE4 carriers [423, 432] and is under clinical investigation (http://clinicaltrails.gov). In addition to the potential for diet to modify AD risk, it is well recognized that physical activity also modifies risk [433]. In AD mouse models, exercise decreases AβO levels and increases cognitive performance [434–437]. Furthermore, evidence has been found for mental/social engagement modifying AD risk and AβO accumulation in a mouse model [438] and in humans [439–441]. Therefore, it is encouraging that some cases of sporadic AD may be delayed or even prevented by modifiable lifestyle factors.
AβOs as biomarkers: can AβOs provide metrics to assess experimental drug efficacy and ultimately give a diagnostic to indicate a patient should start AD treatments?
Newly emerging approaches have begun to focus on therapeutic targeting of AβOs, and not amyloid plaques, as AβOs are the form of Aβ that instigates the neural damage leading to AD. A powerful metric for the efficacy of these new approaches to disease-modifying therapeutics would be to monitor a patient’s AβO levels.
There are two big challenges to using AβO levels as a biomarker. First, there is a need for extraordinary sensitivity. Second, there is a need to discriminate oligomeric Aβ from the other, much more abundant but chemically similar forms of the peptide. A uniquely sensitive and specific AβO immunoassay, capable of attomolar quantitation, initially showed that median AβO levels in AD CSF were 10-30-fold higher than in non-AD controls [32]. Although powerful, this assay was not practical, and it has not been explored for clinical use. Since then, other groups have utilized various immunoassay platforms and Aβ-targeting antibodies with varying results. A team at Merck using the AβO-specific antibody ACU193 (see Therapeutics section above), found a significant 3-5-fold increase in AβOs in AD CSF compared with aged-matched controls [34]. On the contrary, Jongbloed and colleagues found that CSF AβO levels decreased from non-dementia to MCI to AD [165]. Another study found no significant difference between AβO levels in AD CSF and controls. However, this study found a small, but significant increase in AβOs levels in MCI compared to controls, suggesting an early rise in AβO levels followed by a later decrease [35]. Most recently, an AβO-specific plasma assay was able to differentiate AD from controls with 78.3% sensitivity and 86.5% specificity, finding AβOs elevated in AD plasma [442]. CSF/plasma measurements of whole-AβO populations do not seem to be diagnostically useful at present due to inconsistent results, but the possibility remains that an assay targeting specific populations of AβOs may be more useful [33, 443–445]. Indeed, a recent study showed that an assay targeting HMW AβOs could be used to monitor efficacy of an anti-amyloid therapeutic [446]. This finding is consistent with immunoassays showing therapeutic efficacy in a mouse model correlated with reduction in a pool of putative type 1 AβOs specifically recognized by the NU4 monoclonal antibody [120].
An alternative to measurements of AβOs in CSF or blood comes from new technologies for AβO imaging. The Pronucleontrademark platform from Adlyfe consists of series of engineered peptides that provide a unique readout when associated with beta-rich fibers and AβOs. There are indications that it can be used for pre-plaque imaging [447]. Another technology is the development of magnetic resonance imaging (MRI) [96] and positron emission tomography (PET) [199] probes using modified antibodies that have high selectivity and affinity for AβOs. These have been shown to discriminate AD Tg mice from non-Tg littermates and to discriminate human AD brain slices from non-AD specimens. The availability of humanized AβO-specific antibodies makes it likely that these probes will soon be ready for clinical testing [101].
In the future, it is foreseeable that scheduled clinical tests of AβO levels could provide an indicator of whether a patient should begin an appropriate AD treatment. Monitoring levels after diagnosis and treatment would provide an initial indicator of how well a patient is responding to a therapeutic. Use of AβO levels as an AD biomarker would thus be akin to monitoring glycated hemoglobin by A1C levels for diabetics.
Other dysfunctions/degenerative neural disorders linked to AβOs?
As discussed above, TBI and diabetes may be etiological factors in the buildup of AβOs. In addition, it may be the case that once accumulated, AβOs may contribute to further pathogenic consequences in these diseases such as insulin resistance (see discussion above), dementia, or chronic traumatic encephalopathy (CTE). AβOs, specifically, have not yet been implicated in CTE, but CTE is associated with repetitive brain injury and amyloid plaques have been observed in CTE brain tissue [448]. Additional disorders that have been linked to AβOs are inclusion body myositis, glaucoma, and macular degeneration. Inclusion body myositis is the most common progressive muscle disease of patients above the age of 50. Forms of Aβ, including AβOs, are known to accumulate in the muscle fibers of diseased patients [449]. Further evidence implicates AβOs in glaucoma and macular degeneration. AβOs have been found to contribute to apoptosis of retinal ganglion cells in glaucoma [450]. Intravitreal injection of AβOs in rat induces molecular changes associated with apoptosis in the rat retina. Apoptosis is hypothesized to take place in macular degeneration according to bioinformatics analysis [451]. AβOs, through RAGE, upregulate VEGF, which stimulates neovascular age-related macular degeneration [452]. Structures reactive with the OC antibody have been found in drusen, a hallmark of eyes affected by macular degeneration [453]. The OC antibody is well characterized and binds to fibrillar type 2 AβOs [148]. In fact, a modulator of Aβ aggregation (MRZ-99030) is neuroprotective with therapeutic treatment in animal models of glaucoma and macular degeneration [454]. However, much more evidence is needed to understand the role of AβOs in these neural disorders.
CONCLUDING REMARKS
As evidenced by the increasing number of publications concerning AβOs in the past 5 years, and the consistency in data supporting a toxic role for AβOs, the AβO hypothesis for AD pathogenesis has garnered considerable support and acceptance. Accordingly, the number of AβO-targeting therapeutics in the AD pipeline has begun to increase. We believe that this emerging interest in AβO targeting will prove beneficial to the treatment and diagnosis of AD. These efforts potentially can extend to a broader proportion of the population, given the evidence for a role for AβOs in other diseases in addition to AD. Ultimately, for these efforts to result in therapeutic and diagnostic benefits, further advances in AβO structure-function studies are needed. Continued investment into this and other research involving AβOs will enable the closing of critical gaps, thereby paving a smoother and shorter path from bench to bedside.