
Abstract
Many organisms, including humans, have evolved dynamic social behaviors to promote survival. Public health studies show that isolation from social groups is a major risk factor for adverse health outcomes in humans, but these studies lack mechanistic understanding. Animal models can provide insight into the molecular and neural mechanisms underlying how social isolation impacts health through investigations using genetic, genomic, molecular, and neuroscience methods. In this review, we discuss Drosophila melanogaster as a robust genetic model for studying the effects of social isolation and for developing a mechanistic understanding of the perception of social isolation and how it impacts health.
1 Introduction
Social isolation as a major risk factor for negative health outcomes has received attention from public health experts and scientists over the last few decades, but the topic is more relevant now than ever before. Public health mitigation measures intended to prevent the spread of COVID-19, such as social distancing and quarantine, have caused unprecedented social isolation on a global scale since 2020. People reported increased feelings of loneliness, which can be defined as perceived social isolation, after less than three weeks of following shelter-in-place orders [1].
Social neuroscience studies have previously investigated the negative health outcomes of perceived social isolation in humans. Loneliness is associated with increased risk of depression and suicidal thoughts [2 –5] as well as cardiovascular disease [6, 7] and cognitive decline [8].
Social deprivation increases the risk for early mortality [6, 9 –13]. Importantly, this correlation was found to be independent of socioeconomic status and other factors such as smoking, alcohol consumption, obesity, physical activity, and utilization of preventive health care [14].
Social connections are important across the lifespan. Although many prior isolation studies focus on older adults, mental health problems including depression, anxiety, and sleep disturbances were prevalent among children and adolescents during the COVID-19 pandemic [15]. Other studies show that social isolation correlates with mental health disturbances in young people. Notably, the duration of social isolation (i.e. chronic vs. acute) correlates even more strongly with depression than intensity of isolation [16]. One study of young adults found that depression was significantly correlated with both objective and subjective social isolation [17].
Animal models are instrumental for studying the neurobiology of emotion [18]. It is challenging to study the perception of social isolation in animals because they are unable to report loneliness. However, social isolation in animals is associated with robust behavioral and physiological outcomes in both natural and laboratory settings. This allows researchers to search for genetic and neural mechanisms underlying the perception of social isolation and investigate how this phenomenon can transform other behaviors such as feeding and sleep.
Animals such as macaques and cichlid fish exhibit less social or cooperative behavior following a period of isolation [19, 20]. Behavioral changes due to social isolation are often coupled with physiological and developmental changes. When isolated, ants show impaired mushroom body development [21]. Isolated macaques also show changes in brain structure and function, such as lower gray matter density and less activity in the frontal and temporal cortex, which are associated with smaller network size [22]. Isolated piglets display neuroendocrine changes such as higher levels of stress hormones like cortisol [23]. Social isolation of prairie voles disrupts the gut–brain axis, resulting in altered gut microbiome composition, neuronal activation, and neurochemical expression [24].
Social isolation has profound effects on human and animal behavior and physiology, further illuminated by the worldwide COVID-19 pandemic. Studies in popular model organisms such as mice look to elucidate the molecular and genetic underpinnings of social isolation-induced behavioral changes [25]. However, it is often expensive and time-consuming to conduct isolation experiments in mice. Drosophila melanogaster is a more convenient model organism in many respects. Researchers have exploited the short life cycle, large population size, unparalleled molecular and genetic toolkit, and tractable nervous system of the fruit fly to uncover mechanisms underlying social isolation-induced behavioral and physiological effects.
Social isolation of Drosophila melanogaster causes behavioral changes at various levels of social organization: individual behavior such as sleep [26, 27] and feeding [26], two-animal behavior such as aggression [28 –32] and courtship [33 –35], and collective interaction dynamics [36 –39] (Fig. 1). In this review, we demonstrate animal models such as fruit flies are valuable tools for exploring the behavioral effects of social isolation and their molecular and genetic bases.
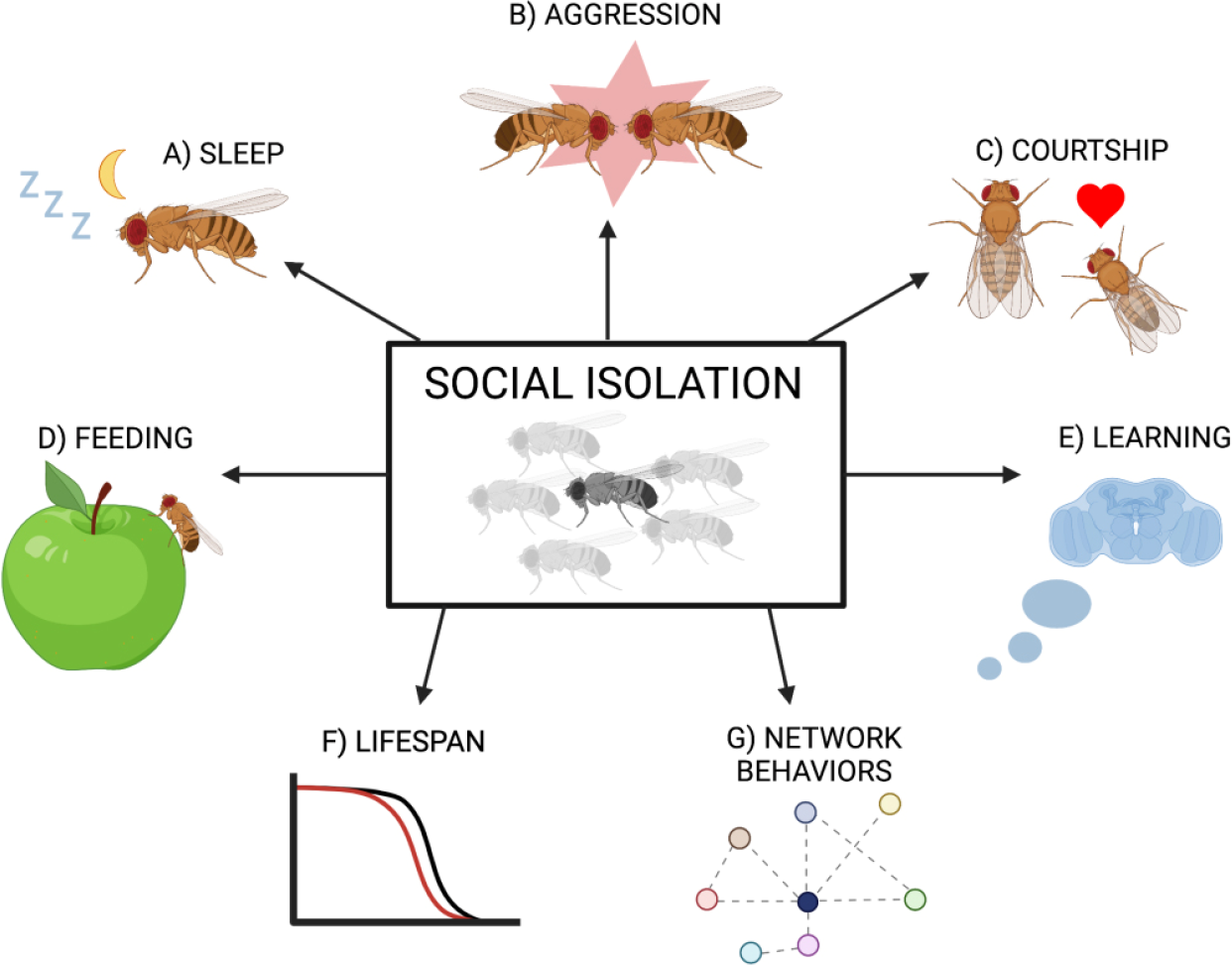
Changes induced by social isolation in Drosophila. A) Isolated Drosophila exhibit reduced and fragmented sleep [26, 27]. B) Isolation-induced aggressive behavior has been found to be regulated by drosulfakinin, reactive oxygen species, cytochrome P450, Or65a olfactory neurons, FruM+ neurons, and CAP neurons [28, 29, 31, 67, 71, 72]. C) Some isolation-induced effects on courtship behavior are dependent on the gene mutant sex pistol and Or47b neurons [33, 35]. D) Social isolation induces a starvation brain state in which fruit flies eat more, and metabolic genes such as limostatin and drosulfakinin are differentially expressed between isolated and group-housed flies [26]. The sleep and feeding responses are controlled by P2 neurons. E) Social learning and memory capabilities are reduced in isolated fruit flies [56]. F) Lifespan is reduced in both male and female isolated flies, but healthspan changes are sex-specific [49]. G) Previously isolated flies exhibit altered group interactions [39]. (A. Vora – created with BioRender.com)
2 Fruit flies as a model for social isolation
Model organisms such as Drosophila melanogaster have contributed to numerous breakthrough discoveries in all fields of biology, including genetics, developmental, and behavioral neuro-biology. High-throughput studies using fruit flies are cost-effective due to their fast generation time and exponential multiplication. Drosophila melanogaster has a well-defined genome and offers diverse genetic tools, including tissue-specific gene expression with the UAS/Gal4 system [40, 41]. Many human genes have fly homologs, making genetic discoveries in fruit flies a good starting point for studying the same topics in humans. Researchers have also developed a high-resolution 3-D reconstruction of the neural circuitry in the Drosophila central brain [42].
Fruit flies are a robust model organism for the study of many social behaviors across the lifespan. For example, they exhibit complex courtship behaviors when identifying and choosing mates and display aggression in certain social contexts and when resources are scarce [43, 44]. Drosophila show collective behaviors and organize into dynamic social networks that are dependent upon group connections. As larvae, Drosophila aggregate and collectively burrow into food together [45]. As adults, they display collective feeding and foraging behaviors [46, 47].
3 The impact of isolation on individual behaviors
Across numerous studies, flies exhibit a multitude of physiological and behavioral changes induced by social isolation such as: reduced and fragmented sleep [26], increased feeding [26], resistance to ethanol sedation [33], increased cellular stress and activation of cellular homeostatic pathways [48], and reduced lifespan and healthspan [49].
Li et al. (2021) reports that chronic social isolation (5–7 days), but not acute isolation (1–3 days), induces sleep loss in the laboratory wildtype strain of D. melanogaster [26]. Chronic isolation not only reduced the total amount of daily sleep but also altered sleep architecture, particularly at dawn and during the daytime [26]. Li et al. (2021) also identified a neural mechanism underlying chronic social isolation-induced sleep loss and overeating. They identified Drosophila P2 neurons that encode the chronic status of social isolation. When these P2 neurons were artificially activated during acute social isolation, flies behaved in a way reminiscent of chronic social isolation [26].
An earlier study by Ganguly-Fitzgerald et al. (2006) shows that socially enriched flies, that typically sleep longer than socially isolated flies, turn into short sleepers after being isolated for four days, whereas isolated flies sleep longer once exposed to social enrichment [27]. In a study by Chi et al., (2014) flies that were isolated as larvae had disrupted sleep patterns as adults [50]. Other organisms such as honeybees also exhibit disrupted sleep when isolated [51]. Studies in social psychology have established that poor sleep quality is a link between persistent loneliness and adverse health outcomes, underscoring the importance of the relationship between social isolation and sleep disruption [52].
Social isolation-induced sleep loss also results in metabolic changes. Li et al. (2021) reports that chronic isolation signals hunger in D. melanogaster despite isolated flies having unrestricted access to food. These isolated fruit flies experience a starvation brain state, resulting in excessive feeding [26]. Transcriptomic analysis of RNA samples from group-housed, acutely isolated, and chronically isolated flies revealed 214 candidate genes for the isolation-induced sleep loss. A number of the candidate genes are relevant to a variety of metabolic processes and sleep, such as Limostatin and Drosulfakinin, consistent with the reduced sleep and overeating phenotype [26].
Other animals, especially other insects, also exhibit metabolic changes after social isolation. When isolated with food, the desert ant Cataglyphis niger does not survive as long as isolated starved ants and group-housed fed ants—potentially due to isolation-induced digestion issues [53]. On the other hand, isolated female cockroaches ate less than their group-housed counterparts, although this effect is related to oocyte production and hormonal release [54]. Socially isolated mice exhibit increases in body weight, obesity, and type 2 diabetes due to increased food consumption and decreased energy expenditure [55].
Studies have also reported aging and longevity changes caused by social isolation. In one study, flies exhibited reduced lifespan in a sex-specific manner with males experiencing a 20% reduction in lifespan and females experiencing a 12% reduction in lifespan [49]. The study also assessed changes in functional senescence by testing climbing ability after social isolation. In isolated female flies in particular, functional senescence was reduced, indicating a lower healthspan. On the other hand, isolated male flies had greater functional senescence than their paired counterparts [49].
Isolation during development can negatively impact social learning in fruit flies and other insects. In a 2018 study, flies that were isolated as larvae and into adulthood communicated less effectively with a socialized “teacher” fly [56]. In another study, worker honeybees (Apis mellifera) that were chronically isolated exhibited reduced performance on discrimination learning tasks compared to bees housed in groups of 8 or 32 [57]. The same study showed an inverse relationship between group size and sucrose responsiveness, with isolated bees being the most responsive. This suggests that social interaction influences the honeybee reward system. These isolated honeybees had the smallest hypopharyngeal glands and the lowest dopamine levels of all three group sizes (1, 8, or 32) [57].
Social isolation affects fly physiology on a molecular level. Brown et al. (2017) reports that isolation-induced sleep loss acts as a stressor on cells, resulting in activation of the unfolded protein response (UPR) [48]. The UPR is a well-conserved homeostatic response to stress in the endoplasmic reticulum [58]. Its purpose is to promote cell survival, however it can induce apoptosis or necrosis if the UPR response is insufficient to relieve cellular stress. The UPR is involved in a variety of diseases such as bipolar disorder and other psychiatric disorders, conformational diseases such as Alzheimer’s and Parkinson’s, metabolic diseases such as diabetes, and even some cancers [58, 59]. Induction of the UPR in response to social isolation is reversible and highly dependent on sleep loss—when isolated flies sleep more, activation of the UPR was lower [48]. Other models such as rats show stress responses when isolated as well, seen by higher levels of corticosterone, among other effects [60]. Isolated mice also display higher levels of corticosterone and a stress response reminiscent of post-traumatic stress disorder [61].
4 The effect of social isolation on two-animal behaviors
4.1 Aggression
Drosophila melanogaster have evolved aggressive behaviors to compete for territory, resources, and mates [44]. These innate behaviors are often influenced by genetic and environmental factors such as isolation. Isolation- induced aggression is an established phenomenon demonstrated in many animals, including mice, rats, and ducklings [62–64]. Drosophila studies characterize specific agonistic behaviors associated with fighting such as “lunging”, “tussling”, and “boxing” [44]. Using these criteria, a quantitative framework was developed for analyzing behavioral differences, qualifying Drosophila as a useful model system to study isolation-induced aggression. Early studies identified increased agonistic behaviors in isolated flies compared to group-housed flies, varying based on age and sexual experience [32].
Other insects beyond fruit flies, such as the Mediterranean field cricket, also display increased aggressive behavior post-isolation. Crickets that were isolated for as little as three hours showed increased aggressive behavior compared to group-housed crickets [65]. Long-term isolation of male mice also results in aggressive behavior, such as physical contact and attacking other mice [66].
More recent studies have focused on the molecular and genetic underpinnings of isolation-induced aggression in male fruit flies. A 2008 study established that social enrichment in fruit flies reversibly suppresses aggressive behavior [29]. The study identified a cytochrome P450 gene, Cyp6a20, as a target for both genetic and environmental factors in mediating isolation-related aggressive behaviors. The Cyp6a20 gene is known to suppress aggressive behaviors and is upregulated with social experience, resulting in isolated flies showing higher levels of agonistic behavior. A 2011 study found that olfactory communication via the pheromone 11-cis-vaccenyl modulates male Drosophila isolation-induced aggression [67]. The study shows that Or65a olfactory pheromone-responsive neurons regulate the effect of prior social enrichment on aggressive behavior.
Drosulfakinin (Dsk) is a Drosophila homolog of vertebrate cholecystokinin, a protein known to modulate aggression and social behavior in mice and other mammals. In fruit flies, Dsk modulates sexual arousal and fighting behavior [68 –70]. Agrawal et al. established the neuropeptide Dsk as a regulator of isolation-induced aggression in D. melanogaster [28]. Knocking down this protein resulted in increased aggression in socially isolated flies [28]. In Li et al.’s work (2021), Dsk expression was also found to be decreased 2.03-fold after social isolation [26].
Female fruit flies also exhibit aggression, however these behaviors often look different than male aggression [30]. Post-isolation, females show more aggression than when group-housed, similar to male flies. Ueda et al. (2009) developed a molecular and genetic framework for isolation-induced aggression in female D. melanogaster [31]. They found that isolation changed nerve and muscle excitability, and that two gene mutants involved in reactive oxygen species (ROS) metabolism, Hyperkinetic (Hk) and glutathione S-transferase-S1 (gsts1), increased female aggression in both isolated and group settings [31]. Therefore, isolation-induced aggression is potentially sensitive to ROS metabolism, particularly in female flies.
Other studies focusing on the neural circuitry underlying aggressive behaviors in Drosophila strengthen our knowledge of the relationship between social isolation and aggression, as well as sex differences in aggressive behaviors. Chiu et al. (2021) identified a circuit logic to explain the shared and differential aggressive behaviors of male and female flies [71]. Common aggression-promoting (CAP) neurons control aggressive approaching in both males and females, while the homologous male aggression-promoting (MAP) neurons in males and fpC1 neurons in females control attacking [71]. When socially isolated, this dimorphic quality of neuronal activity is enhanced, suggesting that these neurons may be significant in isolation-induced aggressive behavior.
Asahina et al. (2014) discovered that FruM+ neurons, which release the neuropeptide tachykinin, mediate intermale aggressive behavior, while leaving male-female courtship behavior unaffected [72]. FruM is a male-specific isoform of the fruitless gene, the products of which are significant mediators of sex-specific behavior [73]. In mice, chronic social isolation causes whole-brain upregulation of tachykinin, whereas administering a tachykinin antagonist blocks the typical behavioral effects seen during social isolation [62]. The neuropeptide tachykinin has been found to mediate aggressive behavior in mammals as well [74].
4.2 Courtship
Courtship and mating behavior in Drosophila involves complex rituals that are highly plastic and influenced by the social environment, especially for male flies. Mating behavior is interrelated with aggressive behavior. For example, recent findings identify a genetic mutant called sex pistol (sxp) that results in male flies displaying higher levels of aggression post-isolation as well as more male-male courtship behavior [33]. Sex pistol is an insertion affecting the hts and CalpA genes. This mutant produces slightly reduced expression of Calpain A and several hts (adducin) isoforms, and higher expression of insulin-like peptide dILP5. Isolated sxp mutant flies showed decreased levels of activity and greater resistance to ethanol sedation than isolated wild-type flies [33].
Other studies assess how social isolation affects courtship success. Mature male fruit flies reared in groups had a better advantage in courting females than those reared in isolation due to better pheromone detection via the Or47b olfactory receptor neurons [35]. The sensitivity of these neurons to population density and the propensity for less effective courtship behavior after isolation is regulated by a calcium signaling pathway involving calmodulin-dependent protein kinase 1 (CaMK1) and CREB binding protein (CBP) downstream of CaMK1 [35].
Plasticity in mating behavior after isolation is also seen via changes in courtship song. Male fruit flies participate in courtship singing by producing wing vibrations, allowing females to choose mates. Male Drosophila melanogaster exhibited longer song bursts when housed with other males than when isolated [34]. Marie-Orleach et al. (2018) argues that the longer song bursts in group-housed male flies could result in greater stimulation for females and therefore greater likelihood of successful mating [34]. Vocal communication in mice is also important for courtship and is affected by the social environment. Keesom et al. (2017) found that early-life isolation of male mice resulted in significant differences in ultrasonic vocal communication such as longer and lower frequency songs, especially when paired with a group-housed same-sex mate [75].
5 The impact of social isolation on collective behaviors
Although fruit flies are not eusocial insects, they form social networks and participate in collective behaviors such as foraging, feeding, and risk-aversion communicated by tactile, acoustic, visual, and chemosensory signals [39, 47, 76]. Isolation significantly affects these collective behaviors. For example, a 2015 study by Ramdya et al. shows that as group size decreases, flies avoid aversive odors less and less, with isolated flies spending very little time avoiding these odors [76]. Flies also engage in collective foraging behaviors such as communicating the presence and quality of food through olfactory and chemosensory cues [47]. Isolated flies are less able to remember and respond to these cues.
Studies utilize video recording and statistical techniques analyzing various parameters to elucidate patterns in interactions and quantify network behavior. Recently, these techniques have improved significantly with the use of machine learning and high-resolution computer vision algorithms, opening the door to studying fly networks more extensively. However, as these advanced techniques are newly established, there is limited and conflicting data regarding the effects of the social environment on network dynamics.
A 2012 paper by Schneider et al. using “fixed criteria” analysis found that there were no differences in social structures when flies were previously isolated or group-housed [38]. However, when the data was re-analyzed by Jezovit et al. (2021) with “automated criteria”, the results led to a different conclusion [39]. Flies that were isolated upon eclosion exhibited longer periods but lower rates of interaction overall than group-housed flies, and reciprocated interactions less frequently [39].
There is further conflicting data regarding the influence of social experience on fruit fly networks. In a 2018 paper, Liu et al. (2018) developed a video-recorded arena and a computer vision setup called Flyworld to analyze network dynamics [37]. Their data showed that flies isolated for 6 days had a higher “clustering coefficient” than socially enriched flies and therefore showed stronger interactions with neighboring flies. On the other hand, Bentzur et al. (2021) showed differing results when conducting similar experiments scoring Drosophila melanogaster social behavior in an arena after exposing them to a certain social environment [36]. In this study, isolated male flies had a significantly lower “social clustering” value and therefore formed less structured, complex, and stable groups than flies with prior social experience [36]. These findings are consistent with Simon et al. (2012) and Scott et al. (2018) who found that flies that were previously isolated exhibited a greater inter-fly distance and less sociability than socially enriched flies, respectively [77, 78]. While these studies do not completely contradict Liu et al. (2018) [37], the results are different in significant ways. Jezovit et al. (2021) concluded that the variability in results can be chalked up to differences and advancements in the analysis process, as well as methodological variation [39].
Network dynamics and the effects of the social environment are well-established in other organisms, especially eusocial insects such as ants and honeybees. When fire ant queens set up new colonies, they are forced to react to the social environment—they either found a colony while completely isolated or found a colony with a group of other queens. One study found that early in life, plasticity in this behavior is controlled by one gene, but later in life a myriad of genes are required to adapt to different environments [79]. Gene expression changes were also broad when ant queens formed colonies in groups, whereas expression changes were limited when queens were isolated or in small groups [79].
6 Discussion
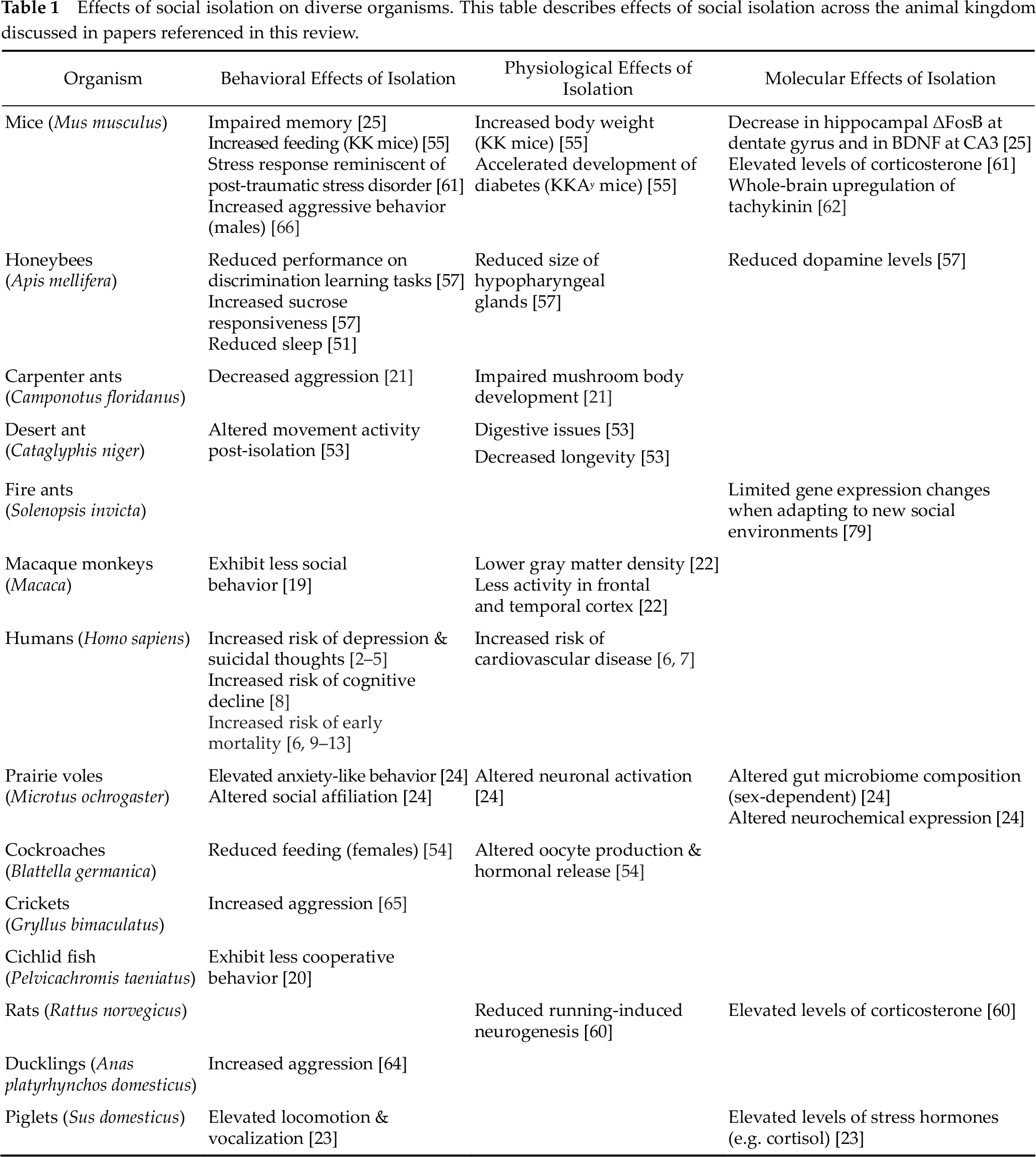
In summary, it’s clear that social isolation affects organismal behavior and physiology at every level of social organization. Many isolation-induced phenomena are found to be conserved across the animal kingdom. The effects of isolation on longevity, sleep, feeding, metabolism, gene expression, epigenetics, learning and memory, aggression, courtship, and social networks are echoed in a diverse group of species – from eusocial insects such as honeybees and ants to mammals such as mice, monkeys, and humans (Table 1).
Effects of social isolation on diverse organisms. This table describes effects of social isolation across the animal kingdom discussed in papers referenced in this review.
While many studies in humans recognize the increased risks of mortality and mental health issues associated with social isolation and loneliness, the mechanisms linking perceived social isolation and negative health conditions remain unknown. Cardiovascular activation and sleep dysfunction have been hypothesized as potential mediators of loneliness on overall health but a causal relationship has not yet been demonstrated [80]. It is ethically and logistically challenging to conduct loneliness experiments on humans. And thus there has been a longstanding call for comparative studies and animal models for studying loneliness and social isolation [81].
Human studies documenting the direct effects of social isolation are observational while animal studies conventionally use social isolation as a strong stressor. However, how social isolation is perceived, centrally processed, and in turn how it influences health behavior is not well studied. A deeper understanding of social isolation-associated phenomena requires further investigation into the molecular and neural pathways that link the perception of social isolation to behavioral and physiological outcomes. Recent theoretical works suggest that these neural pathways are homeostatic regulators of social connectedness, producing direct behavioral and physiological effects to restore social connections to its set point after isolation [82]. This model of social homeostasis provides a new framework to study the mechanisms that signal a prolonged isolated state and promote behavioral changes associated with chronic social isolation [83].
Investigations in Drosophila have already begun to experimentally identify neurons that encode the chronic status of social isolation. Li et al. (2021) established that P2 fan-shaped body columnar neurons in fruit flies are involved in the perception of the duration of social isolation [26]. Artificial activation of P2 neurons results in misperception of acute social isolation as chronic social isolation [26]. P2 neurons are likely to signal to the homeostat that regulates sleep and feeding in Drosophila. Future work focusing on P2 neurons and its anatomical connections may identify neural substrates and circuits underlying the perception and central processing of chronic social isolation.
Using animal models like Drosophila can be a powerful tool for investigating the mechanisms by which chronic social isolation is perceived and how it impacts critical behaviors at the molecular and neuronal levels. This knowledge can ultimately lead to interventions that will alleviate the suffering caused by social isolation and loneliness.
Footnotes
Conflict of interests
All contributing authors have no conflict of interests.
Funding
Wanhe Li is supported by the Cancer Prevention Institute of Texas and the Seed Grant Program for Promoting Research Collaborations from Texas A&M University.
Acknowledgements
We would like to thank Dr. Michael W. Young for his mentorship and support throughout the writing process. We would like to thank Dr. Zhaoyue Zhang for comments and suggestions.
Authors’ contribution
AV, AN, CS, and WL outlined the review, conducted literature research, wrote the manuscript, and provided revisions.