Oral squamous cell carcinoma (OSCC) represents 95% of all cancers of the head and neck region. The five-year survival rate of OSCC patients is about 60% and has not gone throw significant improvements despite recent advances in molecular biology, or the identification of key pathways in its pathophysiology such as cell cycle.
OBJECTIVE:
1) to analyse the inmunoexpression of cell cycle checkpoints (CPs) in an OSCC institutional cohort and to relate it to clinicopathological features and survival, and 2) to study CPs-related genes in the OSCC subset of the TCGA database.
METHODS:
Immunohistochemistry (IHC) for p16, p21, cyclin D1 and p27 protein expression was quantified by tissue microarray analysis in 68 samples from OSCC patients. In order to analyse its correlation with genetic information, a cohort belonging to The Cancer Genome Atlas (TCGA) database was analysed.
RESULTS:
Of 68 patients, 34 (50%) developed recurrence, and 36 (52.09%) died as a result of disease progression (mean survival 34.09 months). IHC staining for nuclear cyclin D1 was associated with worse staging and tobacco use. p16, p21, cyclin D1, and p27 expression was unrelated to overall survival. No statistically significant correlation linked the CPs-related genes mutations to OSCC overall survival in the TCGA database.
CONCLUSIONS:
CPs variations at a phenotype and genotype level seem not to affect significantly clinicopathological features and survival in the studied OSCC cohorts.
Carcinogenesis is an intricate multistep process initiated by the accumulation of genetic variations in proto-oncogenes and tumour suppressor genes (TSGs) [1]. Cells have evolved strategies to prevent those hazardous genetic events. Nonetheless, those machineries are not always able to repair genomic instability triggering cell-cycle defects, angiogenic switch or senescence which ultimately results in a cancer cell generation and preservation [2]. The cell cycle or division is the process in which cell duplicates its contents and divides, resulting in two genetically identical daughter cells. Cell division is monitored by cell cycle checkpoints (CPs). The canonical CPs are cyclins and cyclin-dependent kinases (CDK), these complexes are inhibited by CDK inhibitors (CKIs). These proteins are classified as TSGs due to their ability to disrupt cell proliferation, and according to a bulk of molecular oncology research their coding genes are located in altered chromosomal areas across multiple human cancers [3]. Based on structural and functional characteristics, there are two CKIs families: the CIP/KIP family (including p21, p27, and p57), and the INK4 family (including p15, p16, p18, and p19) [4].
CIP/KIP proteins have a broader spectrum of substrates, binding to cyclin D, cyclin E and CDKA-1 complexes, so having greater influence in carcinogenesis. P21 is activated under stress conditions and can follow two pathways: one dependent on p53, and another independent of p53 stimulated by cell growth factors [5]. The p27 was the first CDK inhibitor discovered and its related gene is located on chromosome 12p-12-12p13.1. Both aforementioned proteins are responsible for blocking the activity of the cyclin E/CDK2 and cyclin D/CDK2 complexes, inhibiting phosphorylation of pRb and preventing the cellular transition from phase G1 to phase S, which triggers apoptosis. P16 is another relevant TSG belonging to the INK4 family whose coding gene locus is located at MTS-1 on chromosome 9p. During the G1 phase, p16 controls cell cycle by inhibiting the ability of the cyclin D/CDK4/6 complexes to phosphorylate pRb. The phosphorylation of pRb leads to the dissociation of the pRB/E2F complex, which leads to cell cycle progression. On the other hand, the release of E2F activates the transcription of p16. The increase of p16 levels, results in CDK4/6 blocking, which ultimately triggers cyclin D/CDK4/6 complex inhibition. CPs variations have been classically related to human tumorigenesis due to its amplification or overexpression can lead to some hallmarks of cancer [5, 6, 7].
Oral squamous cell carcinoma (OSCC) (ICD 01-10) 1 is the 8 most common cancer among men and the 14 most common among women in the US. The overall five-year survival rate of OSCC affected patients has been for decades about 50%, despite advances in therapeutic and aids to prevent diagnostic delay. This standing poor prognosis is associated to the existence of suboptimal anticancer strategies characterised by severe treatment-associated sequels and acquired multidrug resistance leading to recurrences and second primary tumours [8].
Our group has previously unravelled the implications of several CPs in OSCC by means of comprehensive reviews [9, 10, 11], and immunohistochemistry reports [12, 13]. Nonetheless, in current literature there is a lack of studies evaluating multiple CPs altogether constructing a panel in a single cohort of OSCC-affected individuals. Moreover, literature lacks bioinformatics analysis able to integrate available cross-platform data on the implications of CPs in OSCC despite the existence of genomic and molecular profiling datasets like The Cancer Genome Atlas (TCGA) [14]. Prompted by discussed literature, the research hypothesis is that this CPs panel may provide a better characterization of tumour biology and a stronger prognostic value on survival than the use of single markers.
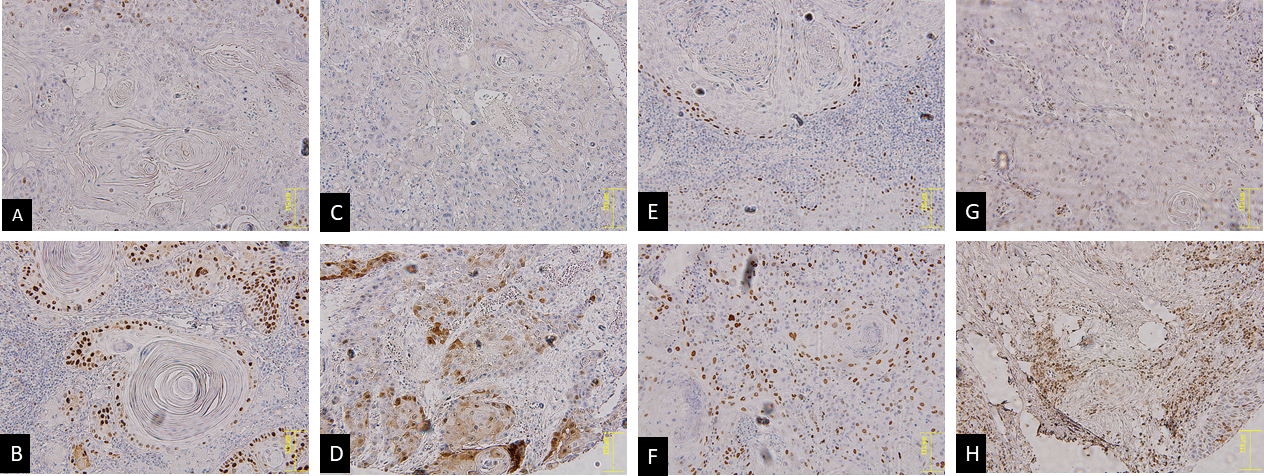
Immunohistochemistry staining examples in cyclin D1 negatively (A) and positively (B) stained, p16 negatively (C) and positively (D) stained, p21 negatively (E) and positively (F) stained, p27 negatively (G) and positively (H).
The objective of this study was twofold: 1) to assess the predictive impact of a immunohistochemical markers panels, specifically of CPs, in an OSCC well defined cohort and with minimum five-year follow-up relating it to clinical and pathological features and 2) to integrate this information with TCGA genomic database.
Association between p16, p21, cyclin D1 and p27 staining expression with the clinicopathological parameters
Clinicopathological
Variables
p16INK4a
p21Cip1
p27Kip1
Cyclin D1
information
Total (%)
Positive (%)
value
Positive (%)
value
Positive (%)
value
Positive (%)
value
Age
55 years
14 (25.59)
13
5 (38.46)
0.728
12
5 (41.67)
0.356
9
8 (88.89)
0.310
9
3 (33.34)
0.803
55 years
54 (79.41)
55
17 (30.91)
50
14 (28%)
29
21 (72.41)
29
11 (37.93)
Gender
Male
33 (48.53)
31
13 (41.94)
0.217
30
9 (30)
0.915
15
12 (80)
0.666
24
6 (25)
0.557
Female
35 (51.47)
33
9 (27.27)
32
10 (31.25)
23
17 (52.17)
14
8 (57.14)
Tumour location
Tongue
6 (8.82)
5
1 (20)
0.803
5
4 (0.8)
0.204
4
3 (0.75)
0.098
3
2 (1.5)
0.652
Gingiva
17 (25)
17
7 (41.18)
16
4 (25)
6
5 (83.33)
7
4 (57.14)
Retromolar trigone
6 (8.82)
6
2 (33.34)
6
1 (16.67)
4
4 (100)
4
1 (25)
Lip
22 (32.35)
22
8 (36.36)
19
5 (26.32)
14
10 (71.43)
14
4 (28.57)
Floor of the mouth
8 (11.76)
7
1 (14.29)
7
3 (42.86)
4
1 (25%)
4
1 (25)
Palate
8 (11.76)
7
3 (42.86)
8
2 (25)
6
6 (100)
6
2 (33.34)
Differentiation
Well
30 (44.12)
29
9 (31.03)
0.090
27
9 (52.94)
0.922
15
11 (73.33)
0.408
15
6 (40)
0.913
Moderate
30 (44.12)
28
8 (28.57)
28
8 (28.57)
18
13 (72.22)
18
6 (33.33)
Poor
8 (11.76)
7
5 (71.43)
7
2 (28.57)
5
5 (100)
5
2 (40)
T stage
I
20 (29.41)
18
8 (44.44)
0.473
17
4 (23.53)
0.884
12
11 (91.67)
0.176
12
3 (0.25)
0.095
II
12 (17.65)
12
4 (33.33)
12
4 (33.33)
8
4 (50)
8
1 (0.125)
III
9 (13.24)
8
1 (12.5)
7
2 (28.57)
1
1 (100)
1
1 (100)
IV
27 (39.71)
26
9 (34.62)
26
9 (34.62)
17
13 (76.47)
17
9 (52.94)
N stage
0
55 (80.88)
53
19 (35.85)
0.774
50
18 (36)
0.144
36
27 (75)
0.418
36
13 (36.11)
0.692
1
7 (10.29)
6
2 (33.34)
6
1 (16.67)
2
2 (100)
2
1 (50)
2b
6 (8.82)
5
1 (20)
6
0
0
0 (0)
0
0 (0)
Stage
I II
32 (47.06)
30
12 (43.33)
0.373
29
8 (42.11)
0.624
20
15 (75)
0.841
20
4 (20)
0.023*
III IV
36 (52.94)
34
10 (29.41)
33
11 (33.33)
18
14 (77.78)
18
10 (55.56)
Tobacco use
Non-smoker (never)
32 (47.06)
30
13 (43.33)
0.286
27
8 (29.63)
0.902
15
12 (80)
0.907
14
6 (42.86)
0.027*
Former smoker ( 10 years)
12 (17.65)
11
2 (18.18)
11
4 (36.36)
8
6 (75)
9
6 (66.67)
Current smoker ( 5 years)
24 (35.29)
23
7 (30.43)
24
7 (29.17)
15
11 (73.33)
15
2 (13.33)
Alcohol use
Non drinker
33 (48.53)
30
13 (43.33)
0.347
28
8 (28.57)
0.620
16
13 (81.25)
0.621
15
7 (46.67)
0.515
Non-current drinker
22 (32.35)
21
6 (28.57)
21
8 (38.09)
17
13 (76.47)
18
6 (33.33)
Current drinker
13 (19.12)
13
3 (23.08)
13
3 (23.08)
5
3 (0.6)
5
1 (20)
Recurrence
No
34 (50)
31
14 (45.16)
0.078
30
7 (23.33)
0.227
18
15 (83.33)
0.454
18
5 (27.78)
0.272
Yes
34 (50)
33
8 (24.24)
32
12 (37.5)
20
14 (70)
20
9 (45)
Adjacent dysplasia
No
38 (55.88)
35
13 (37.14)
0.255
34
12 (35.29)
0.662
20
15 (75)
0.150
20
7 (35)
0.297
Mild
8 (11.76)
8
1 (12.5)
7
3 (42.86)
4
2 (50)
4
3 (75)
Moderate
4 (5.88)
4
3 (75)
4
1 (25)
3
3 (100)
3
0 (0)
Severe
1 (1.47)
1
0 (0)
1
0 (0)
1
0 (0)
1
0 (0)
In situ
17 (25)
16
5
16
3 (18.75)
10
9 (90)
10
4 (0.4)
*Stands for statistically significant.
Material and methods
Patients
The present retrospective cross-sectional study involved a cohort of 68 patients diagnosed with OSCC surgically treated at the University Hospital of Santiago de Compostela, Galicia, Spain, between April 2005 and March 2010. The study was performed in accordance with the Declaration of Helsinki of the World Medical Association, approved by the Ethics Committee of the Servizo Galego de Saúde (ID: 2013/541) as well as following the STROBE (Strengthening the Reporting of Observational Studies in Epidemiology) guidelines [15]. For patients included in this study, the staging was stablished according to the 7th American Joint Committee on Cancer (AJCC). The mean follow-up period was 48.55 47.29 months with a minimum of 2 months and a maximum of 211.81 months. Some of clinical and demographic data were published in our previous report [12].
Tissue microarray (TMA) generation
Paraffin blocks were selected on the basis of the availability of suitable formalin-fixed, paraffin-em- bedded tissue (at least 1-mm thick). For characterization of immunohistochemical protein expression, we constructed three different TMAs containing representative areas of every tumour. After microscopic evaluation, two areas of each tumour were selected, avoiding necrosis and keratin pools. Each tissue array was assembled as previously described. The areas chosen for the cylinder core were representative of the tumours. For each inmunolabelling run, a positive (tonsil) and negative (omission of primary antibody) controls were used to ensure quality of staining.
Immunohistochemistry of p16, p21, cyclin D1 and p27
Three-micrometer tissue sections from the TMA blocks were sectioned and applied to special immuno- histochemistry-coated slides (DAKO, Glostrup, Denmark). For the immunohistochemical study the following antigen retrieval, and clones were used according to the instructions of manufacturer: p16 (Dako, FLEX, RTU; clone E6H4), p21 (Dako, FLEX, RTU, Clone SX118), cyclin D1 (Dako, FLEX, RTU clone EP12), and p27 (Dako, FLEX, RTU, Clone SX53G8). Epitope retrieval was performed in 10 mM sodium citrate buffer (pH 6.0) using a water bath for 40 min at 95–99C. Endogenous peroxidase was blocked using peroxidase-blocking reagent (DAKO) for 5 min. Incubation with the primary antibody was performed at room temperature for 20 min, and staining was revealed with EnVision DAB (Dako). Isotype controls were used as negative controls in place of primary antibodies.
Evaluation of immunohistochemical results
Two of the authors (JSP and CCP) performed an observational quantification analysis in a joint session. For quantification, slides were digitized using an automate slides scanner to produce high-resolution images for visual quantitative analysis using an automatic image analysis system ACISRIII (Dako, Glostrup, Denmark). The digital images of the slides were captured by ACIS scanner at low power magnification and the entire slide images were viewed on a monitor. The region of highest immune percentage of positive cells were selected for further automatic scoring. A minimum of three of these areas containing only tumoral cells was selected manually for quantitative evaluation. The final score was the average result of the different areas measured. To confirm the accurateness of the measurements, selected areas from ten of the specimens were measured three times.
Staining evaluation was restricted to the nuclei for p21Cip1 and cyclin D1. Nonetheless, any degree of cytoplasmic or nuclear staining was considered for p27Kip1 and p16INK4a evaluation (Fig. 1). The expression of markers was semiquantitatively evaluated on the basis of the following thresholds as cut-off points for positivity or not: 5% (p21 and p16), and 50% (p27, and cyclin D1). These cut-offs were determined for each marker based on the mean of its expression score.
Bioinformatics analyses
All the raw data available from TCGA was processed by a public bioinformatics tool (www.cbiopor tal.org). By this platform, only OSCC-affected patients were extracted from the Head and Neck Cancer TCGA database (i.e., 172 patients/samples) in order to study genetic variations related to the previous evaluated CPs. Retrieved information was used to perform statistical analyses of correlation with survival [14].
Statistical analysis
Statistical analyses were performed by using SPSS statistical software 22.0 for Windows. In order to evaluate the correlation of marker expression with clinic-pathological variables, Spearman correlation test was used. Gaussian distribution of variables was checked through Kolmogorov-Smirnov normality test. The non-parametric test of was used in order to investigate the difference in expression between groups. Univariate Kaplan-Meier method was used to estimate the overall survival rates by means of the log-rank test. All the reported p values were 2-sided, and considered significant under 5%.
Association of p16, p21, cyclin D1 and p27 staining in the present oral squamous cell carcinoma (OSCC) cohort
p16 positive (%)
value
p21 positive (%)
value
Cyclin D1
0.361
0.010*
Negative
23
8 (34.78)
24
4 (16.67)
Positive
14
7 (50)
14
8 (57.14)
p27
0.039*
0.289
Negative
9
1 (11.11)
9
4 (44.44)
Positive
28
14 (50)
29
8 (27.59)
*Stand for statistically significant.
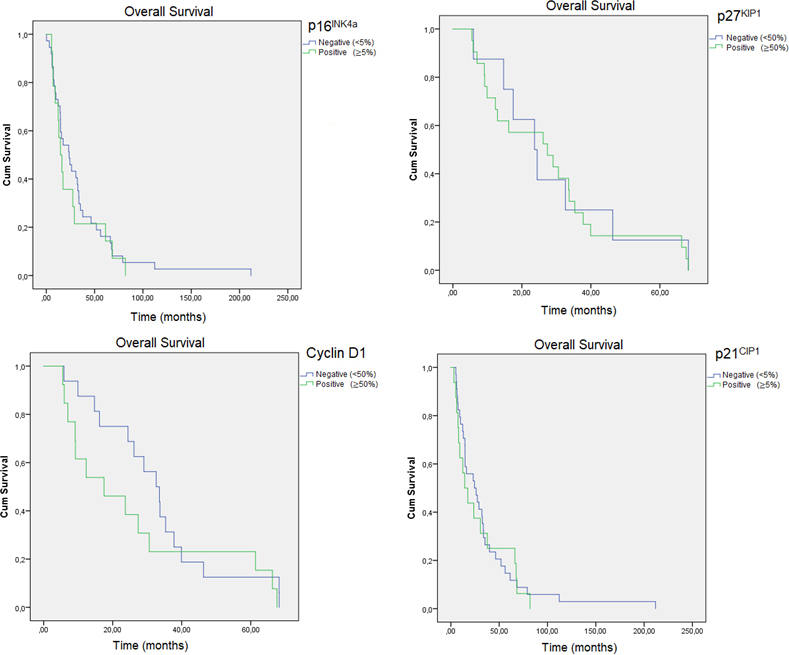
Kaplan-Meier curves of OS for the staining of p16, p21, cyclin D1 and p27 in the present OSCC cohort.
Results
Analysis of p16, p21, cyclin D1 and p27 expression in TMAs of OSCC mucosa samples
Clinicopathological characteristics and its association with semiquantitative protein expression is displayed in Table 1. Briefly, the majority of the patients were older than 55 and the distribution regarding gender was almost equal (48.53 vs. 51.47). In terms of classification, tumours have been grouped according to initial stage tumours (i.e., I–II) with 32 cases (47.8%) and advanced stage tumours (i.e., III–IV), with 36 cases (52.9%), most of the tumours did not show an adjacent dysplasia (55.88), nor regional lymph nodes metastasis (80.88%). None of the tumours presented distant metastasis. In stages I–II, 4 (20%) of tumours were positive for Cyclin D1, whereas in advanced stages 10 were (i.e., 55.56%) ( 0.023). The overexpression of this protein was also a fact among former and current smokers vs. never smokers as checked by the bonferroni correction ( 0.023).
Mean overall survival (OS) time by means of Kaplan–Meier method in each selected prognostic factors
Variable
Univariate analysis overall survival months SD
value*
HR
Age
55 years
35.942 10.299
0.751
0.101
55 years
33.630 5.736
Gender
Male
26.016 5.845
0.104
2.651
Female
41.880 7.845
Tumour stage
I II
48.971 8.905
0.006*
7.445
III IV
22.573 4.726
p16
Negative
34.021 6.418
0.593
0.286
Positive
26.392 6.744
p21
Negative
34.738 6.780
0.646
0.211
Positive
28.681 6.778
p27
Negative
29.189 7.035
0.632
0.229
Positive
27.798 4.392
Cyclin D1
Negative
32.633 4.452
0.233
1.424
Positive
26.431 6.525
* Log-rank test; HR, hazard ratio SD, standard deviation; CI, confidence interval. *Stands for statistically significant.
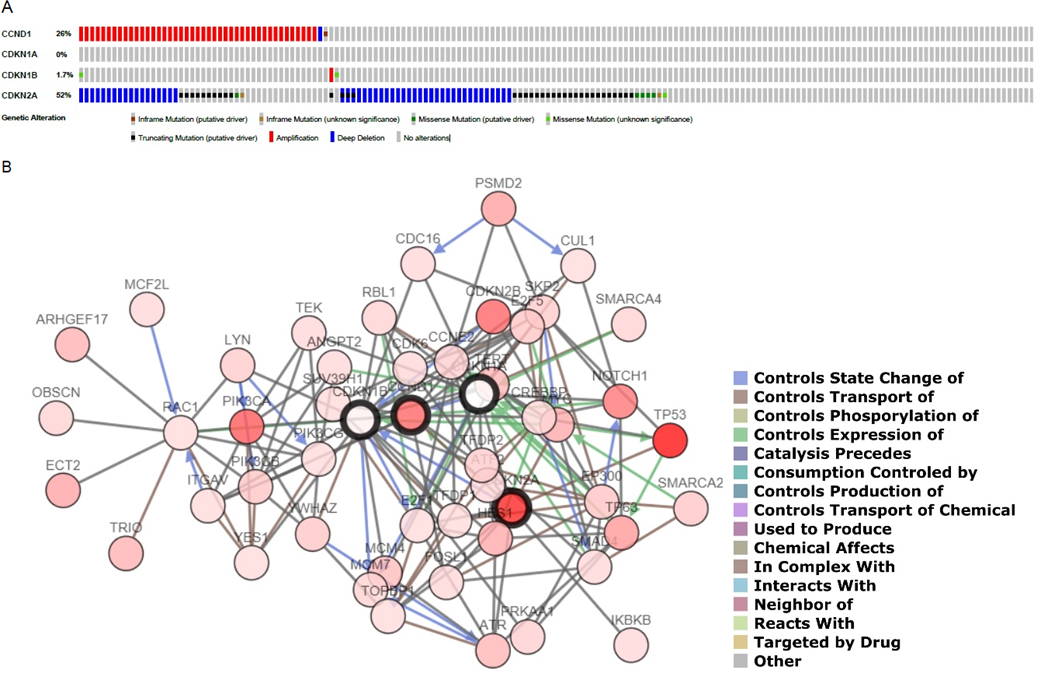
(A) Profiled mutation for CDKN2A, CDKN1A, CCND1 and CDKN1B in each OSCC-affected patient included in TCGA. (B) Network involved in the expression of the CPs genes. Dark circles represent selected genes and an interaction legend is available on the right corner.
The most frequent coexpressions detected in the present cohort are represented in Table 2. When Spearman’s test between markers was applied two strong correlations were detected, once between p27-p16 ( 0.340; 0.04) and the other between p21-Cyclin D1 ( 0.420; 0.009).
Mean overall survival (OS) time was 34.09 37.01 months. Tumours in advanced stages showed a lower survival (22.57 months; 95% CI 13.31–31.84) than those in early stages (48.97 months; 95% CI 31.52–66.42) (HR 7.45, 0.006). Intensity of staining for any IHC marker was not significantly associated with lower OS according to Kaplan-Meier analysis (Fig. 2).
Analysis of CDKN2A, CDKN1A, CCND1 and CDKN1B mutations in OSCC from The Cancer Genome Atlas (TCGA) database
Mutations in aforementioned genes from the 172 TCGA cohort are shown in Fig. 3. The most frequent mutated genes were CDKN2A affecting 90 patients (52%) and CCND1 affecting 26 patients (15.12%), specifically the most frequent variations in CDKN2A were truncating mutations and deep deletions, whereas in the case of CCDN1 gene mutations were mostly amplifications (Fig. 3A). Network involved in the expression of selected CPs is displayed in Fig. 3B.
In an OS analysis within TCGA database, OSCC cases with alterations in selected genes reached a total of 65 cases of which 31 died with a 17.91 median month survival. In the other hand, OSCC cases without alterations in aforementioned genes reached a total of 37 cases of which 16 deceased with a 28.29 median month survival. This OS tendency was not statistically significant by means of Kaplan-Meier method (log-rank test -value 0.414).
Discussion
The oncogenic role of aberrant signalling pathways that regulate cell division has been investigated in various malignancies including OSCC [1, 2, 3, 4]. The aim of this study was to identify the inmunoexpression signature of four canonical CPs in our institutional database and to integrate these data with genetic information available from TCGA project. Available data gathered from literature presents certain gaps of knowledge regarding the implications of these markers expression towards cancer-related features and survival. To the best our knowledge, only three meta-analyses has been previously published by pooling data from studies evaluating the significance of three out of the four selected CPs (p16, cyclin D1 and p27) as a prognostic factor in OSCC patients [16, 17, 18]. Sedghizadeh et al. provided a pooled analysis regarding OS hazard ratios for p16 patients versus p16-concluding a favourable survival rate for p16 (HR 0.360; 95% CI 0.258–0.503; 0.001) [16]. A strong correlation between human papilloma virus (HPV) infection and p16 protein expression has consistently been reported in head and neck cancers. This molecular event is triggered by pRb inactivation by the HPV E7 oncoprotein. In this vein, p16 protein expression is part of the current medical gold standard algorithm to stablish the presence of an HPV-induced carcinogenesis in head and neck and tumours [12]. Curiously, Ang et al. demonstrated that p16 expression is a stronger independent prognostic factor for OS than the presence of HPV DNA [13]. CDKN2A is the p16 related gene, is located on chromosome 9p21, and has three exons. The most frequent mutations detected on this gene according to TCGA are deep deletions [15] (Fig. 3). Padhi et al. verified that this loss of function has been linked per se to a poor prognosis and survival rate in OSCC [19]. The present report failed to ascertained the implications of this protein as a valuable tool for OSCC prognosis. Scully and Bedi reported that ethnic and geographical difference in oral cancer affected populations could induce an accentuated risk of bias for result interpretation [20]. Taking in account this hypothesis two close studies emerge. González-Moles et al. observed that 41.67% of the tumours studied had negative expression for both of p21 and p16 proteins but do not found significant difference when they associated their expression with clinical-pathological parameters, including recurrence and survival [21]. On the other hand, Monteiro et al. do found p16 as a useful tool for prognosis when studying cancer-specific survival ( 0.019); it is worth mentioning the moderate cut-off point variation among mentioned studies and the present [22]. In the present report, this OS tendency is evidenced although it does not reach statistical significance (Table 3).
P21 and p27 are inhibitors of the CIP/KIP family, whose function is the inhibition of DNA replication and cell proliferation. The loss of expression of these proteins has been related to worse OSCC prognosis [3, 4]. Evidences are particularly stronger towards p27 expression; Gao et al. performed a pooled analysis on its prognosis significance verifying that p27 low expression was significantly correlated with poorer OS for OSCC patients (HR 0.743 (95% CI 0.62–0.89, 0.002) [18]. Evidences toward the effect of P21 in OSCC prognosis is much more diffuse as evidenced by our group on a comprehensive review [10]. Choi et al. obtained an inverse correlation tendency between the two markers ( 0.08), although this group failed to observe any relation with clinical-pathological parameters [23]. Zhang et al. also obtained a statistically significant correlation between the expression of both CPs through Pearson analysis [24]. Fillies et al. did not observe any significant result regarding OS through Cox regression models [5]. Kapranos et al. analysing tumours of the oral cavity and larynx at the same time, found no correlation on their inmunoexpression, nor obtained differences in survival in any combinations of expression [25].
A bulk of research has ascertained the pivotal role of cyclins in oral carcinogenesis, specially the role of Cyclin D1. Cyclin D1 turnover and nuclear accumulation is regulated by NF-B pathway activation and the inactivation/phosphorylation of GSK3b. A recent meta-analysis involving 2942 patients correlated its overexpression with worse OS for OSCC patients (HR 2.00, 95% CI 1.59–2.51, 0.001) [17]. The main underlying mechanism explaining this rationale is amplification of the CCND1 gene, which in turns was verified in the present study via TCGA database analysis (Fig. 3) [15]. Nonetheless, other interferers can modulate its transcription; of which the best characterised are AP-1, ETS1, -Catenin, c-Myc, ETS1, and STAT3/5A [17]. Pignataro et al. studied the correlation of p27 and Cyclin D1 in head and neck squamous cell carcinoma finding it inverse-the majority of tumours that show positive expression of p27, show negative expression of Cyclin D1 and tumours with positive expression of Cyclin D1, present negative expression of p27Kip1. This group also noticed that Cyclin D1/ tumours have a worse prognosis and that Cyclin D1-/p27 in terms of OS ( 0.0015) and recurrence-free survival ( 0.0001) [26]. In this vein, Rodolico et al. also observed that tumours with overexpression of Cyclin D1 and with subexpression of p27 are related to the appearance of lymph node metastases in patients with squamous cell carcinomas of the lower lip [27].
Finally, authors would like to point out a few limitations. The discordance with previously published studies may be a result of geographic variations and ethnic disparity that result in different etiological factors for OSCC and also due to differences related to technical procedures such as the use of different monoclonal antibodies, cut-off points for positivity interpretation, or the use of univariate vs. multivariate analysis.
Overall, the present report ascertained neither the presence nor the degree of CPs expression influenced OS in the present OSCC cohort. Integration with TCGA database supported the presence of moderate cell cycle variations at a phenotype and genotype level in OSCC.
Footnotes
Acknowledgments
A.I.L.P, and C.M.C.P are supported by a fellowship from the Health Research Institute of Santiago de Compostela (IDIS).
References
1.
HaP.K. and CalifanoJ.A., The molecular biology of mucosal field cancerization of the head and neck, Critical Reviews in Oral Biology and Medicine14 (2003), 363–369.
2.
AliJ.SabihaB.JanH.U.HaiderS.A.KhanA.A. and AliS.S., Genetic etiology of oral cancer, Oral Oncology70 (2017), 23–28.
3.
ShackneyS.E. and ShankeyT.V., Cell cycle models for molecular biology and molecular oncology: Exploring new dimensions, Cytometry35 (1999), 97–116.
4.
BessonA.DowdyS.F. and RobertsJ.M., CDK inhibitors: Cell cycle regulators and beyond, Developmental Cell14 (2008), 159–169.
5.
FilliesT.WolteringM.BrandtB.Van DiestJ.P.WerkmeisterR.JoosU. et al., Cell cycle regulating proteins p21 and p27 in prognosis of oral squamous cell carcinomas, Oncology Reports17 (2007), 355–359.
6.
SchoelchM.L.RegeziJ.A.DekkerN.P.NgI.O.McMillanA.ZioberB.L. et al., Cell cycle proteins and the development of oral squamous cell carcinoma, Oral Oncology35 (1999), 333–342.
7.
ShintaniS.MiharaM.NakaharaY.KiyotaA.UeyamaY.MatsumuraT. et al., Expression of cell cycle control proteins in normal epithelium, premalignant and malignant lesions of oral cavity, Oral Oncology38 (2002), 235–243.
8.
Gleber-NettoF.O.BraakhuisB.J.TriantafyllouA.TakesR.P.KelnerN.RodrigoJ.P. et al., Molecular events in relapsed oral squamous cell carcinoma: Recurrence vs. secondary primary tumor, Oral Oncology51 (2015) 738–744.
9.
Perez-SayansM.Suarez-PenarandaJ.M.Gayoso-DizP.Barros-AngueiraF.Gandara-ReyJ.M. and Garcia-GarciaA., p16(INK4a)/CDKN2 expression and its relationship with oral squamous cell carcinoma is our current knowledge enough? Cancer Letters306 (2011), 134–141.
10.
Perez-SayansM.Suarez-PenarandaJ.M.Gayoso-DizP.Barros-AngueiraF.Gandara-ReyJ.M. and Garcia-GarciaA., The role of p21Waf1/CIP1 as a Cip/Kip type cell-cycle regulator in oral squamous cell carcinoma (Review), Medicina Oral Patolologia Oral Cirugia Bucal18 (2013), 219.
11.
De AlmeidaM.R.Perez-SayansM.Suarez-PenarandaJ.M.Somoza-MartinJ.M. and Garcia-GarciaA., p27Kip1 expression as a prognostic marker for squamous cell carcinoma of the head and neck, Oncology Letters10 (2015), 2675–2682.
12.
Perez-SayansM.Suarez-PenarandaJ.M.Padin-IruegasM.E.Gayoso-DizP.Reis-De AlmeidaM.Barros-AngueiraF. et al., The loss of p16 expression worsens the prognosis of OSCC, Applied Immunohistochemistry and Molecular Morphology23 (2015), 724–732.
13.
Perez-SayansM.Suarez-PenarandaJ.M.IruegasE.P.de AlmeidaM.R.Barros-AngueiraF.TorreiraM.G. et al., p21{Waf1/CIP1} is a poor diagnostic and prognostic marker for OSCC although its expression increases in patients with N1 regional metastasis, Cancer Biomarkerks15 (2015), 19–26.
14.
Cancer Genome Atlas Network, Comprehensive genomic characterization of head and neck squamous cell carcinomas, Nature517 (2015), 576–582.
15.
von ElmE.AltmanD.G.EggerM.PocockS.J.GotzscheP.C.VandenbrouckeJ.P. et al., The strengthening the reporting of observational studies in epidemiology (STROBE) statement: Guidelines for reporting observational studies, International Journal of Surgery12 (2014), 1495–1499.
16.
SedghizadehP.P.BillingtonW.D.PaxtonD.EbeedR.MahabadyS.ClarkG.T. et al., Is p16-positive oropharyngeal squamous cell carcinoma associated with favorable prognosis? A systematic review and meta-analysis, Oral Oncology54 (2016), 15–27.
17.
Ramos-GarciaP.Gonzalez-MolesM.A.Gonzalez-RuizL.Ruiz-AvilaI.AyenA.Gil-MontoyaJ.A., Prognostic and clinicopathological significance of cyclin D1 expression in oral squamous cell carcinoma: A systematic review and meta-analysis, Oral Oncololy83 (2018), 96–106.
18.
GaoL.GuW.ZhengJ.RenW.ChangS.WangX. et al., Clinicopathological and prognostic significance of p27 expression in oral squamous cell carcinoma: A meta-analysis, International Journal of Biological Markers28 (2013), 329.
19.
PadhiS.S.RoyS.KarM.SahaA.RoyS.AdhyaA. et al., Role of CDKN2A/p16 expression in the prognostication of oral squamous cell carcinoma, Oral Oncology73 (2017), 27–35.
20.
ScullyC. and BediR., Ethnicity and oral cancer, Lancet Oncology1 (2000), 37–42.
21.
Gonzalez-MolesM.A.Gil-MontoyaJ.A.Ruiz-AvilaI.EstebanF.Delgado-RodriguezM. and Bascones-MartinezA., Prognostic significance of p21WAF1/CIP1, p16INK4a and CD44s in tongue cancer, Oncology Reports18 (2007), 389–396.
22.
MonteiroL.S.Diniz-FreitasM.WarnakulasuriyaS.Garcia-CaballeroT.FortezaJ. and FragaM., An immunohistochemical score to predict the outcome for oral squamous cell carcinoma, Journal of Oral Pathology and Medicine47 (2018), 375–381.
23.
ChoiH.R.TuckerS.A.HuangZ.GillenwaterA.M.LunaM.A.BatsakisJ.G. et al., Differential expressions of cyclin-dependent kinase inhibitors (p27 and p21) and their relation to p53 and Ki-67 in oral squamous tumorigenesis, International Journal of Oncology22 (2003), 409–414.
24.
ZhangM.LiJ.WangL.TianZ.ZhangP.XuQ. et al., Prognostic significance of p21, p27 and survivin protein expression in patients with oral squamous cell carcinoma, Oncology Letters6 (2013), 381–386.
25.
KapranosN.StathopoulosG.P.ManolopoulosL.KokkaE.PapadimitriouC.BibasA. et al., P53, P21 and P27 protein expression in head and neck cancer and their prognostic value, Anticancer Research21 (2001), 521–528.
26.
PignataroL.SambataroG.PaganiD. and PruneriG., Clinico-prognostic value of D-type cyclins and p27 in laryngeal cancer patients: A review, Acta Otorhinolaryngologica Italica25 (2005), 75–85.
27.
RodolicoV.AragonaF.CabibiD.Di BernardoC.Di LorenzoR.GebbiaN. et al., Overexpression of cyclin D1 and interaction between p27Kip1 and tumour thickness predict lymph node metastases occurrence in lower lip squamous cell carcinoma, Oral Oncology41 (2005), 268–275.