: 14-3-3 protein plays an important role in multiple cellular processes. The role of 14-3-3 in the progression of intrahepatic cholangiocarcinoma (ICC) has not been well understood.
OBJECTIVE:
We performed this research to explore the relationship between 14-3-3 level and clinical characteristics and prognosis of ICC patients. Besides, we used ICC cell lines HCCC-9810 and RBE to assess the biological function of 14-3-3.
METHODS:
We examined 14-3-3 expression in 28 ICC tissues and matched paratumor tissues by quantitative real-time PCR and immunohistochemistry. Additionally, ICC tissue array from 100 patients and normal liver tissue array from 24 healthy people were also analyzed by immunohistochemistry. 14-3-3 was knocked down in ICC cell lines and the functions and mechanisms of 14-3-3 were assessed.
RESULTS:
14-3-3 is highly expressed in ICC tissues and high expression of 14-3-3 correlates poor overall survival in ICC patients. Knocking down of 14-3-3 in ICC cell lines reduced cells migration, invasion and anoikis resistance. Furthermore, 14-3-3-silenced ICC cells showed significantly decreased invasion-related protein MMP2 and MMP9 expression.
CONCLUSIONS:
Our results demonstrate prognostic value of 14-3-3 and its role in metastasis, which is associated with ICC cell lines migration, invasion and anoikis resistance.
Cholangiocarcinoma (CC) is the second most common tumor of primary liver cancers in the world. It is generally classified into three forms including intrahepatic cholangiocarcinoma (ICC), hilar cholangiocarcinoma and extrahepatic cholangiocarcinoma(ECC) [26]. ICC is an aggressive malignancy with very poor prognosis and increased incidence worldwide recently. Regardless of the advances in diagnostic and therapeutic procedures for liver cancers, the prognosis of ICC patients remains pessimistic [22, 31, 40]. So a better understanding of the molecular mechanisms in the progression of ICC will improve the prognosis of ICC patients.
14-3-3 (also called stratifin) is a member of a highly conserved family of 14-3-3 proteins [2, 42]. It is clear that 14-3- plays an important role in many biological functional pocesses of cells, including negative regulation of cell cycle progression [1, 10], regulation of signal transduction pathways [17] and modulation of cell growth [6], differentiation [12] and apoptosis [5, 27]. 14-3-3 has been considered a tumor suppressor due to its role in protecting p53 from MDM2-mediated ubiquitination and degradation and in mediating a G2 /M checkpoint following DNA damage [37, 38]. Downregulation of 14-3-3 expression has been observed in various human cancers [13, 23, 33, 36]. However, many studies have reported that 14-3-3 overexpression predicted poor prognosis of pancreatic [20], lung [18, 25, 30], colorectal [7] and liver neoplasms [21, 41], and that its expression contributed to resistance to DNA-damaging drugs in gastric, breast and colorectal cancers [8, 15, 43]. This raises the double-edged sword role of 14-3-3 in human cancers [19, 36].
So far, little is known about the role of 14-3-3 in ICC. In this study, we reported the expression of 14-3-3 in ICC tissue and analyzed the prognosis of ICC. Besides, we also used ICC cell lines to investigate the expression of 14-3-3 and to assess the biological function of the 14-3-3 protein. Our results suggest that the reduction in 14-3-3 expression suppresses ICC cell migration and invasion by decreasing MMP2 and MMP9 expression in vitro. Moreover, 14-3-3 knockdown enhances anoikis of ICC cells. In summary, our study investigates the prognostic value of 14-3-. We are the first to investigate the role of 14-3-3 in metastasis of ICC patients, and to investigate its mechanim, which contributes to ICC cells migration, invasion and anoikis resistance.
Materials and methods
Clinical specimen collection
Samples from 28 patients receiving hepatic resection between 2008 and 2011 were obtained from patients with ICC at the First Affiliated Hospital of Zhejiang University (Zhejiang, China). Moreover, intrahepatic cholangiocarcinoma tissue array from 100 patients and normal liver tissue array from 24 healthy persons between 2008 and 2014 were also obtained from the First Affiliated Hospital of Zhejiang University. The research was approved by the First Affiliated Hospital of Zhejiang University Ethics Committee, and informed consent was obtained from each patient following institutional review board protocols. Patient charts were reviewed to obtain clinical data about age, gender, tumor size, AFP, and tumor differentiation. The histopathologic diagnosis was based on World Health Organization criteria. The histological grade of tumor differentiation was determined according to the classification proposed by Edmondson and Steiner. Overall survival (OS) was defined as the interval between the dates of surgery and death.
Cell culture and transfection
Human ICC cell lines (HCCC-9810, RBE) were purchased from Cell Bank of Type Culture Collection of Chinese Academy of Sciences and were cultured in 1640 complete medium supplemented with 10% fetal bovine serum (Gibco, USA) and penicillin/streptomycin. For siRNA transfection, cells were seeded on 6-well tissue culture plates with 40% confluence. The next day, cells were transfected with 100 nM siRNA targeting human 14-3-3 or control siRNA (Shanghai GenePharma Co., China) using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer’s instructions. For 14-3-3, targetcoding sequences of siRNA were 5’ UGAAGAU GAAGGGU GACUATT 3’ and 5’ UAGUCACCCUUCAUCUUCATT 3’.
Immunohistochemistry
The paraffin-embedded tissue that used for the HE-stained section was chosen for immunohistochemistry. They were cut at m, deparaffinized in xylene, and rehydrated in a series of graded alcohol dilutions. Heat epitope retrieval was carried out for 20 minutes in target-retrieval solution at pH 7.5. Histological sections were immunostained using a rabbit monoclonal antibody anti-14-3-3 (tyle ID: HPA011105, USA) at dilution of 1:100 overnight at 4C. Slides were then incubated with HRP at room temperature for 30 minutes and were visualized using DAB as chromogen for 5–10 minutes. Sections were scored semiquantitatively as follows: (negative), 0% immunoreactive cells; ( weak) 25% immunoreactive cells; ( moderate) 25%–50% immunoreactive cells; ( strong) 50% immunoreactive cells. For statistical purposes, cases with scores negative and were considered low expression and those with scores and were considered high expression.
Western blot analysis
Soluble proteins were collected after cells were lysed. Protein concentration was determined by the Bradford assay (Bio-Rad). Equal amounts of lysate were applied to a 10% NuPAGE Bis-Tris Gel (Invitrogen) and electrophoresed. Then the lysates were transferred onto PVDF membrane for 90min, followed by blocking in 5% skim milk. The membrane was washed once with TBST, then incubated for overnight at 4C with relevant antibodies (1:1000). The membrane was again washed three times with TBST, then incubated with secondary antibody (Goat Anti-Rabbit IgG 1:5000) for 2 h at room temperature. The membrane was washed three times with TBST and then ECL liquid was added and placed in a darkroom for the reaction to proceed. -actin (1:5000 dilution, Sigma-Aldrich) or GAPDH (1:5000 dilution, Sigma-Aldrich) was used as the loading control.
RNA extraction and quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was isolated from clinical samples using TRIzol reagent according to the manufacturer’s recommended protocol (Invitrogen). For quantitative real-time RT–PCR analysis, the Roche LightCycler was used with Takara SYBR Premix Extaq system. Primers were synthesized by Shanghai Sangon Biological Engineering Technology Services Co., Ltd. The nucleotide sequences of the primers were as follows: GAPDH, 5’-ATGGGGAAGGTGAAGGTCG-3’ and 5’-GGGGTCATTGATGGCAACAATA-3’; 14-3-, 5’-TGGGAGATCAAGATCGACCC-3’ and 5’-GG GCCATTGGCAGAGGATG-3’; Each sample was taken in triplicate. Gene expression was calculated relative to the expression of -actin in tumor cell lines or clinical samples using the 2Ct method.
Proliferation and colony formation assay
For proliferation assay, ICC cells were seeded in 96-well plates at 1000 cells/well. Cell viability and growth was determined by Cell-Counting Kit (CCK)-8 (Dojindo), reading absorbance at 450 nm according to the manufacturer’s instructions. For colony formation assay, cells were seeded into 6-well plates at a concentration of 2000 cells/well and cultured at 37C for 10–16 days according to the character of each cell line. Then, the cells were fixed with 100% methanol and stained with 0.1% crystal violet. Megascopic cell colonies were counted by Image-Pro Plus 5.0 (Media Cybernetics).
Cell cycle analysis
Cells (1 10) were fixed in 75% ethanol at 20C for 24 hours. Fixed cells were stained as the protocol of Cycle TESTTM PLUS DNA Reagent Kit (BD Biosciences) and analyzed by flow cytometry (Beckman Coulter FC 500).
Wound healing assay
Cells were seeded in a 6-well plate, grown until confluence, and then starved for 24 h to synchronize cell proliferation. After starvation, each cell monolayer was scraped three times with a sterile p200 tip, and in each wound three micrographs were taken at 0 h. On the same area of the scratch, micrographs were then taken at 48 h to measure the distance of the scratch edge using LuciaG 5.0 software (Nikon), and each time point was normalized to 0 h.
Migration and invasion assays
Cell migration and invasion assays were performed in the BD Falcon 24-multiwell insert system (BD Biosciences, San Jose, CA) according to the manufacturer’s instruction. For migration assays, cells which had been permanently transfected in serum-free medium were placed on the upper chamber with a non-coated membrane. For matrigel invasion assays, filters were precoated with 3 l Marigel (BD Biosciences) for 2 h. In both assays, culture medium containing 10% FBS was added to lower chambers and aliquots of 5 10 cells in 20 l of FBS-free meium were seeded into upper chambers. The cells were incubated for 24 h (migration assay) and 48 h (invasion assay). Non-migrated or non-invaded cells were removed from the upper surface of the filter. Cells on the lower surface of the membrane were fixed with ice-cold methanol and stained with crystal violet. Cell numbers were counted under an optical microscope. Each experiment was repeated at least three times.
Apoptosis and anoikis assay
Apoptosis was detected by use of Annexin V. The percentage of cell death was calculated as the sum of percentage of Annexin V-positive/propidium iodide-negative cells (undergoing early stages of apoptosis) and Annexin V-positive/propidium iodide-positive (undergoing late stages of apoptosis) in a total cell population. Cells were cultured on either ultra-low attachment plates (Corning, 3471) or plastic plates (Corning, 3516) for 24 h. After incubation, the suspended and adherent cells were harvested, and apoptosis was measured using AnnexinV and propidium iodide flow cytometry.
Statistical analysis
Statistical analysis was performed using SPSS 17.0 software (SPSS). Comparisons of the quantitative data were made using the Student t test or Wilcoxon signed-rank test between 2 groups. The chi-square test or Fisher’s exact test was used to evaluate any potential association between the 14-3-3 expression and the clinic pathologic parameters. Statstical significance was set at 0.05.
Results
14-3-3 is high level expressed in IC patients
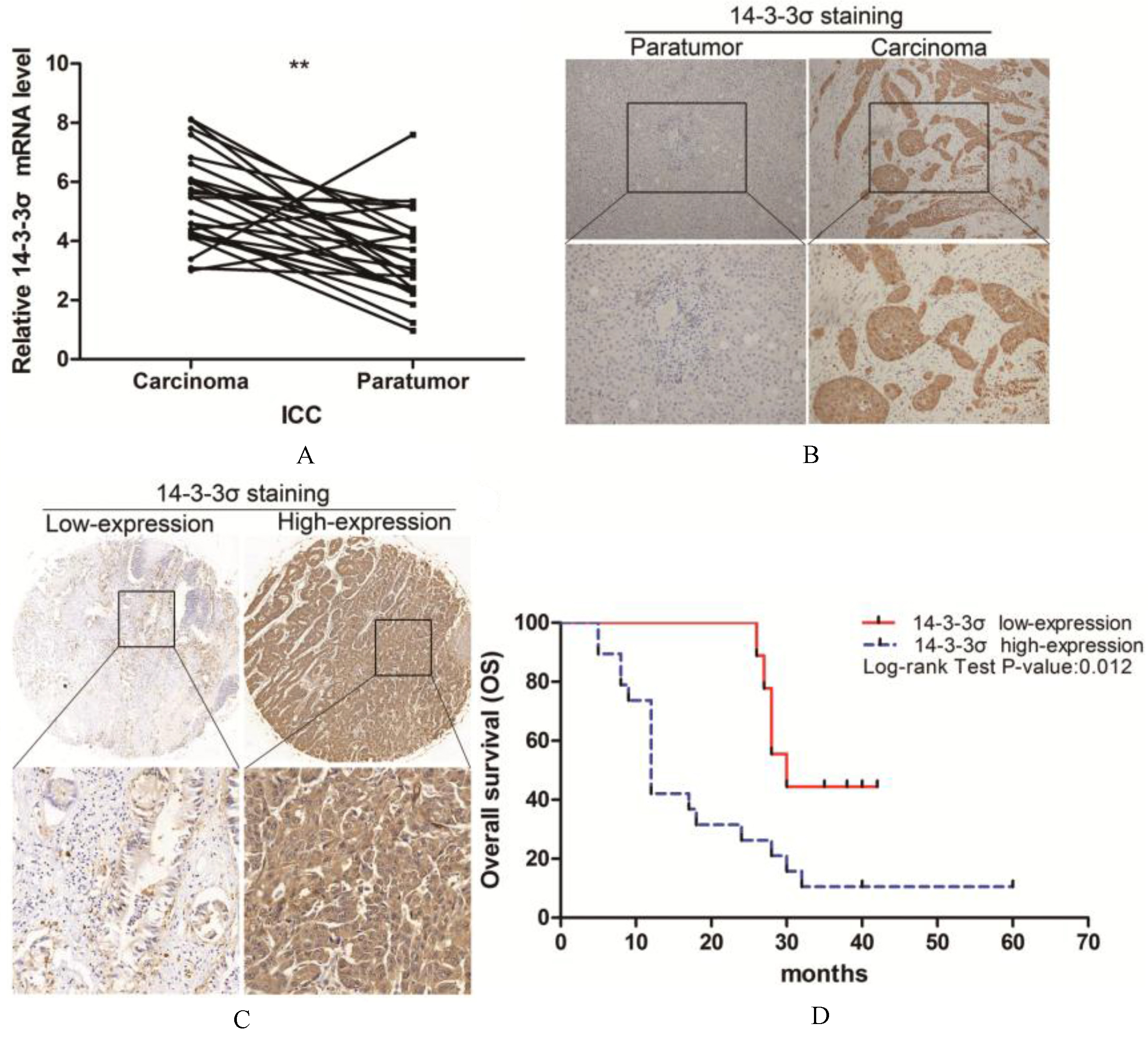
To analyze the expression of 14-3-3 in ICC, 28 patients’ tumor tissues and tissues adjacent to the cancer were examined by quantitative real-time PCR. As shown in (Fig. 1A), the level of 14-3-3 mRNA was significantly higher in ICC than in adjacent tissues ( 0.01). Similarly, assessment via immunohistochemistry (IHC) revealed that 14-3-3 was upregulated in ICC tumors compared with adjacent tissues (Fig. 1B). A total of 67.8% (19/28) samples of the tumor tissue showed overexpression of 14-3-3 compared with adjacent tissue.
14-3-3 is highly expressed in ICC tissues and predicts poor prognosis of patients. (A) Quantitative real-time PCR analysis of 14-3- expression in 28 paired human ICCs and paratumor tissues. (B) Immunohistochemical (IHC) staining of 14-3- expression in 28 paired human ICCs and paratumor tissues. The representative views were shown. (C) Comparison of 14-3-3 levels in 100 human ICCs tissue array using IHC staining and scoring. Representative image was shown. (D) Overall survival time after surgery of the 28 human ICCs patients were compared between the “14-3-3 low” and “14-3-3 high” groups, 0.05.
To verify this result, we studied the expression of 14-3-3 in ICC tissue array and normal liver tissue array. In ICC tissue microarray, 16.7% (16/96) were negative or weakly positive and 83.3% (80/96) were moderately or strongly positive. In normal liver tissue array, only 16.7% (4/24) were strongly positive (Table 1).
Immunohistochemistry of 14-3-3 in tissue array (p 0.001)
14-3-3
14-3-3
Total
Low-expression
High-expression
Normal liver tissue
20
4
24
ICC tissue
16
80
100
Total
36
84
120
Abbreviations: ICC, intrahepatic cholangiocarcinoma. Statistical analyses were performed with chi-square test.
We then analyzed the relationship between 14-3-3 expression and clinic pathologic features of the tumors. Patients’ characteristics were shown in Tables 2 and 3, 14-3-3 staining was significantly associated with tumor differentiation ( 0.001), lymph node metastasis ( 0.026) and blood vessel invasion ( 0.023). Thus, 14-3-3 expression appears to be a valuable predictive factor for tumor metastasis and ICC aggressiveness.
Clinical Specimen Collection Samples and tumor characteristics by 14-3-3 stage
Characteristics
14-3-3
14-3-3
P-value
Low-expression
High-expression
( 9)
( 19)
Age(years)
61.2 12.6
57.8 4.6
0.459
Gender
0.317
Male
8
13
Female
1
6
Vascular invasion
0.023
Negative
9
8
Positive
0
12
Tumor size (cm)
1.000
5
2
6
5
7
13
AFP (ng/ml)
0.321
20
8
19
20
1
0
CA-199(U/ml)
0.371
39
4
4
39
5
15
Abbreviations: AFP, -fetoprotein; CA-199, carbohydrate antigen 19-9. Statistical analyses were performed with the Student’s t-test or Fisher’s exact test. 0.05.
Tissue array and tumor characteristics by 14-3-3 stage
Characteristics
14-3-3
14-3-3
Low-expression
High-expression
P-value
( 16)
( 80)
Age (years)
50.5 10.7
55.4 10.9
0.119
Gender
0.780
Male
9
48
Female
7
32
Lymph node
0.026
Negative
14
46
Positive
2
34
TNM stage (AJCC)
0.285
I and II
9
33
III and IV
7
47
Tumor differentiation
0.001
I and II
8
7
III and IV
8
73
Abbreviations: TNM, tumor-node-metastasis; Statistical analyses were performed with the Student’s t-test or chi-square test. 0.05; 0.01.
Survival analysis in patients with ICC
To assess the prognostic value of 14-3-3 expression, Kaplan-Meier survival analysis was performed in the 28 patients surviving more than 1 month after surgery. The median survival time based on 28 subjects was 24.0 months. As shown in Table 4 and Fig. 1D, for subjects with low expression of 14-3-3 ( 9), the median survival time was 30.0 months (95%CI 24.3, 35.8). In sharp contrast, for subjects with high expression of 14-3-3 ( 19) the median survival time was 12.0 months (95%CI 9.9, 14.1). These data indicate that 14-3-3 is associated with a significant reduction in survival.
Relationship between 14-3-3 expression and survival rate
Medians for Survival Time (months)
Estimate
95% Confidence Interval
14-3-3 low-expression
30
(24.156, 35.844)
14-3-3 high-expression
12
(9.891, 14.109)
Overall
24
(11.036, 36.964)
Statistical analyses were performed with the Kaplan-Meier survival analysis.
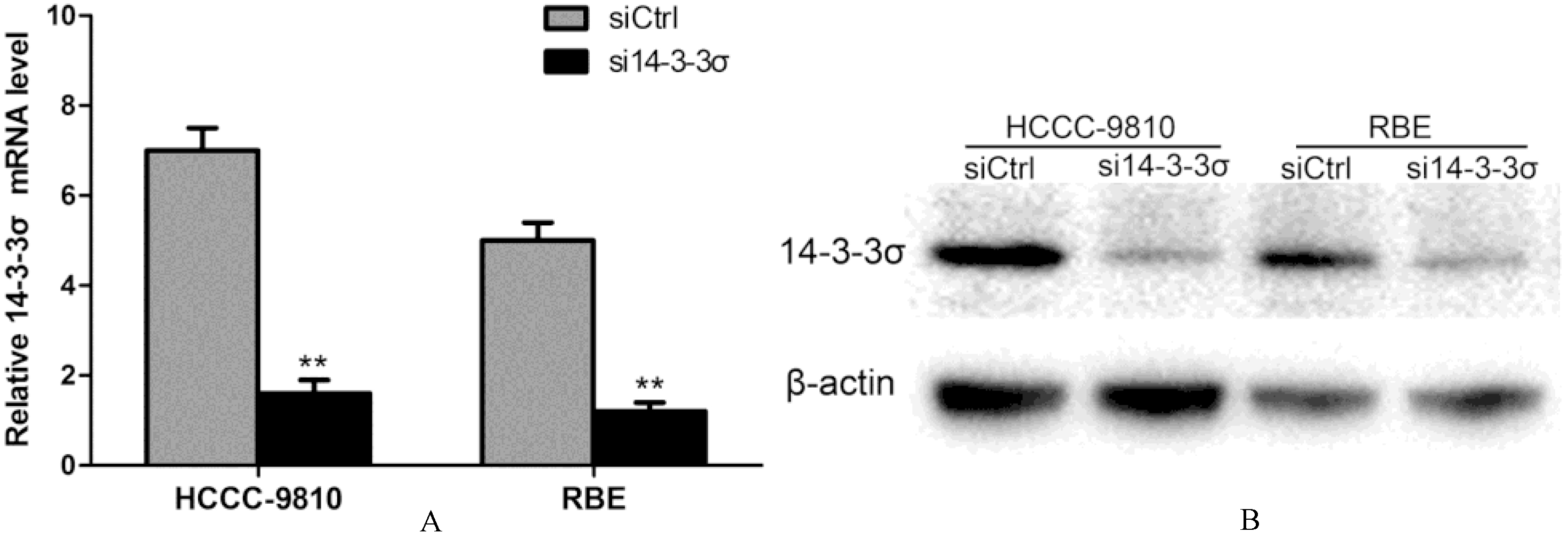
Establishment of 14-3-3 knockdown ICC cell lines. (A) HCCC-9810 and RBE cells were transfected with control siRNA or 14-3--specific siRNA, and the expression of 14-3- was measured by quantitative real-time PCR. (B) The protein levels of 14-3- knockdown ICC cell lines were measured by Western blotting. -actin was used as a control. Experiments were performed three times. 0.01.
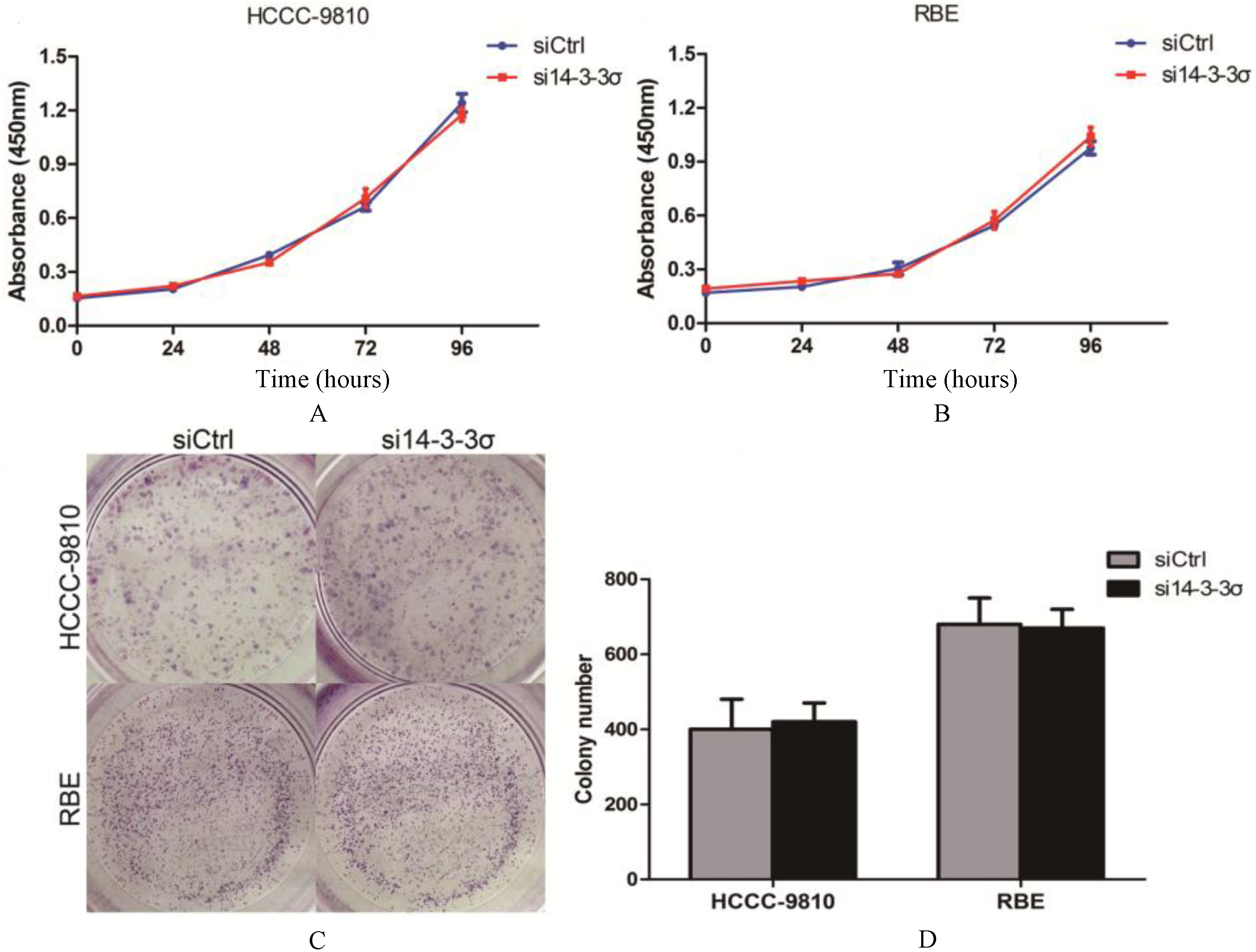
14-3-3 knockdown has no efect on ICC cell lines proliferation. (A) Proliferation rates of 14-3- silenced HCCC-9810 cells and control HCCC-9810 cells were evaluated by Cell Counting Kit 8 Assay. (B) Proliferation rates of 14-3-3 silenced RBE cells and control RBE cells were evaluated by Cell Counting Kit 8 Assay. (C and D) Colony formation analysis of 14-3-3 silenced ICC cell lines and control cells.
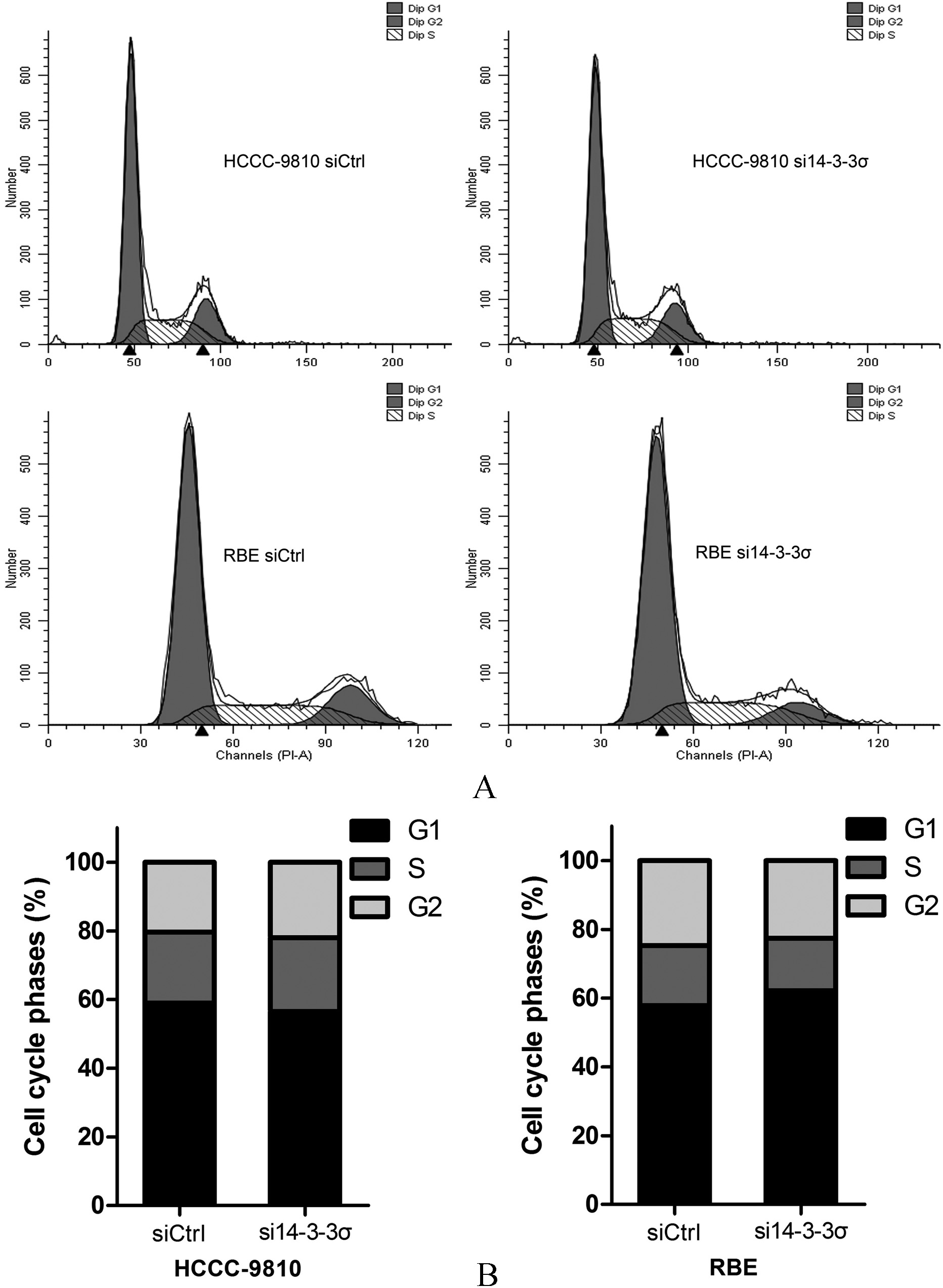
14-3-3 knockdown did not remarkably lead to cell cycle arrest in ICC. (A and B) Analysis of cell cycle distribution in HCCC-9810 and RBE cells transfected with control siRNA or 14-3-3-specific siRNA demonstrates no remarkable change in G1 and G2 fractio. Experiments were performed three times. 0.05.
Establishment of 14-3-3 knockdown ICC cell lines
To study the mechanism of 14-3-3 in ICC, we established 14-3-3 knockdown cell lines. HCCC-9810 and RBE cells were transfected with 14-3-3 small interfering RNA (siRNA) and negative control siRNA. 14-3-3 mRNA level was measured by quantitative RT–PCR. Compared with the control group, 14-3- transcripts were significantly reduced in 14-3--siRNA-transfectedcells ( 0.01) (Fig. 2A). Accordingly, the 14-3-3 protein levels were markedly decreased as shown by Western blotting (Fig. 2B). These results demonstrate that 14-3-3-siRNA transfection leads to the reduction in 14-3-3 expression.
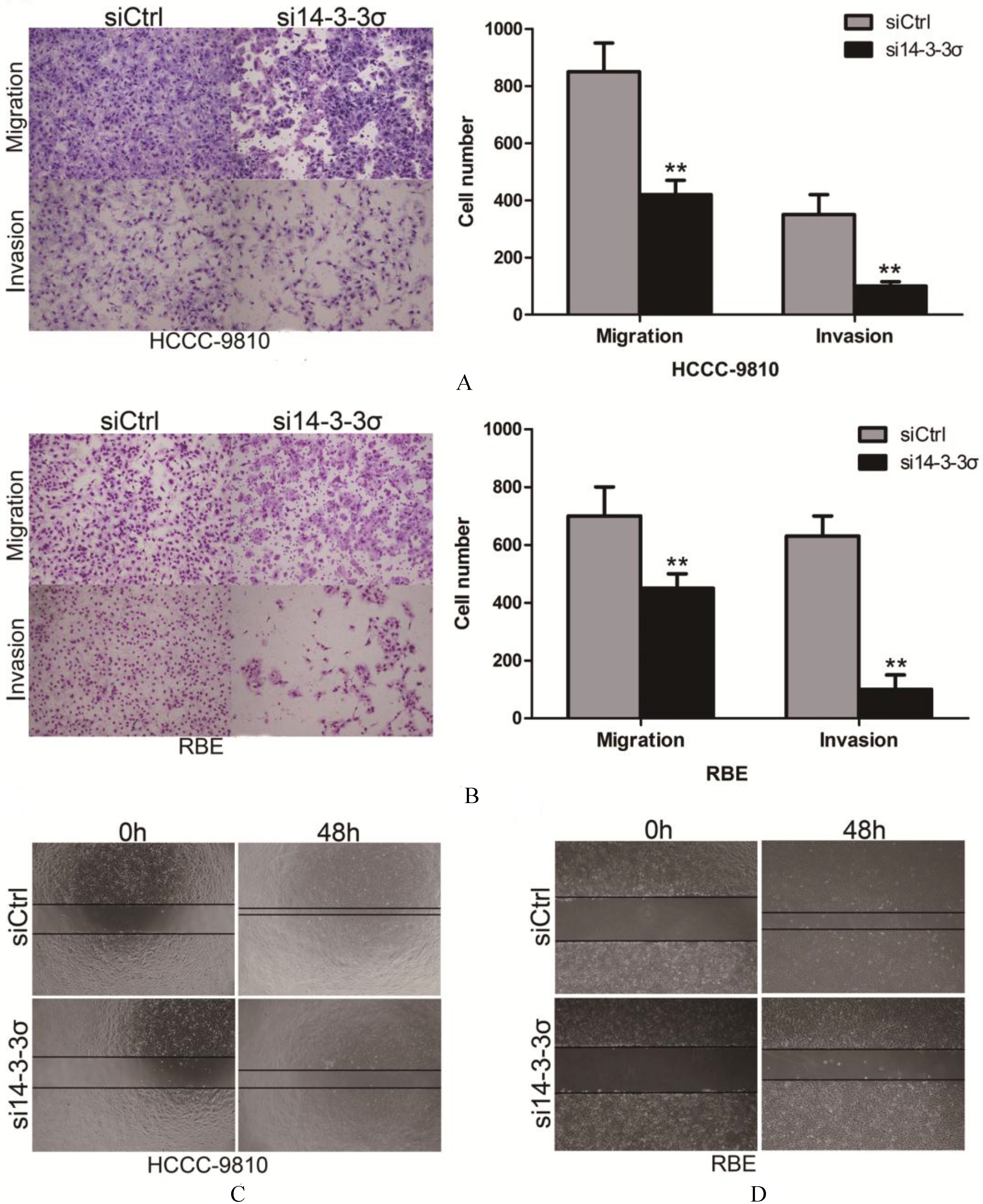
14-3-3 knockdown inhibits migration and invasion of ICC cell lines. (A and B) Migration and invasion of 14-3- knockdownHCCC-9810 and RBE cells was measured by a transwell assay. Results were quantitated by counting migrating and invasive cells in five randomly chosen high-power fields for each replicate. (C) Wound healing assay was performed to compare the migratory properties of 14-3- silenced ICC cell lines and control cells. Experiments were performed three times. 0.01.
14-3-3 knockdown has no effect on ICC cell lines proliferation
We employed ICC cell lines HCCC-9810 and RBE to determine the biological function of 14-3-3 by knocking down using siRNA targeting 14-3-3. However, we found no difference in proliferative rates among the ICC cell lines and the 14-3-3 silenced clones (Figs 3A and B). Moreover, 14-3-3 depletion did not form fewer or smaller colonies compared to the control groups (Figs 3C and D). Since 14-3-3 is a cell cycle regulator and has been implicated in G2/M cell cycle arrest by p53 [1, 10], we conducted cell cycle analysis. However, our results showed that 14-3-3 knockdown did not remarkably lead to cell cycle arrest in HCCC-9810 and RBE cells (Fig. 4).
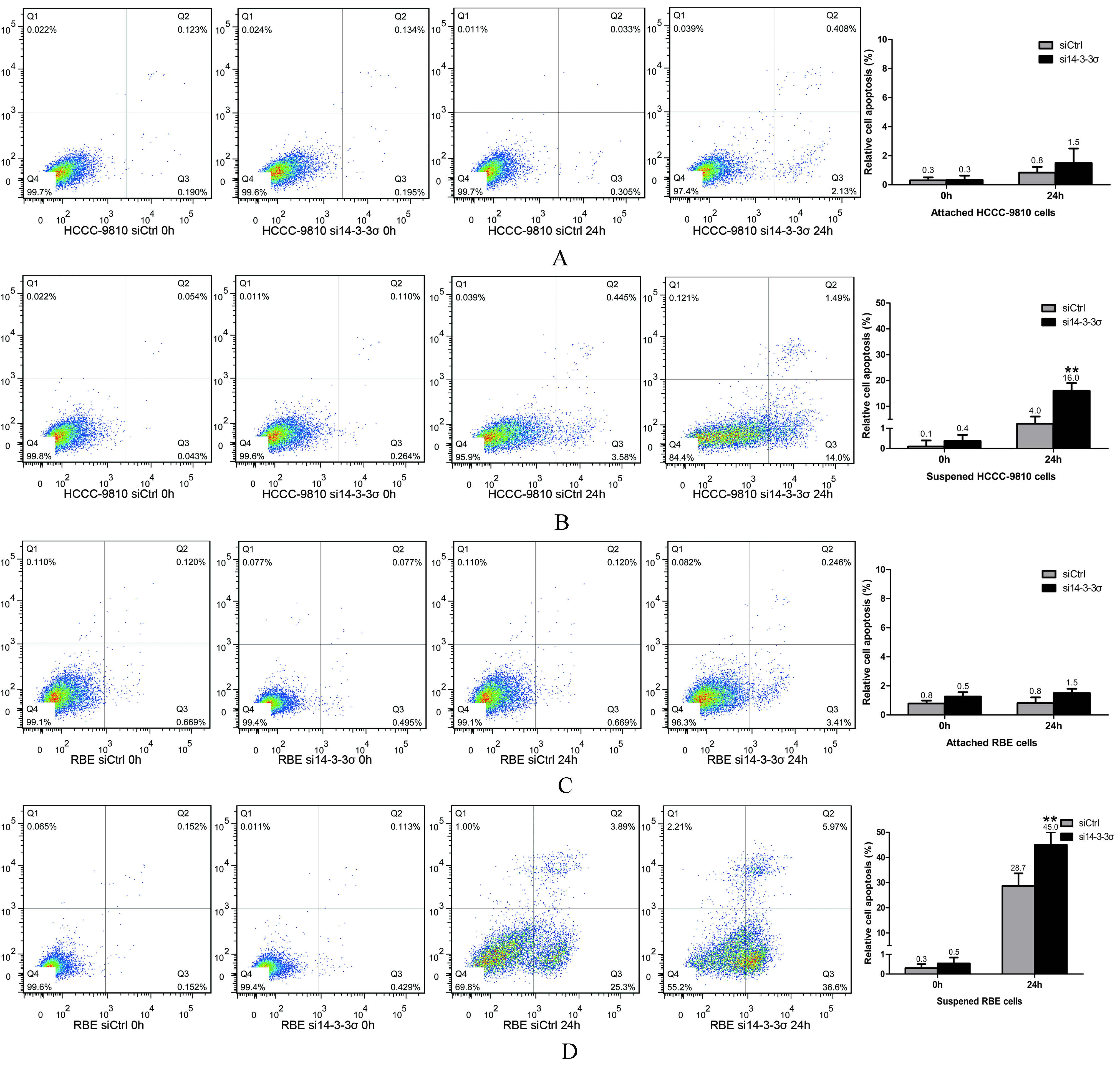
14-3-3 knockdown attenuates anoikis resistance of ICC cell lines. (A) The in vitro anoikis assay indicated that 14-3-3knockdown significantly increased the apoptosis rate of HCCC-9810 cells when cells are suspended for 24 h. (B) 14-3-3 knockdown did not induce apoptosis of attached HCCC-9810 cells. (C) The in vitro anoikis assay indicated that 14-3-3 knockdown significantly increased the apoptosis rate of RBE cells when cells are suspended for 24 h. (D) 14-3-3 knockdown did not induce apoptosis of attached RBE cells. Experiments were performed three times. 0.01.
14-3-3 knockdown inhibits migration and invasion of ICC cell lines
To elucidate the role of 14-3-3 in ICC metastasis, we evaluated the effect of 14-3-3 down-regulation on the cell migration of HCCC-9810 and RBE cells. As shown in Figs 5A and B, both 14-3-3 deficient HCCC-9810 and RBE cells (transfection with the 14-3-3 siRNA) displayed impaired invasive ability compared to those transfected with control siRNA ( 0.01). Matrigel invasion chamber assay was also performed following 14-3-3 knockdown. Consistently, the 14-3-3-siRNA-transfected cells showed a remarkably decreased invasiveness compared to the controls ( 0.01). We next used a wound healing assay to verify this result. Compared with the 14-3-3 silenced clones, the controls rapidly reduced the distance between the wound edges as illustrated in Figs 5C and D.
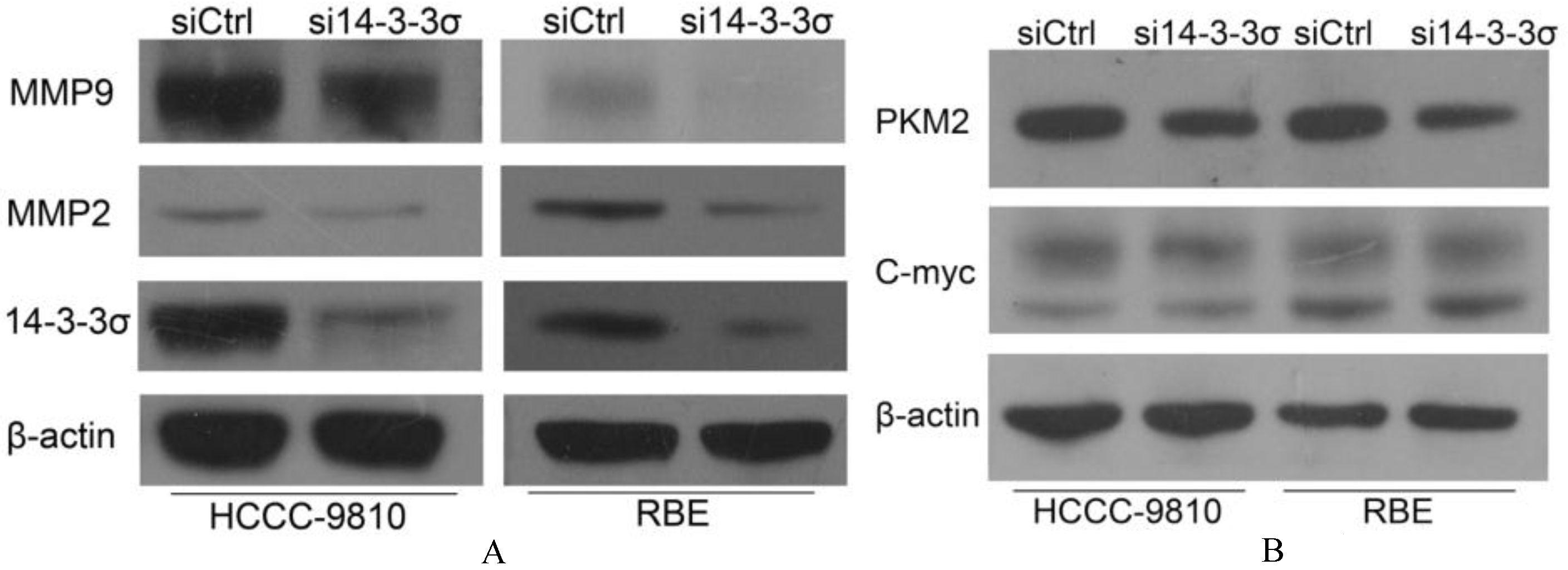
14-3-3 knockdown suppresses expression of MMP2, MMP9 and pyruvate kinase isozymes M2 (PKM2). (A) Effects of 14-3-3 knockdown on MMP-2 and MMP-9 expression were measured by Western blotting. (B) Effects of 14-3-3 knockdown on C-myc and PKM2 expression were measured by Western blotting. -actin was used as a control. Experiments were performed three times.
14-3-3 knockdown enhances anoikis of ICC cells
Anoikis resistance is responsible for the survival of tumor cells when they detach from the extracellular matrix (ECM) and disseminate into the circulatory system [29]. We found that 14-3--specific siRNA can trigger anoikis of ICC cells (Fig. 6). The apoptosis rates of suspended HCCC-9810 cells were 4.06 1.00% at 24 h after cell detachment. When 14-3-3 was inhibited, the apoptosis rates of suspended of HCCC-9810 cells significantly increased to 19.96 1.9% at 24 h. By contrast, 14-3--specific siRNA did no induce the apoptosis of attached HCCC-9810 cells (Figs 6A and B). We then repeated the anoikis assay in RBE cells. After inhibition of 14-3-3, the apoptosis rates of suspended RBE cells also significantly increased. Indeed, when downregulation of 14-3-3 was enforced, the attached RBE cells did not undergo more apoptosis compared to the control group (Figs 6C and D). Thus, 14-3-3 protein plays a crucial role in anoikis resistance of ICC cells.
14-3-3 knockdown suppresses expression of MMP2 and MMP9
MMP2 and MMP9 play key roles in the invasion of ICC cell. To investigate whether 14-3-3 could regulate the expression of MMP2 and MMP9, HCCC-9810 and RBE were treated with 14-3-3-specific siRNA and the changes of MMP2 and MMP9 protein expression was measured by Western blot. As shown in Fig. 7A, 14-3-3 knockdown significantly suppresses the expression of both MMP2 and MMP9 compared with the control groups.
14-3-3 knockdown suppresses expression of pyruvate kinase isozymes M2 (PKM2)
Overexpression of PKM2 in the aerobic glycolysis was associated with nodal metastasis [9, 11]. A recent research by Lee et al. has found that 14-3-3/C-myc as a new regulatory axis of cancer metabolism in breast cancer, including regulation of PKM2 [24]. We found that 14-3-3 knockdown has no effect on C-myc but can regulate pyruvate kinase PKM2 (Fig. 7B). We suggest that PKM2 and glycolysis might be part of the mechanisms of 14-3-3 in regulating the metastasis of ICC cells.
Discussion
Cholangiocarcinoma (CCA) is a hepatobiliary cancer with features of cholangiocyte differentiation. It can be classified anatomically as intrahepatic cholangiocarcinoma (ICC), hilar cholangiocarcinoma and extrahepatic cholangiocarcinoma (ECC) [26]. These subtypes differ not only in their anatomic location but in epidemiology, origin, etiology, pathogenesis and treatment [26]. ICC is a rare hepatobiliary malignancy that arises from the epithelial cells of the intrahepatic bile ducts. Epidemiologic data suggest that whereas ECC has remained stable or slightly declined, ICC incidence has increased significantly in recent years [14]. So a better understanding of the molecular mechanisms in the progression of ICC will improve the prognosis of ICC patients.
14-3-3 has been studied in many primary tumors and tumor cell line [16, 27]. However, there are few reports on the relationship between 14-3- function and ICC. In our research, we used two human ICC cell lines (low grade malignant cells RBE and high grade malignant cells HCCC-9810). For migration and invasion ability, there is moderate difference between the two cell lines (the migration ability of HCCC-9810 is slightly stronger than RBE). For anoikis resistance ability, HCCC-9810 is more resistant to anoikis than RBE. The reasons for this are complex and we have found that part of explanation for high tolerance to anoikis of HCCC-9810 seemingly lies in the higher expression of 14-3- in HCCC-9810. Further understanding of the difference needs to be further investigated. The findings of this study are, first, that tumor tissue studies demonstrate that 14-3- is highly expressed in ICC tumor tissues, compared with the adjacent tissues, as well as in normal liver tissue. Second, 14-3-3 plays an important role i invasion of ICC cell lines and regulates the invasion of ICC through regulating MMP2 and MMP9 expression and anoikis resistance. Consistent with our results, a previous study has demonstrated that overexpression of 14-3-3 was significantly associated with poor outcome in 78 patients with intrahepatic mass-forming cholangiocarcinoma after partial hepatectomy [39]. In conclusion, 14-3-3 is associated with the invasiveness of ICC and may be a novel therapeutic approach against ICC.
We suggest that 14-3-3 may play a role as double-edged sword in tumorigenesis [19], which may be due to its unique structure [2] and its property as a molecular chaperone by binding to various protein ligands important to many cellular processes. 14-3-3 is a direct target of p53 and protects p53 from MDM2-mediated ubiquitination and degradation. By binding to CDC2 and activating Chk1, 14-3-3also induces G2 arrest in response to DNA damages. Therefore, the loss or absence of 14-3-3 expression is currently considered an early event during carcinogenesis in many types of cancers. Also by binding to Bad or Bax, 14-3-3 prevents DNA damage-induced apoptosis [28, 34]. Upregulation of 14-3- was associated with reduced cell deat upon DNA damage [3]. DNA damage triggers p53 accumulation and transactivation of p53 target genes Wig-1. Levels of 14-3- mRNA are further induced by Wig-1 [3].
Our results show that 14-3-3 protein plays a crucial role in invasion of ICC cells. Similarly, the comparison of metastatic lymph node sample (LNs) with histologically negative lymph node sample (LNs) confirmed that LNs() express significantly more transcripts of 14-3-3 in vulvar carcinoma patients [16]. Anoikis is programmed death of cells triggered by detachment from a basement membrane or extracellular matrix [29]. The ability of the cell to survive in an anchorage-independent manner is necessary for tumor metastasis and aggressiveness [4, 32]. We also found 14-3-3 knockdown enhances anoikis of ICC cells. However, more mechanistic studies are needed to elucidate the role of 14-3-3 in anoikis resistance and invasion.
A previous study demonstrated that 14-3-3 appears to be important for supprssing the cisplatin resistance of gastric cancer cells [15] and the standard practice of care for advanced-stage ICC is systemic chemotherapy with cisplatin and gemcitabine [26, 31, 35]. Therefore, we will exam whether 14-3-3 downregulation can reduce ICC cells cisplatin resistance.
In summary, our study investigates the prognostic value of 14-3-. We are the first to investigate the role of 14-3-3 in metastasis of ICC patients, and to investigate its mechanism, which contributes to ICC cells migration, invasion and anoikis resistance. The limitations of our study are listed as follows. Firstly, the lacking of patients is unfavorable to the effective statistical analysis and our clinical study is retrospective. Secondly, since HCCC-9810 and RBE cells could not form tumors in nude mice and non-SCID mice (5 10 cells were injected subcutaneously into mice and the mice were sacrificed 8 weeks). Therefore, metastasis assay via tail intravenous injection or orthotopic implantation are impossible. This is largely due to a lack of different cell lines available.
Conclusion
High expression of 14-3-3 in ICC is associated with shorter overall survival and the invasiveness of ICC. It may be not only a potential prognostic marker, but also a possible therapeutic approach against ICC.
Footnotes
Acknowledgments
This study was sponsored by grants from Science and Technology Department of Zhejiang Province (No. 2015C03034), Major projects of Zhejiang Province in 12th Five-Year (NO.2013T301-15), National Health and Family Planning Commission of China (No. 2016 138643), National S&T Major Project (No. 2012ZX10 002017) and Innovative Research Groups of National Natural Science Foundation of China (No. 81421062).
References
1.
AitkenA., Post-translational modification of 14-3-3 isoforms and regulation of cellular function, Seminars in Cell & Developmental Biology22 (2011), 673–680.
2.
BenzingerA.PopowiczG.M.JoyJ.K.MajumdarS.HolakT.A. and HermekingH., The crystal structure of the non-liganded 14-3-3sigma protein: insights into determinants of isoform specific ligand binding and dimerization, Cell Res15 (2005), 219–227.
3.
BersaniC.XuL.D.VilborgA.LuiW.O. and WimanK.G., Wig-1 regulates cell cycle arrest and cell death through the p53 targets FAS and 14-3-3sigma, Oncogene33 (2014), 4407–4417.
4.
BuchheitC.L.WeigelK.J. and SchaferZ.T., Cancer cell survival during detachment from the ECM: multiple barriers to tumour progression, Nat Rev Cancer14 (2014), 632–641.
5.
ChoiH.H.GullyC.SuC.H.Velazquez-TorresG.ChouP.C.TsengC.ZhaoR.PhanL.ShaikenT.ChenJ.YeungS.C. and LeeM.H., COP9 signalosome subunit 6 stabilizes COP1, which functions as an E3 ubiquitin ligase for 14-3-3[sigma], Oncogene30 (2011), 4791–4801.
6.
CianfaraniF.BernardiniS.De LucaN.DellambraE.TatangeloL.TiveronC.NiessenC.M.ZambrunoG.CastigliaD. and OdorisioT., Impaired Keratinocyte Proliferative and Clonogenic Potential in Transgenic Mice Overexpressing 14-3-3[sigma] in the Epidermis, J Invest Dermatol131 (2011), 1821–1829.
7.
DowlingP.HughesD.J.LarkinA.M.MeillerJ.HenryM.MeleadyP.LynchV.PardiniB.NaccaratiA.LevyM.VodickaP.NearyP. and ClynesM., Elevated levels of 14-3-3 proteins, serotonin, gamma enolase and pyruvate kinase identified in clinical samples from patients diagnosed with colorectal cancer, Clinica Chimica Acta441 (2015), 133–141.
8.
FreemanA.K. and MorrisonD.K., 14-3-3 Proteins: Diverse functions in cell proliferation and cancer progression, Seminars in Cell & Developmental Biology22 (2011), 681–687.
9.
GaoY.XuD.YuG. and LiangJ., Overexpression of metabolic markers HK1 and PKM2 contributes to lymphatic metastasis and adverse prognosis in Chinese gastric cancer, International Journal of Clinical and Experimental Pathology8 (2015), 9264–9271.
10.
GardinoA.K. and YaffeM.B., 14-3-3 proteins as signaling integration points for cell cycle control and apoptosis, Seminars in Cell & Developmental Biology22 (2011), 688–695.
11.
GiannoniE.TaddeiM.L.MorandiA.ComitoG.CalvaniM.BianchiniF.RichichiB.RaugeiG.WongN.TangD. and ChiarugiP., Targeting stromal-induced pyruvate kinase M2 nuclear translocation impairs oxphos and prostate cancer metastatic spread, Oncotarget6 (2015), 24061–24074.
12.
HammondN.L.HeadonD.J. and DixonM.J., The Cell Cycle Regulator Protein 14-3-3[sigma] Is Essential for Hair Follicle Integrity and Epidermal Homeostasis, J Invest Dermatol132 (2012), 1543–1553.
13.
IwataN.YamamotoH.SasakiS.ItohF.SuzukiH.KikuchiT.KanetoH.IkuS.OzekiI.KarinoY.SatohT.ToyotaJ.SatohM.EndoT. and ImaiK., Frequent hypermethylation of CpG islands and loss of expression of the 14-3-3 sigma gene in human hepatocellular carcinoma, Oncogene19 (2000), 5298–5302.
14.
KhanS.A.Taylor-RobinsonS.D.ToledanoM.B.BeckA.ElliottP. and ThomasH.C., Changing international trends in mortality rates for liver, biliary and pancreatic tumours, J Hepatol37 (2002), 806–813.
15.
KimI.K.ParkS.M.ChoH.J.BaekK.E.NamI.K.ParkS.H.RyuK.J.RyuJ.ChoiJ.HongS.C.KimJ.W.LeeC.W.KangS.S. and YooJ., 14-3-3sigma attenuates RhoGDI2-induced cisplatin resistance through activation of Erk and p38 in gastric cancer cells, Oncotarget4 (2013), 2045–2056.
16.
KowalewskaM.RadziszewskiJ.GorycaK.BujkoM.Oczko-WojciechowskaM.JarzabM.SiedleckiJ.A. and BidzinskiM., Estimation of groin recurrence risk in patients with squamous cell vulvar carcinoma by the assessment of marker gene expression in the lymph nodes, BMC cancer12 (2012), 223.
17.
LeeM.H. and LozanoG., Regulation of the p53-MDM2 pathway by 14-3-3 sigma and other proteins, Seminars in Cancer Biology16 (2006), 225–234.
18.
LiD.J.DengG.XiaoZ.Q.YaoH.X.LiC.PengF.LiM.Y.ZhangP.F.ChenY.H. and ChenZ.C., Identificating 14-3-3 sigma as a lymph node metastasis-related protein in human lung squamous carcinoma, Cancer Letters279 (2009), 65–73.
19.
LiZ.LiuJ.Y. and ZhangJ.T., 14-3-3sigma, the double-edged sword of human cancers, American Journal of Translational Research1 (2009), 326–340.
20.
LiZ.DongZ.MyerD.Yip-SchneiderM.LiuJ.CuiP.SchmidtC.M. and ZhangJ.T., Role of 14-3-3sigma in poor prognosis and in radiation and drug resistance of human pancreatic cancers, BMC Cancer10 (2010), 598.
21.
LiuC.C.JanY.J.KoB.S.WuY.M.LiangS.M.ChenS.C.LeeY.M.LiuT.A.ChangT.C.WangJ.ShyueS.K.SungL.Y. and LiouJ.Y., 14-3-3sigma induces heat shock protein 70 expression in hepatocellular carcinoma, BMC Cancer14 (2014), 425.
22.
MoeiniA.SiaD.BardeesyN.MazzaferroV. and LlovetJ.M., Molecular Pathogenesis and Targeted Therapies for Intrahepatic Cholangiocarcinoma, Clinical Cancer Research22 (2016), 291–300.
23.
OhS.ShinS.LightfootS.A. and JanknechtR., 14-3-3 proteins modulate the ETS transcription factor ETV1 in prostate cancer, Cancer Research73 (2013), 5110–5119.
24.
PhanL.ChouP.-C.Velazquez-TorresG.SamudioI.ParrenoK.HuangY.TsengC.VuT.GullyC.SuC.-H.WangE.ChenJ.ChoiH.-H.Fuentes-MatteiE.ShinJ.-H.ShiangC.GrabinerB.BlonskaM.SkerlS.ShaoY.CodyD.DelacerdaJ.KingsleyC.WebbD.CarlockC.ZhouZ.HsiehY.-C.LeeJ.ElliottA.RamirezM.BanksonJ.HazleJ.WangY.LiL.WengS.RizkN.WenY.Y.LinX.WangH.WangH.ZhangA.XiaX.WuY.HabraM.YangW.PusztaiL.YeungS.-C. and LeeM.-H., The cell cycle regulator 14-3-3[sigma] opposes and reverses cancer metabolic reprogramming, Nat Commun6 (2015).
25.
RamirezJ.L., 14-3-3 Methylation in Pretreatment Serum Circulating DNA of Cisplatin-Plus-Gemcitabine-Treated Advanced Non-Small-Cell Lung Cancer Patients Predicts Survival: The Spanish Lung Cancer Group, Journal of Clinical Oncology23 (2005), 9105–9112.
26.
RizviS. and GoresG.J., Pathogenesis, diagnosis, and management of cholangiocarcinoma, Gastroenterology145 (2013), 1215–1229.
27.
SambandamS.A.KasettiR.B.XueL.DeanD.C.LuQ. and LiQ., 14-3-3sigma regulates keratinocyte proliferation and differentiation by modulating Yap1 cellular localization, J Invest Dermatol135 (2015), 1621–1628.
28.
SamuelT.WeberH.O.RauchP.VerdoodtB.EppelJ.T.McSheaA.HermekingH. and FunkJ.O., The G2/M regulator 14-3-3sigma prevents apoptosis through sequestration of Bax, The Journal of Biological Chemistry276 (2001), 45201–45206.
29.
SchaferZ.T.GrassianA.R.SongL.JiangZ.Gerhart-HinesZ.IrieH.Y.GaoS.PuigserverP. and BruggeJ.S., Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment, Nature461 (2009), 109–113.
30.
Shiba-IshiiA.KimY.ShiozawaT.IyamaS.SatomiK.KanoJ.SakashitaS.MorishitaY. and NoguchiM., Stratifin accelerates progression of lung adenocarcinoma at an early stage, Mol Cancer14 (2015), 142.
31.
SiaD.TovarV.MoeiniA. and LlovetJ.M., Intrahepatic cholangiocarcinoma: pathogenesis and rationale for molecular therapies, Oncogene32 (2013), 4861–4870.
32.
SimpsonC.D.AnyiweK. and SchimmerA.D., Anoikis resistance and tumor metastasis, Cancer Letters272 (2008), 177–185.
33.
SuC.H.ZhaoR.ZhangF.QuC.ChenB.FengY.H.PhanL.ChenJ.WangH.WangH.YeungS.C. and LeeM.H., 14-3-3sigma exerts tumor-suppressor activity mediated by regulation of COP1 stability, Cancer Research71 (2011), 884–894.
34.
SubramanianR.R.MastersS.C.ZhangH. and FuH., Functional conservation of 14-3-3 isoforms in inhibiting bad-induced apoptosis, Experimental Cell Research271 (2001), 142–151.
35.
ValleJ.WasanH.PalmerD.H.CunninghamD.AnthoneyA.MaraveyasA.MadhusudanS.IvesonT.HughesS.PereiraS.P.RoughtonM. and BridgewaterJ., Cisplatin plus Gemcitabine versus Gemcitabine for Biliary Tract Cancer, New England Journal of Medicine362 (2010), 1273–1281.
36.
WuY.J.JanY.J.KoB.S.LiangS.M. and LiouJ.Y., Involvement of 14-3-3 Proteins in Regulating Tumor Progression of Hepatocellular Carcinoma, Cancers7 (2015), 1022–1036.
37.
YangH.ZhaoR. and LeeM.H., 14-3-3sigma, a p53 regulator, suppresses tumor growth of nasopharyngeal carcinoma, Molecular Cancer Therapeutics5 (2006), 253–260.
38.
YangH.Y.WenY.Y.ChenC.H.LozanoG. and LeeM.H., 14-3-3 sigma positively regulates p53 and suppresses tumor growth, Mol Cell Biol23 (2003), 7096–7107.
39.
YehC.N.PangS.T.JungS.M.ChenT.W.JanY.Y. and ChenM.F., cCytoplasmic overexpression with membrane accentuation of stratifin in intrahepatic mass-forming cholangiocarcinoma after hepatectomy: correlation with clinicopathologic features and patient survival, Journal of Surgical Oncology102 (2010), 608–614.
40.
ZhangH.YangT.WuM.C. and ShenF., Intrahepatic cholangiocarcinoma: Epidemiology, risk factors, diagnosis and surgical management, Cancer Letters379 (2016), 198–205.
41.
ZhangY.LiY.LinC.DingJ.LiaoG. and TangB., Aberrant Upregulation of 14-3-3σ and EZH2 Expression Serves as an Inferior Prognostic Biomarker for Hepatocellular Carcinoma, PLoS ONE9 (2014).
42.
ZhaoJ.MeyerkordC.L.DuY.KhuriF.R. and FuH., 14-3-3 proteins as potential therapeutic targets, Seminars in Cell & Developmental Biology22 (2011), 705–712.
43.
ZhengG.XiongY.YiS.ZhangW.PengB.ZhangQ. and HeZ., 14-3-3sigma regulation by p53 mediates a chemotherapy response to 5-fluorouracil in MCF-7 breast cancer cells via Akt inactivation, FEBS Lett586 (2012), 163–168.