
Abstract
Background:
Bladder cancer (BC) cell lines are indispensable in basic and preclinical research. Currently, an up-to-date and comprehensive overview of available BC cell lines is not available.
Objective:
To provide an overview and resources on the origin, pathological and molecular characteristics of commonly used human, murine and canine BC cell lines.
Methods:
A PubMed search was performed for relevant articles published between 1980 and 2017 according to the following MeSH terms: cell line; cell line, tumor; urinary bladder neoplasms; carcinoma, transitional cell. The Cellosaurus database was searched, using the term “bladder” and/or “urothelial carcinoma”. We followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses guidelines.
Results:
We provide information on 157 human, murine and canine BC cell lines. 103 human BC cell lines have molecular data available, of which 69 have been profiled by at least one “omic” technology. We outline how these cell lines are currently being used for in vitro and in vivo experimental models. These results allow direct comparison of BC cell lines to patient samples, providing information needed to make informed decisions on the most genomically appropriate cell line to answer research questions. Furthermore, we show that cross-contamination remains an issue and describe guidelines for prevention.
Conclusions:
In the BC field, multiple human, murine and canine BC cell lines have been developed and many have become indispensable for in vitro and in vivo research. High-throughput -omic technologies have dramatically increased the amount of molecular data on these cell lines. We synthesized a comprehensive overview of these data as a resource for the BC scientific community.
INTRODUCTION
In 2017, an estimated 17.000 deaths are caused by bladder cancer (BC) in the United States. Worldwide, BC is the ninth most common cancer, with a prevalence of more than 1.300.000 patients [1, 2]. Urothelial carcinoma is the predominant histological subtype with 70–80% being non-muscle invasive disease (NMIBC) at diagnosis. NMIBC patients have a high 5-year overall survival, but tumors frequently recur and ∼30% of these patients will have progression to muscle invasive bladder cancer (MIBC), which is associated with a high morbidity and mortality [3, 4]. Important goals for BC research include the discovery of (urine) biomarkersfor non-invasive surveillance of NMIBC patients, biomarkers to predict response to treatment or progression and development of novel therapeutics for both NMIBC and MIBC.
To address these goals, human tissue and cell lines have been the primary tools used in preclinical studies [5]. BC cell lines reflect some of the genetic aberrations found in human BC and thus provide the opportunity to investigate basic tumor biology and preclinical pharmacological responses [6–11]. For example, urothelial cell lines containing Fibroblast Growth Factor Receptor-3 (FGFR3) mutations and cell lines with FGFR3-transforming acid coiled coil 3 (FGFR3-TACC3) fusions, which are highly sensitive to FGFR-selective agents, have been used to evaluate the efficacy of these new targeted treatments and are the foundation of currently running clinical trials [NCT01004224, NCT01732107, NCT02278978 and NCT02529553] [12, 13]. Furthermore, the COXEN (co-expression extrapolation) bioinformatics algorithm was used to generate multigene predictive models for various malignancies by using in vitro chemotherapy and radiation sensitivity data of transcriptionally profiled cancer cell lines [14]. This work also led to the identification of novel drugs for BC as well as potential drug targets that could enhance radiosensitivity of cancercells [15, 16].
In recent years, large-scale cell line characterization by major research consortia has increased significantly [17–19]. For example, project “Achilles” is aimed at identifying and cataloguing genes that are essential for cell survival [20]. In this project, genome-scale RNAi and CRISPR-Cas9 knockout is used in multiple human cancer cell lines to silence individual genes and data is available online as a resource. The Cancer Cell Line Encyclopedia (CCLE) has so far catalogued 1457 cell lines and performed multi-omic profiling of the majority of these lines [17]. Genomic characterization has led to opportunities for new in vitro and in vivo studies [21, 22]. Combined, these models offer different and, in many cases, complementary benefits. To fully exploit their potential, detailed information on cell line characteristics and preclinical models is essential for experimental design by every BC researcher. At the same time, concern about adequate selection, cross-contamination and verification of cell lines has not yet been resolved [23]. Here, we present a systematic review on the origin and molecular characteristics of currently available human, murine and canine BC cell lines for preclinical BC research. We discuss in vitro and in vivo BC models using these cell lines, together with their advantages, shortcomings and points of interest.
EVIDENCE ACQUISITION
A literature search was conducted using the PubMed, Medline, Embase, Web of Science OvidSP, Google Scholar and Cellosaurus databases for all published results on BC cell lines from 1980 to 2017. The PubMed archive was searched for BC cell lines (November 2017) and the search strategy was as follows: (cell line[MeSH Major Topic] OR cell line, tumor[MeSH Terms]) AND (Urinary Bladder Neoplasms[MeSH Terms] OR Carcinoma, Transitional Cell[MeSH Terms]). The search resulted in 3147 English language, full text articles. BC cell lines were included if they were published at least twice since 1980, of which at least one publication since 2000. Incidentally found urothelial cancer cell lines of ureter or renal pelvis and a single urachal adenocarcinoma cell line were included as well. Contaminated BC cell lines, genetically engineered BC cell lines and patient-derived xenografts (PDX) were excluded. A search on Cellosaurus for ‘bladder’ and ‘urothelial’ provided 387 and 137 hits respectively (December 2017). After excluding duplicates, 419 remained. All cell lines were investigated and eligible cell lines were verified on PubMed and/or Google Scholar and added to the search results. Eventually, a total of 157 human, murine and canine bladder cancer cell lines were included in this systematicreview (Fig. 1).
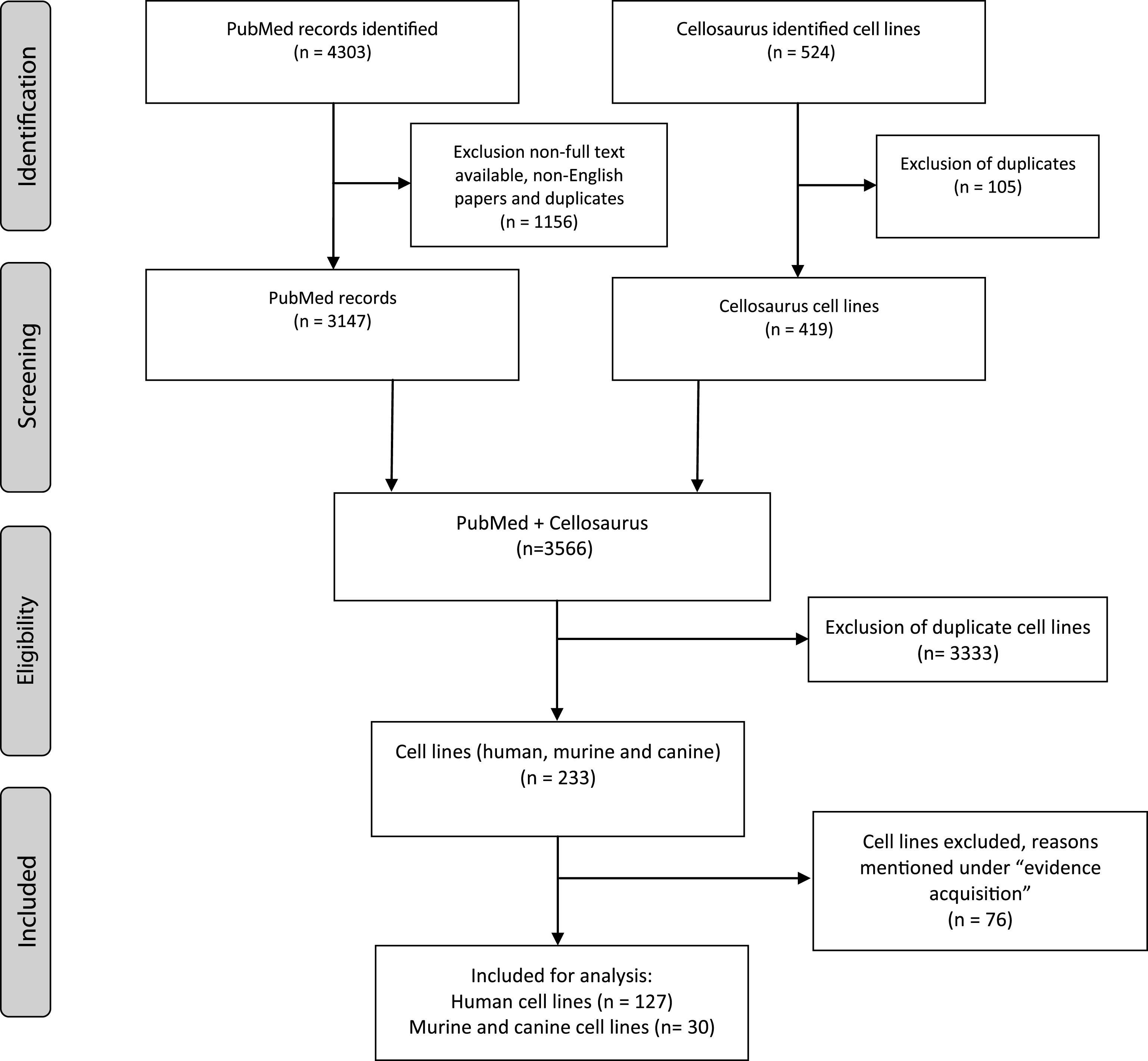
Summary of evidence search and selection. Selection process for studies to be included is in compliance with PRISMA.
Historical cell line data like origin, gender, development, stage and grade were collected for each cell line. Thereafter, databases of the Catalogue of Somatic Mutations in Cancer (COSMIC), CCLE, the International Agency for Research on Cancer (IARC) TP53, Earl et al. (2015) and Nickerson et al. (2017) were used to report on the molecular characterization of the reviewed cell lines [17–19, 24]. These large databases incorporate molecular data from multiple international research projects working with cell lines, while in the past characterization was limited to low-throughput techniques [25, 26]. An overview of widely-used online databases for BC cell line research is reported inTable 1.
Overview of available online resources for bladder cancer cell lines
IN VITRO MODELS: CELL LINE ORIGIN, DERIVATION AND MOLECULAR CHARACTERISTICS
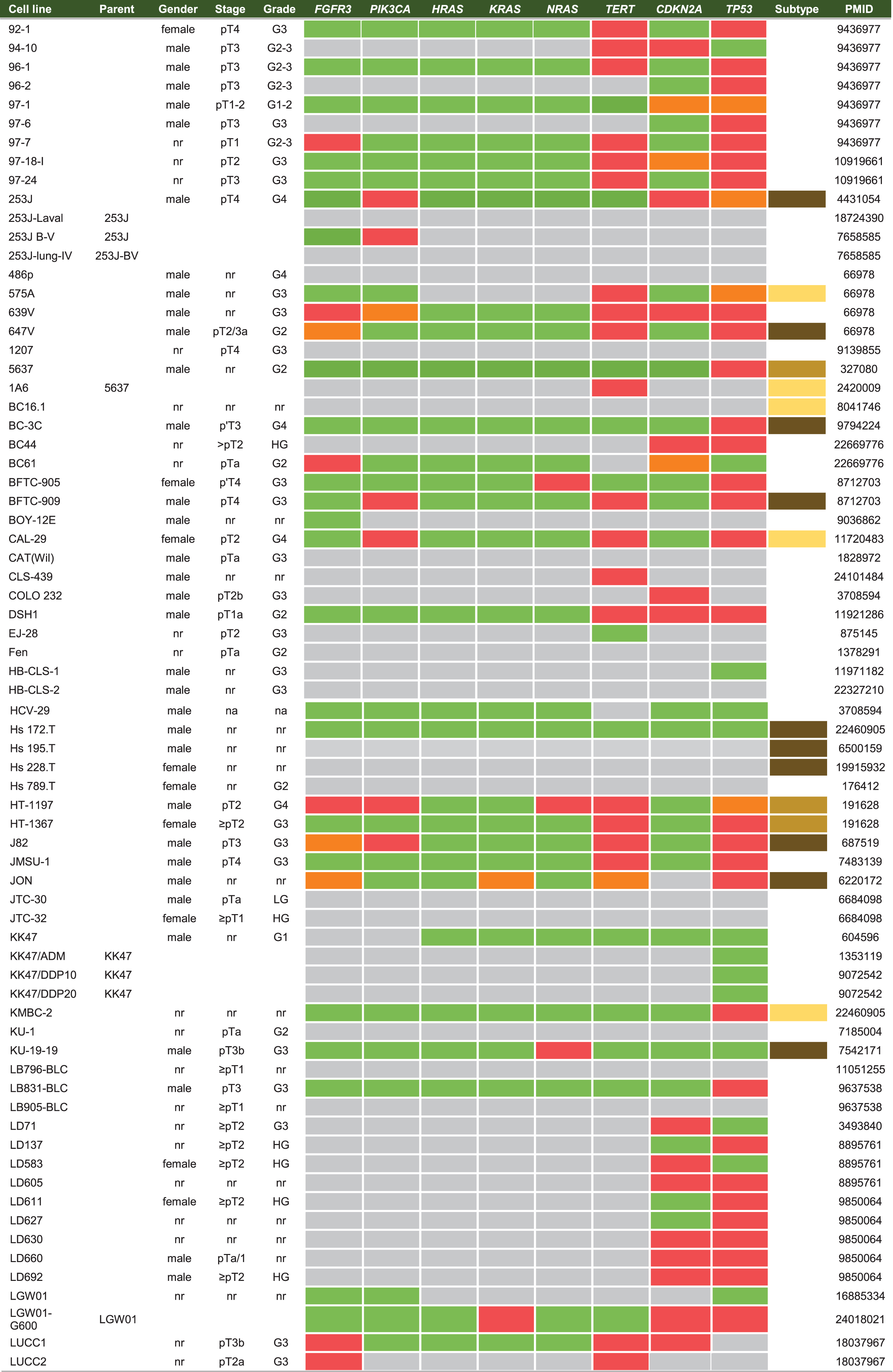
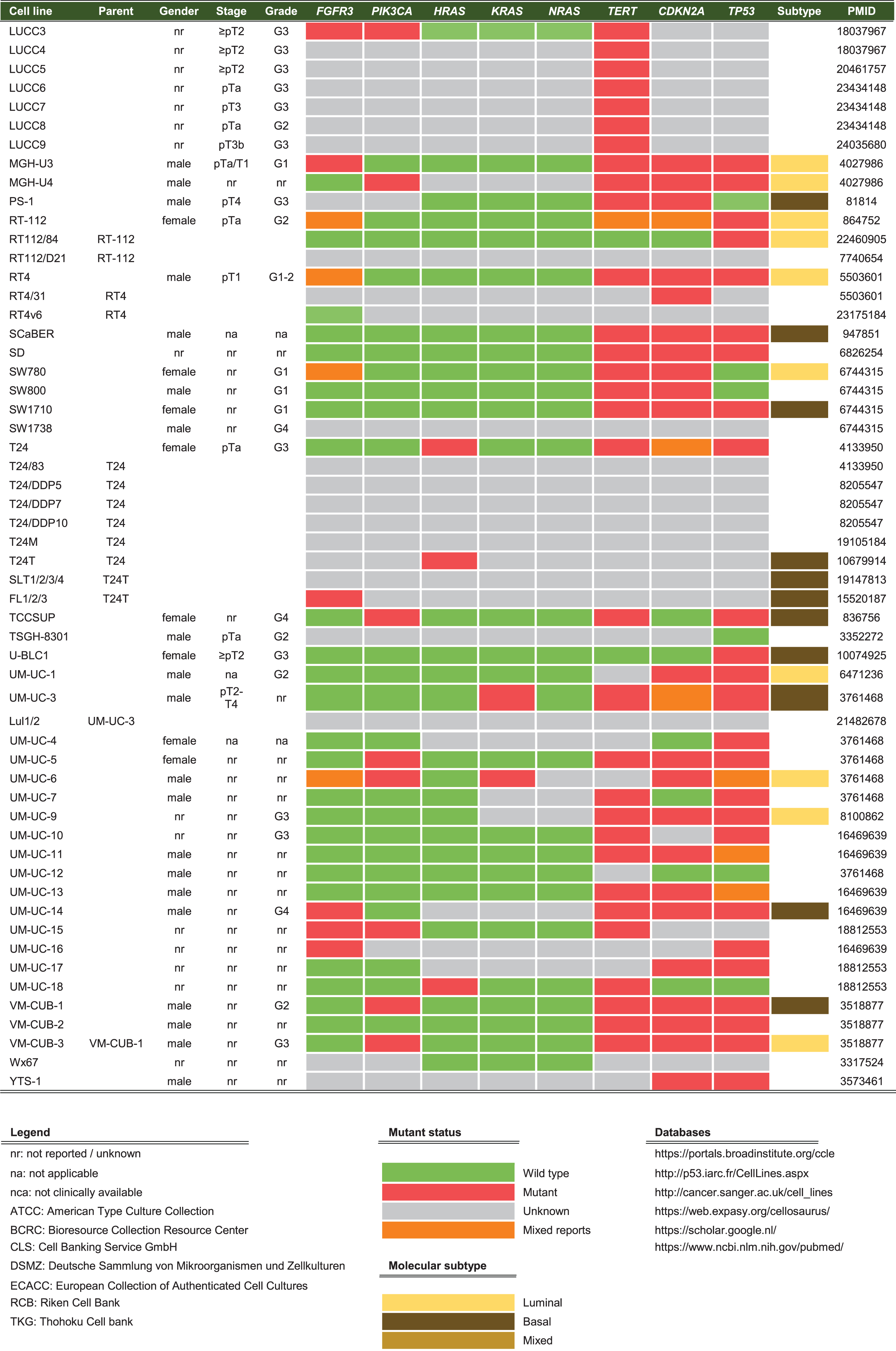
Cell lines are produced by either direct in vitro culture of patient tumor cells or by implantation of a patient’s tumor cells in immunodeficient mice and passaging the tumor multiple times before taking cells into culture. Currently, approximately 103 human BC cell lines of different stages and grades have molecular data available, of which 69 have been profiled by at least one – omic technology [18]. In general, molecular characterization of BC cell lines revealed that most genetic aberrations found in BC cell lines resemble those of the original tumor [19, 27–29, 19, 27–29]. Condensed historical and molecular data of the reviewed human BC cell lines is summarized in Table 2 (Complete information available in Supplementary Table 1). Many cell lines are now part of panels that have been characterized by different research groups. For example, Genomics of Drug Sensitivity in Cancer (GDSC) determined molecular alterations by using 20 BC cell lines to predict response to anti-cancer therapeutics [30]. Similarly, The Cancer Proteome Atlas (TCPA), included 11 BC cell lines in their effort to measure expression of cancer related proteins and how this may impact drug sensitivity [31]. More recently, whole exome sequencing on a subset of the UBC-40 panel (UC-25) confirmed the detection of mutations in TP53, TERT, PIK3CA and ERBB2 and other cancer-associated genes, in line with earlier TCGA findings in patients [18, 32]. Also, 12 cell lines had homozygous deletions of chromosome 9p21, resulting in significant loss of expression of MTAP, CDKN2A and CDKN2B, concordant with previous findings [19, 33]. Because cell lines now overlap in different panels, we provide a full report of which cell lines were profiled in each panel in Table 2 and Fig. 2. In total, 69 of 127 human BC cell lines are represented in at least one of the five different panels. The most frequently characterized cell lines that have been profiled in all five panels are HT-1197, T24, and TCCSUP (Fig. 2). Table 2 shows that a relatively small number of human BC cell lines are of low stage (12/127) and low-grade (8/127). This is mainly caused by the difficulty of culturing low-grade tumors in vitro and the clinical need for late-stage and high-grade disease models. However, NMIBC are the bulk of all BC tumors worldwide and the high costs associated with bladder cancer mainly originates from these patients [2]. Thus, development of low-grade cell lines should not be neglected by the BC community. Finally, molecular subtyping of BC has shown promising results to predict clinical outcomes in BC patients [34]. Luminal and basal molecular subtypes have also been demonstrated in BC cell lines [11, 35]. Our data shows that 39/127 human cell lines currently have been analysed; 15 luminal cell lines, 21 basal cell lines and 3 cell lines have a mixed report (Table 2 and Supplementary Table 2). Overall, data in Table 2 show that cell line mutational frequencies correspond to frequencies reported in in patients [32]. These findings could be useful in selecting the optimal cell line for specific research questions.
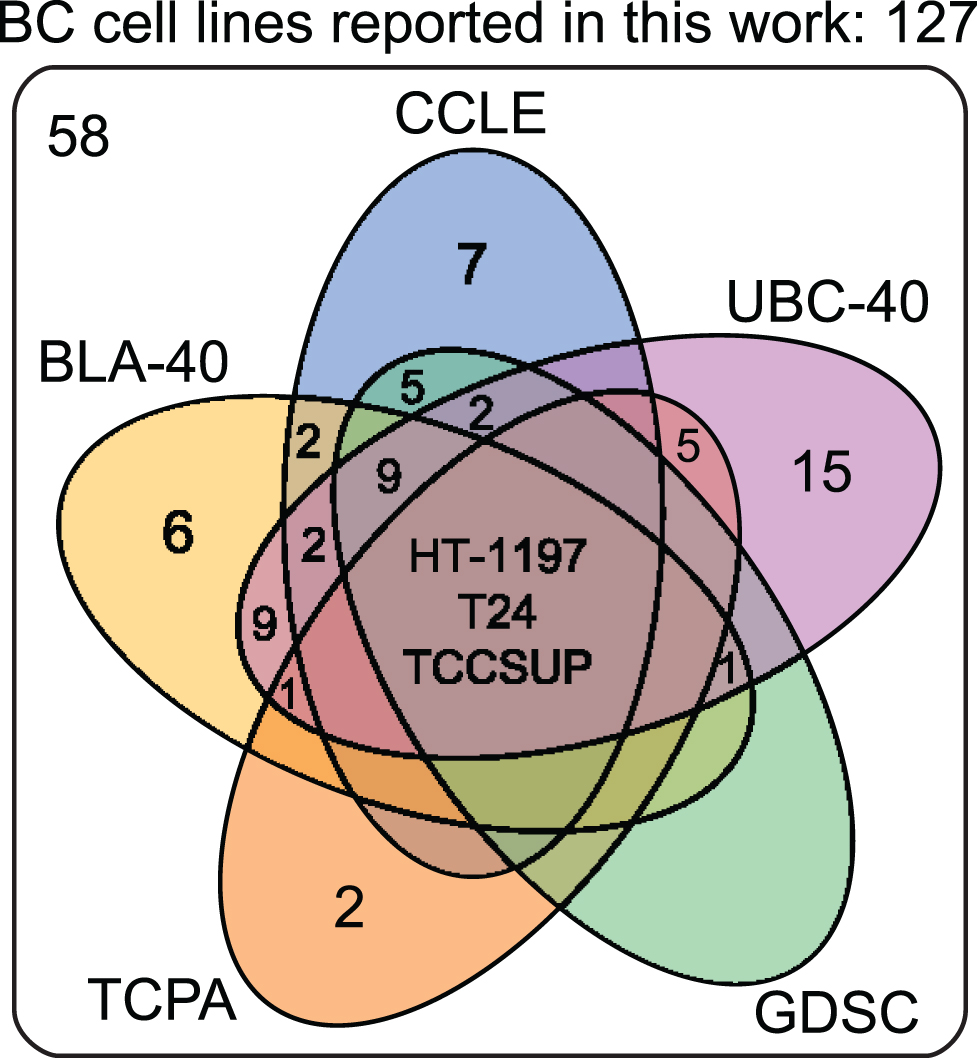
Venn diagram of human bladder cancer cell lines analyzed in five panels. 127 human BC cell lines are present in five panels. The overlap between different molecular panels is visualized. Only HT-1197, T24 and TCCSUP have been profiled in all five panels. CCLE: The Cancer Cell Line Encyclopedia; UBC-40: Urothelial Bladder Cancer-40; GDSC: Genomics of Drug Sensitivity in Cancer; TCPA: The Cancer Proteome Atlas; BLA-40:Bladder-40.
Summary of human BC cell lines (see Supplementary Table 2 for complete overview)
MURINE AND CANINE BLADDER CANCER CELL LINES
As in vivo use of human cell lines requires immunodeficient models, multiple BC cell lines from different species have been developed to study aspects of tumor immunity in vivo. Studies in the 1960s first explored chemically induced BC in mice by exposure to different carcinogens [36]. After exposure, most mice developed poorly differentiated tumors, which were heterogeneous and resembled human MIBC [37]. For instance, murine BC can be induced by exposure to carcinogenic chemicals like, 7, 12-dimethylenzanthacene (DMBA), N-methyl-N-nitrosourea (MNU), (N-butyl-N-(4-hydroxybutyl) nitrosamine (BBN), or N-[4-(5-nitro-2-furyl)-2thiazolyl]-formamide (FANFT) [36]. DMBA is a mutagen that causes DNA damage after activation. MNU is a genotoxic compound, which is administered intravesically and acts directly on urothelium with tumor formation after ∼8 weeks. BBN is administered orally to mice, with tumors developing after a ∼16-week diet of 0.05% BBN in drinking water [38–40]. Like BBN, FANFT can also be administered orally, resulting in carcinomas of both urothelial and squamous cell origin after ∼8 months [41]. After tumor formation in the bladder, tumor cells are harvested, cultured and passaged in vitro to develop a novel murine BC cell line. Currently, multiple carcinogen-induced BC cell lines are available. However, two murine BC cell lines are predominantly used in BC research: MB49 and MBT2 [42, 43]. MB49 was developed by orthotopic transplantation of mouse bladder urothelial cells, which were transformed by exposure to DMBA for 24 hours [42]. Although a male mouse was the original donor, karyotypic exploration showed MB49 has lost the Y-chromosome after in vitro culture [44]. MBT2 was produced by serial transplantation of a FANFT-induced tumor in mice [43]. Recently, Fantini et al. reported that BBN tumors had an overexpression of markers of the MIBC basal subtype [45]. Moreover, BBN tumors showed a high mutational burden similar to MIBC patients. Interestingly, the predominantly used MB49 (DMBA) and MBT2 (FANFT) cell lines in research showed important differences in their mutational profiles compared to human and BBN-induced tumors [45]. These findings demonstrate the importance of molecular profiling to select the appropriate research model and to ultimately improve translational research. BTT-T739 is another murine cell line that causes multiple visceral metastases, but is less frequently used in practice [46]. A fourth murine line is AY-27, generated after feeding FANFT to Fisher 344 rats. AY-27 causes in vivo high-grade NMIBC with progression to MIBC after ∼6 days [47]. A condensed overview of all murine cell lines is given in Table 3 (complete information available in Supplementary Table 3).
Summary of murine & canine BC cell lines (see Supplementary Table 3 for complete overview)
Legend: nr = not reported/unknown. Databases: https://portals.broadinstitute.org/ccle; http://p53.iarc.fr/CellLines.aspx; http://cancer.sanger.ac.uk/cell_lines; https://web.expasy.org/cellosaurus/; https://scholar.google.nl/; https://www.ncbi.nlm.nih.gov/pubmed/.
Canine cell lines are increasingly available and used in BC research. In dogs, BC is a naturally occurring disease, with some breeds (Scottish Terriers) even having a life-time risk of BC than other breeds [48]. Unfortunately, BC is typically diagnosed when the tumor is already muscle-invasive or metastasized, leaving most dogs incurable [49]. Therefore, canine BC offers a unique opportunity to study advanced BC and as a result characterization of canine BC has increased in the last years [50]. Invasive BC in dogs is similar to human MIBC both histologically and molecularly and therefore provides us with a potential BC model [51]. Knapp et al. generated one of the first canine lines (K9TCC) and thereafter more canine BC cell lines followed [52, 53]. Recently, extensive molecular characterization of a canine tumor cell line panel was done by the Flint Animal Cancer Center (FACC) [54]. FACC data will prove a valuable resource for BC research. An overview of our search for commonly used canine BC cell lines is presented in Table 3. Of note is the lack of high-throughput molecular characterization in both murine and canine cell lines. As these cell lines are frequently used, it would be of great benefit to the research community to expand the molecular characterization of murine and canine cell lines so the correct model can be chosen accordingly.
BLADDER CANCER CELL LINES FOR IN VIVO RESEARCH
The use of immunocompromised mouse models for human xenografts limits investigation of interactions between the tumor and its host. In vivo immunocompetent mouse tumor models have the advantage of an intact host immune system and tumor microenvironment (TME) and thus provide a platform to study tumor-host interactions and immunotherapeutics [6, 56]. In vivo models can be divided in two main groups: 1) spontaneous or autochthonous tumor models (subdivided in carcinogen-induced and genetically engineered/transgenic models) and 2) engraftment or non-autochthonous models (subdivided in allograft/syngeneic and xenograft/xenogeneic models). In contrast to tumor heterogeneity of carcinogen-induced models, BC in genetically engineered mouse models (GEM) originates due to fixed experimentally applied genetic changes found to be associated with BC and complement carcinogen-induced mouse models. Recently published TCGA data will be used to aid the development of novel GEMs [32]. More detailed information on GEMs and carcinogen-induced BC models has been published elsewhere [57–59]. Syngeneic models are generated by implantation of murine BC cell lines into immunocompetent mice and provide insight into the biology of TME interactions and are excellent tools to evaluate immunotherapy such as intravesical Bacillus Calmette-Guérin or checkpoint inhibitors [60–62]. Xenogeneic models are generated by implantation of human BC cell lines (xenografts) or PDXs into immunodeficient mice [63–65]. PDXs are molecularly more related to the original tumor, show greater intratumoral heterogeneity and histological similarity to the primary tumor, even after several passages in mice [65–67]. PDXs can therefore be used to directly evaluate a patient’s response to treatment [65–67]. Here, we only focus on immortalized, tumor-derived cell lines and reviews on PDX models have been published elsewhere [58, 68].
In vivo models can also be classified according to the site of tumor implantation; orthotopic (in the bladder) or heterotopic (elsewhere, not in the bladder) [58, 70]. Orthotopic models can either be spontaneous (i.e. orthotopic carcinogen-induced) or engrafted [71]. Damage to the bladder wall is generally used to increase orthotopic tumor engraftment [69], yet the “tumor take rate” varies considerably between studies [69, 73]. Multiple procedures exist to damage the bladder wall, for example transurethral mechanical injuring, intravesical exposure to corrosive chemicals, laparotomy or minimal invasive percutaneous surgery [73–76]. The main advantage of inoculating tumor cells inside the bladder, is a more realistic simulation of the TME found in BC patients. Challenges with orthotopic models include the requirement for imaging methods to measure tumor growth or procedures of sequential cohort euthanasia and necropsy [77]. Non-invasive assessment of tumors can be accomplished through magnetic resonance imaging, intravesical ultrasonography and in vivo fluorescence of tumors [61, 79]. Common heterotopic tumor inoculation sites include subcutaneous or subrenal capsule models [61, 65]. Subcutaneous models allow easy tumor size measurements but may have different responses to drugs and a different TME compared to the orthotopic location [80, 81]. Alternatively, subrenal capsule models have a higher tumor take than orthotopic engraftment models, probably due to elimination of orthotopic tumor cells by voiding or because subrenal capsules tumors benefit from the rich blood supply of the subrenalarea [82].
METASTATIC BLADDER CANCER MODELS
Patients with metastases have a poor clinical outcome and contribute to the bulk of mortality in BC. Experimental models are invaluable to investigate the complex multistep metastatic process that enables invasive growth of the primary tumor, detachment from the primary site, intravasation, migration, extravasation and colonization of distant organs. This multistep process is difficult to replicate in vivo, because mice often die before metastases develop. Moreover, kidney failure occurs frequently, as both engrafted and carcinogen-induced orthotopic tumors cause post-renal obstruction due to a large bladder tumor. Another limitation is the lack of available tissue from highly metastatic human tumors and thus limited availability of metastatic BC models. Therefore, researchers use in vivo serial passaging to acquire more aggressive cell lines. Serial passaging can be orthotopic, by portal vein injections, tail vein injections or intracardiac injections. The different routes determine the metastatic pattern of spreading [83]. For example, metastatic cell line Lul1/2 was produced from the UM-UC-3 parent cell line by tail vein injection, causing tumor formation in the lungs. Next, lung nodules were harvested and re-injected into the tail vein and the cycle was repeated multiple times, generating Lul1/2 cells with greater capacity to proliferate and aggressively expand in a metastatic site [84]. Two models from the parental cell line T24 have been developed by using the same approach and are currently available for studying lung and liver tropism in BC [85, 86]. Lung metastasis model T24T/FL1-3 was developed by repeated tail vein injections of T24T followed by harvesting of lung nodules to develop cell lines with increased capabilities to colonize distant organs [85]. T24T/SLT1-3 liver metastatic lines were developed by intrasplenic injections of T24T followed by harvesting of liver nodules and repeated intrasplenic injections [86]. These models allowed identification of molecular changes associated with the metastatic phenotype to liver and lung.
For syngeneic murine models, MBT-2 and MB49 cell lines are frequently used. MBT-2 has a low frequency of metastatic spreading to the lungs when injected intravenously, but metastatic potential increases when tumor cells are injected intramuscularly [87]. To improve the metastatic potential of MB49, cells were passaged multiple times subcutaneously, causing cells to acquire a more invasive phenotype (MB49-I). After orthotopic inoculation of MB49-I cells, mice developed multiple lung metastasis [88]. In summary, there are limited metastatic BC cell lines and models available. Expanding the number of metastatic BC lines and optimizing current metastatic models is an area that would benefit the BC research community.
CAVEATS OF CANCER CELL LINES
Despite their advantages and contributions to the field, researchers should be aware of cell line limitations. With each passage, cells experience genetic changes due to selective pressure, which may lead to a change in phenotype [89]. After multiple passages, BC cell lines showed loss of heterozygosity, microsatellite instability and changes in ploidy compared to the original tumor [90–92]. Genetic changes and selection are also induced by passaging cells for long time periods under suboptimal culture conditions (e.g. lack of nutrients or passaging cells at low density) [89, 93]. It is therefore important to be aware of how a cell line is developed (single clone or heterogeneous tumor cell culture), culture conditions and to realize that cell lines may represent a subset of cellular features of the original tumors [6, 94].
Cross-contamination
Another challenging aspect when using cancer cell lines is cross-contamination. Cross-contamination is the result of contamination of an original cell line with a contaminant cell line that has a dominant growth pattern, eventually outgrowing the original cell line. Cross-contamination was discovered as early as 1967, when Gartler et al. accidently found that a panel of cell lines of mixed gender all shared a common X-linked genetic polymorphism (G6PD allele) that was almost exclusive for people of African descent. Further analysis showed that his cell lines were likely contaminated with HeLa cells from donor Henrietta Lacks, who is of African descent [95, 96]. Unfortunately, the process of cross-contamination can be gradual and insidious, especially if visual morphology of the contaminating cells is similar to that of the host line. HeLa cells have often been associated with cross-contamination of other cultures, because HeLa cells grow rapidly and are widely used [97, 98]. Genetic fingerprinting of various BC cell lines suggests some are now HeLa cells [99, 100]. For example, cell line KU-7 was derived from a LG papillary tumor, yet acquired invasive growth characteristics after multiple passages [99]. KU-7 turned out to be contaminated with HeLa cells, likely even before 1984, thereby raising questions about several studies [99, 102]. Another example is T24, which has contaminated a variety of cell lines [103–106]. Recent estimates suggest that up to a third of all documented cell lines are contaminated and no longer represent the original tumor [107, 108]. Given the magnitude of the cross-contamination problem, the National Institute of Health (NIH) requires authentication of cell lines, for instance by using short tandem repeat profiling (STR-profiling) when cell lines are used in NIH funded research [23, 109–111]. Additionally, multiple tests are available to determine which species a cell line originated from, like isoenzymology and DNA-barcoding [112]. Last, consulting overviews of cross-contaminated cell lines before usage is strongly recommended. An overview of published cross-contaminated BC cell lines is given in Table 4 [113]. Verification of cell lines before manuscripts are accepted will hopefully reduce the frequency of cross-contamination and improve the quality of cell line-related research in the future[114–116].
Overview of documented cross-contaminated BC cell lines
Legend: GEM: Genetically Engineered Model; DSMZ: Deutsche Sammlung von Mikroorganismen und Zellkulturen; na: cells derived from parental cells found to be cross-contaminated. Database: http://iclac.org/wp-content/uploads/Cross-Contaminations-v7_2.pdf; https://web.expasy.org/cgi-bin/cellosaurus/search - ‘Problematic cell line bladder’.
CONCLUSIONS
Many cell lines have been developed for BC research, yet a relatively small selection of these lines is currently being used. Important considerations for cell line research and selection of preclinical models are; 1) each model has unique advantages and limitations to take into account; 2) cell line authentication is considered mandatory before use; 3) orthotopic inoculation more accurately represents clinical BC, however these models are more challenging in use;4) human BC cell lines implanted in immunodeficient mice cannot be used to explore the tumor microenvironment in vivo and 5) extensive genomic and molecular characterization of cell lines allow investigators to select lines with desired genetic characteristics. While there is no flawless model, cell lines are an indispensable part of ongoing research. Optimization of current models is needed, as well as development of new metastasis models, especially from highly metastatic tumors. Finally, current in vitro and in vivo models using cell lines can be used in conjunction with new GEMs and PDXs to create a wide range of modelling options for the BC research community.
AUTHOR CONTRIBUTIONS
Concept: Theodorescu
Figure 1: De Jong
Figure 2: Costello, De Jong
Search: De Jong, Zuiverloon
Tables: De Jong, Zuiverloon
Writing: Zuiverloon, De Jong, Costello, Theodorescu