
Abstract
The pleiotropic nature of the apolipoprotein E (APOE) gene is associated with complex diseases in different populations. We analyzed APOE polymorphisms in 76 individuals from Jujuy – Argentina using NGS technology. The observed genotypes align with the expected Hardy–Weinberg equilibrium. APOE3 was the most common allele, followed by APOE4 and APOE2. The allele distribution pattern is consistent with findings in previously studied populations of Native Americans and Asians. The E4 allele’s low frequency, always observed in a heterozygous state, raises questions regarding its relevance in explaining dementia and longevity associated with this marker in the Central Andes.
INTRODUCTION
The APOE gene is linked to lipoproteins of both very low and high density, as well as chylomicrons, and it plays a crucial role in lipid metabolism [1]. There are three major isoforms, namely E2, E3, and E4, which are identified through isoelectric focusing and coded for by three alleles (epsilon 2, 3, and 4). These isoforms vary in their amino acid sequence at site 2, residue 112 (site A), and residue 158 (site B). The APOE alleles have shown pleiotropic effects in various populations, being linked to a plethora of complex diseases and traits, including neurodegenerative disorders, proteinopathies, lipid metabolism, cardiovascular and cerebrovascular disease, and longevity [2–4]. Furthermore, recent studies indicate that individuals homozygous for APOE4 face an elevated risk of severe COVID-19 infection [5]. Notwithstanding, it is important to note that not all alleles have an equal likelihood of developing the described associations. There is evidence of a gradual increase in risk and loss of protein function from APOE2 to APOE3 and APOE4, with some controversy surrounding APOE2 having a protective effect. Furthermore, the probability of association increases in accordance with the number of APOE4 alleles present, ranging from 4 in heterozygotes to 12 in homozygotes [3].
APOE allele distribution is heterogeneous globally, with APOE3 being the most common allele across all populations, followed by APOE4. On a continental level, Africa and Oceania present the highest E2 and E4 frequencies, while E3 is the predominant allele in Asia [6, 7]. However, there is a scarcity of studies concerning APOE polymorphism distribution in indigenous populations in the Andes [8–10].
Jujuy’s population, located in the Andes, exhibits a high frequency of autochthonous surnames in their onomastics and an elevated Native American ancestry [12, 13]. This study analyses the distribution of APOE allele in an admixed population from Jujuy and compares it with other Native American groups from the Central Andes.
MATERIALS AND METHODS
In this study, exome data on 76 pediatric patients, unrelated to one another, was examined for genomic studies on various genetic diseases initially unrelated to APOE. The patients were invited by the Medical Genetics Unit of the Maternal and Child Public Hospital of Jujuy and each gave written consent to participate. The study has been approved by the Provincial Bioethics Committee.
Genomic DNA was extracted from whole blood (300μl) utilizing a QIAamp DNA Blood purification kit as per QIAGEN GmbH’s directions. Library preparation, exome, and neighboring intronic regions capture were conducted with Agilent’s SureSelect All Exon V6 kit. Full exome sequencing by NGS was performed using an Illumina NovaSeq PE150 platform. The protocol followed for mapping, alignment, and variant calling was based on the Best Practices of the Eli and Edythe L. Broad Institute of Harvard and MIT considering GRCh37 as the reference genome [14].
The identification of alleles in each patient was carried out by utilizing two APOE markers, rs429358 and rs7412. These markers determine the presence of E2 alleles (rs429358 T, rs7412 T), E3 alleles (rs429358 T, rs7412 C), and E4 alleles (rs429358 C, rs7412 C). Abbreviations were explained when first used. Genotypic and allelic frequencies were computed, and Hardy-Weinberg equilibrium was evaluated via Arlequin v3.5 software [15]. Genetic differentiation among samples was appraised utilizing an exact G test from Genepop v4 software [16].
To facilitate exploratory analysis, allelic frequencies from other Native American populations were included [9, 10], and the dataset was evaluated through cluster analysis employing Euclidean distances for the E2, E3 and E4 allelic frequencies with R software [17].
RESULTS
The recorded genotypes conform to those anticipated by the Hardy-Weinberg equilibrium. Our findings are presented in Table 1, where we collated them with comparable populations from the Central Andes. Despite conducting the exact G test, we did not identify any statistically significant difference. Nevertheless, increasing the sample size could reveal such differences. APOE3 dominated as the primary allele, with APOE4 and APOE2 manifesting in low frequencies. The APOE3 allele was present in a homozygosis state with a frequency of 0.89. Figure 1 presents cluster analysis results on APOE allele frequencies in various Native American populations, indicating frequencies measured in this study to be comparable to those found in other Andean populations. This study also notes that the lower frequency of the E2 and E4 alleles is not unique to the Andean population.
Genotypic and allelic frequencies of APOE3 in the population of Jujuy and its comparison with populations from the Central Andes
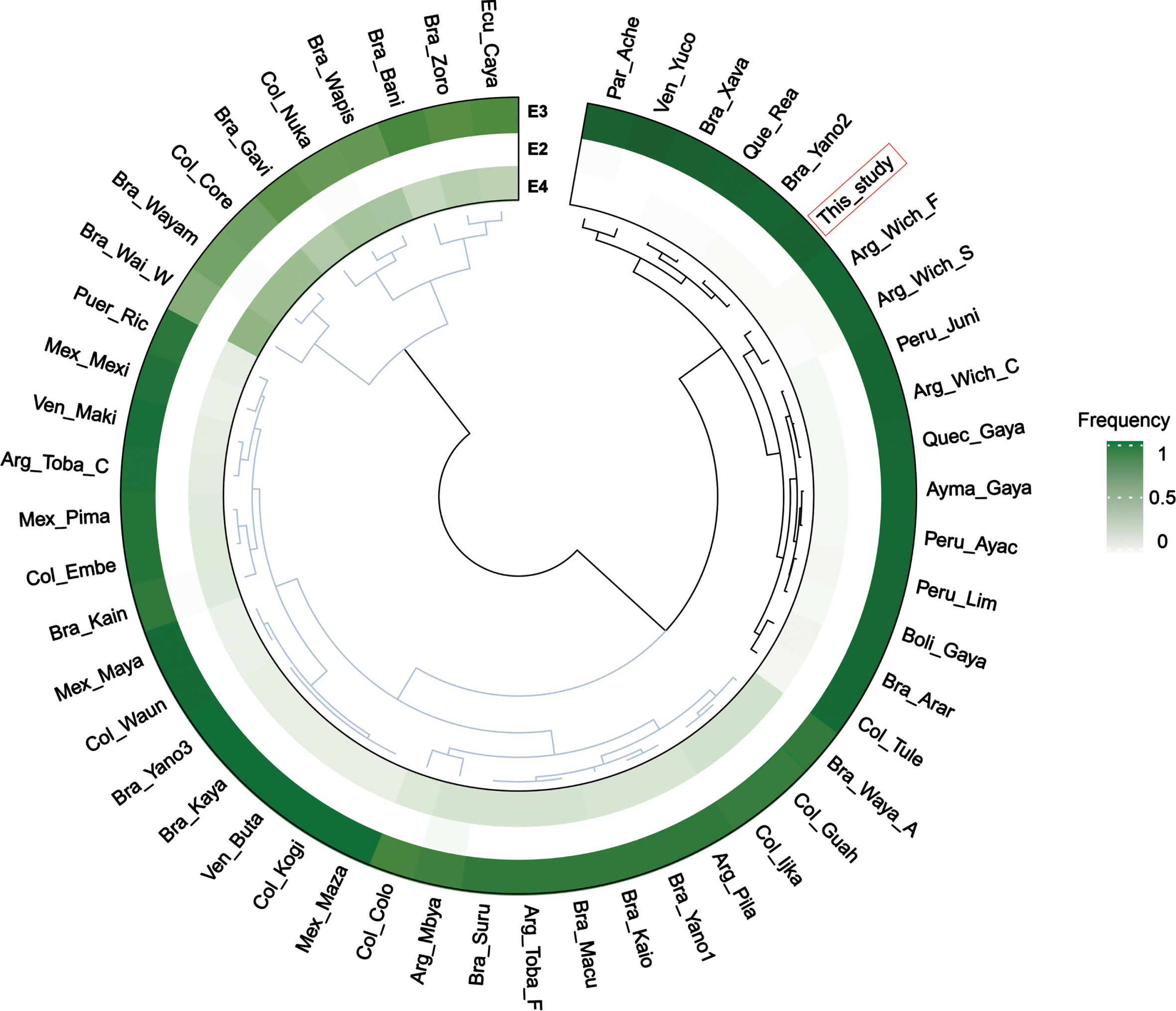
Clustering analysis of APOE allele frequencies in different Native American populations [9]: Arg_Mbya (Argentina – Mbyá–Guarani); Bra_Kain (Brazil – Kaingang); Arg_Toba_F (Argentina – Toba Formosa); Arg_Toba_C (Argentina – Toba Chaco); Arg_Wich_C (Argentina – Wichí Chaco); Arg_Pila (Argentina – Pilagá); Arg_Wich_F (Argentina – Wichí Formosa); Par_Ache (Paraguay – Aché, Paraguay); Bra_Kaio (Brazil – Kaiowá/Ñandeva–Guarani); Arg_Wich_S (Argentina – Wichí Salta); Bra_Xava (Brazil – Xavante); Bra_Suru (Brazil – Suruí); Bra_Zoro (Brazil – Zoró); Bra_Gavi (Brazil – Gavião); Bra_Kaya (Brazil – Kayapo); Bra_Arar (Brazil – Arara); Bra_Waya_A (Brazil – Wayana–Apalai); Bra_Wayam (Brazil – Wayampi); Bra_Wai_W (Brazil – Wai Wai); Ecu_Caya (Ecuador – Cayapa); Bra_Bani (Brazil – Baniwa); Col_Core (Colombia – Coreguaje); Col_Embe (Colombia – Embera); Col_Nuka (Colombia – Nukak); Col_Tule (Colombia – Tule); Bra_Yano1 (Brazil – Yanomami 1); Bra_Yano2 (Brazil – Yanomami 2); Bra_Yano3 (Brazil – Yanomami 3); Bra_Wapis (Brazil – Wapishana); Col_Waun (Colombia – Waunana); Bra_Macu (Brazil – Macushi); Col_Guah (Colombia – Guahibo); Ven_Maki (Venezuela – Makiritare); Ven_Yuco (Venezuela – Yuco); Ven_Buta (Venezuela – Butaregua); Col_Ijka (Colombia – Ijka); Col_Kogi (Colombia – Kogui); Mex_Maza (Mexico – Mazatecan); Mex_Maya (Mexico – Maya); Mex_Pima (Mexico – Pima/Maricopa/Papago); Que_Rea (Quechua [28]); Ayma_Gaya (Aymara [12]); Quec_Gaya (Quechua [12]); Boli_Gaya (Bolivia [12]); Peru_Ayac (Ayacucho [29]); Peru_Juni (Peru (Junin [29]). This_study (Argentina – This study). Current American populations [10]: Col_Colo (Colombians from Medellin, Colombia); Mex_Mexi (Mexican Ancestry from L.A. USA); Peru_Lim (Peruvians from Lima, Peru); Puer_Ric (Puerto Ricans from Puerto Rico).
DISCUSSION
The distribution of alleles conformed to the pattern observed in other Andean populations and South American Native Americans [8, 9]. These populations are characterized by a low frequency of APOE4, and the absence of the potentially most detrimental E4/E4 genotype, with the exception of Gavião (7%), Zoró (17%), Wai-Wai (7%), and Myá-Guarani (4.5%), which are non-Andean populations.
The infrequent occurrence of E4, even in a heterozygous state, suggests that this allele could be significant in explaining the epidemiology of Alzheimer’s disease, coronary disease, and longevity among the Native Andean population. In contrast, this genotype appears to be linked with these illnesses in communities of diverse ethnicities. We must consider that, in Andean populations, there have been a limited number of molecular or epidemiological studies on these conditions and their relationship with APOE, making the analysis more intricate [18]. According to the GBD 2016 Dementia Collaborators group (2019), the worldwide proportions of age-standardized death rates for Alzheimer’s disease experienced a 3.6 change (1.1 to 5.6) between 1990 and 2016. In Argentina, Bolivia and Peru, the change was –2.6 (–10.5 to 5.7), 2.9 (–11.7 to 2.3) and –4.0 (–19.2 to 12.8), respectively. During this time frame, the global age–standardized prevalence rate experienced a percentage change of 1.7 (1.0 to 2.04). Conversely, the prevalence rate in Argentina, Bolivia, and Peru decreased by –0.4 (–7 to 0.7), –3.5 (–6.2 to –0.5), and –7.5 (–11.6 to –3.4), respectively. According to the 2019 Global Burden of Disease Database, the worldwide standardized incidence and prevalence rates of Alzheimer’s disease were 7.24 (6.22–8.23) and 51.62 (44.28–59.02) cases per million, respectively. These figures were 0.045 (0.04–0.05) and 0.29 (0.25–0.34) for Andean Latin America, respectively [19, 20].
Population differences have also been detected in relation to the burden of cardiovascular diseases. For example, global ischemic heart disease age-standardized incidence in 2017 was 135.6 (122.2 to 150.4), while in Andean Latin American population was 78.8 (68.5 to 90.1) [21].
Contradictory findings have emerged from case/control studies assessing the impact of APOE in Alzheimer’s disease in admixed Latin American populations. Moreno et al. [22] found no significant differences in the proportion of Native American ancestry between Colombian carriers and non-carriers of APOE4, despite both groups having a similar allele frequency. Conversely, Marca et al. [23] revealed that local Amerindian ancestry considerably increased the risk of Alzheimer’s disease in APOE4 carriers in Peru. It is worth noting, however, that investigating dementia in Native American groups and obtaining reliable statistics regarding this issue presents substantial impediments due to cultural and linguistic dissimilarities.
A metanalysis by Garatachea et al. [24] revealed that extreme longevity (>100 years) is negatively associated to the E4 allele and the genotypes E4/E4, E3/E4, and E2/E4 while positively to E2/E3. There have been reports of extreme longevity in some Andean populations of Peru (Huancavelica) and questioned in Ecuador (Vilcabamba) [25, 26]. According to the 2010 Bolivian census, population > 65 years old was 9.8% in the departments of the Andean and Sub-Andean regions, in contrast with 5.8% in the Llanos Regions (lowlands).
The APOE system is revealed as a useful marker to analyze new and insufficiently explored traits such as longevity and dementia regarding the adaptation to an extreme environment such as high altitude and lifestyle.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
The authors have no funding to report.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVALABILITY
Data sharing is not available as it was not explicitly stated in the consent form signed by the participants.