
Abstract
Memory impairment in children is an ongoing issue worldwide related to a learning disability. This neurobiological condition has been suggested to associate with bisphenol A (BPA) exposure during pregnancy. BPA is an inorganic compound used to produce polycarbonate plastics and epoxy resins. We conduct this study to investigate the effects of prenatal BPA exposure on the level of the N-methyl-D-aspartate (NMDA) receptor subunits, synaptic markers of the hippocampus and neurobehavioral outcomes in rats. The pregnant rats were given a daily dose of 5 mg/kg and 50 mg/kg of BPA with 0.5% Tween 80 orally from gestation day 2 until 21 (GD21). The level of GluN2A, GluN2B, PSD-95 and synapsin I in the hippocampus and its neurobehaviour outcomes were quantified and evaluated in the male foetus and adolescent rat. Prenatal BPA exposure reduced GluN2A, GluN2B, synapsin I and PSD-95 (Postsynaptic Density-95) in the male foetus and adolescent rat hippocampus compared to the control group. The prenatal BPA exposed rats demonstrated anxiety-related behaviour and impairment in aversive and spatial memory. The findings suggested that the impairment in neurobehavioral performance may inhibit the signalling pathway in the NMDA receptor subunits in the male foetus rat hippocampus leading to learning and memory deficits when reaching adolescence.
1 Introduction
Bisphenol A (BPA) is a colourless crystalline solid widely used to produce polycarbonate plastics and epoxy resins. BPA is detected in many applications, particularly in the packaging of food and beverage products, coating of cans, sports products, toy items, dental materials, polyvinyl chloride (PVC) pipe walls, thermal papers and many more [1, 2]. The commercial production of BPA in numerous applications has been warned to be harmful to human health when risk assessment studies showed BPA compounds leach into foodstuff and the environment [3]. Oral diet contributes more than 90% of BPA exposure to other sources such as inhalation and dermal contact [4].
Due to these consequences, Food and Drug Administration (FDA) has set the No-Observe-Adverse-Effect-Level (NOAEL) at 5 mg/kg/day and the Low-Observe-Adverse-Adverse-Effects-Level (LOAEL) at 50 mg/kg/day [3]. Nevertheless, these BPA doses remained controversial as many studies have revealed detrimental effects on animal and human health such as infertility, cardiovascular disease, cancer, diabetes, obesity, neurological disorder and many more [4–6]. The toxicity of BPA in pregnant mothers is of particular concern as unconjugated BPA has been reported to be able to pass through the placenta and transferred to the foetus’s biological system [4]. The presence of BPA has been demonstrated in umbilical cord blood, amniotic fluid, placenta tissue and foetal plasma [7 –9]. The development of the nervous system at the early life stage is believed to be susceptible to any environmental change such as prenatal BPA exposure [10]. Emerging evidence from human and animal studies revealed that maternal BPA exposure is linked with social impairment and anxiety behaviour, lower scores on the intelligence test, depressive and hyperactivity and increased memory error in experimental neuron tasks, particularly in young males [11 –14].
BPA has been recognized as an endocrine disruptor chemical (EDC) as it can alter the hormone receptor expression such as oestrogen receptor and disrupts gene expression of the target tissue [15]. The development of N-methyl-D-aspartate (NMDA) is essential for the efficiency of excitatory synaptic transmission during the matured stage. In addition, the NMDA receptors-mediate plasticity is believed to be closely related to learning and memory function [16]. Recently, the hippocampus has been part of the brain that plays vital roles in learning and memory, spatial navigation, and emotional behaviour [17]. There are predominates of GluN2 subunits, GluN2A and GluN2B, which are coupled to make up the obligatory NMDA receptors. GluN2A and GluN2B subunits were studied as they are primarily found in the hippocampus and cortex than the other subtypes, GluN2C and 2D. The expression of these subunits is dynamic with age where GluN2B is highly expressed at the prenatal stage but decreases into adulthood. At the same time, GluN2A is lowly expressed at the prenatal stage but increases when reaching adulthood [18].
The presence of GluN2 subunits at the presynaptic membrane is stabilized by their interaction with postsynaptic density-95 (PSD-95). GluN2 assists the NMDA in inducing the long-term potentiation (LTP) process [18]. The early success of LTP contributes to the function of a family of protein synapsin I at presynaptic terminals. The phosphorylation of synapsin I have been studied to boost the release of a synaptic vesicle, allowing future action potential [19]. Prenatal BPA exposure in rodents has been reported to inhibit the expression of NMDA receptor subunits followed by learning and memory impairment when reaching the adult stage [15, 20, 21].
Nevertheless, to our knowledge, there is no study involving the foetus. Based on these findings, it is speculated that prenatal BPA exposure may modulate NMDA receptor subunits expression at the foetus stage and is inheritably expressed when reaching adolescence. Hence, the present study was designed to investigate the effects of prenatal BPA exposure on the expression of NMDA receptor subunits in the hippocampus both at the foetus and adolescent stage and, subsequently, its neurobehavioral outcomes. In the present study, the expression of PSD-95 and synapsin I in adolescents revealed its possible relationship with the NMDA receptor subunits during signalling activity.
2 Materials and methods
2.1 Animals and treatments
The male and female Sprague Dawley rats (200–250 g body weight) at age 8–12 weeks were housed in standard cages in a controlled environment (temperature 23–34°C, 12:12 hour light/dark cycle) with free excess to water and food. All experimental procedure was approved by the Committee on Animal Research and Ethics (UiTM CARE) with ethical number UiTM CARE: 351/2021. After mating overnight, a vaginal smear test was checked in the morning and (gestational day 0) GD0 was noted with the presence of sperm. The mothers were individually separated in each dam. The study addresses the issue by looking at the NOAEL and LOAEL of human exposure. From GD2 until GD21, the pregnant mother (N = 8) was orally exposed (ad libitum) to a drinking solution with 5 mg/kg/day (NOAEL) and 50 mg/kg/day (LOAEL) of BPA with 0.5% Tween 80 (vehicle) in reverse osmosis water. The control was consumed the same except without BPA. At GD21, the appropriate number of mothers were anaesthetized and underwent caesarean section. The hippocampus of male foetuses were collected and kept at –80°C. The remaining mothers were monitored until spontaneous delivery and the pups were separated until they finished weaning. Then, the male pups were monitored until the adolescent age of 35 days (AD35) and were assessed with neurobehavioral tests. The males’ hippocampus were collected after the tests and kept at –80°C.
2.2 Tissue preparation and enzyme-linked immunosorbent assay (ELISA)
The hippocampus tissues were extracted by using the Mem-Per Plus membrane protein extraction kit (Thermo Fisher, US) for membrane-localized protein and RIPA lysis and extraction buffer for cytoplasmic-localized protein (Thermo Fisher, US) together with Halt Protease and Phosphatase inhibitor cocktail (Thermo Fisher, US). Coomassie Plus (Bradford) assay kit (Thermo Fisher, US) was used to determine the total protein concentration from each sample. Then ELISA assays were used to quantify the surface expression of the GluN2A (DLDevelop, China), GluN2B (DLDevelop, China), PSD-95 (DLDevelop, China) and synapsins I (FineTest, China). All tests using the respective kits were performed according to the manufacturer’s instructions.
2.3 Neurobehavioral test
2.3.1 Open field test
The open field test is an established method to evaluate exploratory and anxiety-related behaviour in animals [22]. The open field apparatus was a square arena of 100 cm (L) × 70 cm (H) × 100 cm (W) with the installed camera above. The test was conducted for 10 min under dim light. Each male rat was tested on the open field once daily for 6 consecutive days (Fig. 1). The behaviour of the male rats was analysed by total distance travelled, number of crossings of the centre area, number of entries into the peripheral area, freezing episodes, number of grooming and number of faecal boli. All sessions were automatically recorded by ANY-Maze software (Stoelting Co.). The field was wiped with 75% ethanol before each test to prevent the transmission of olfactory cues.
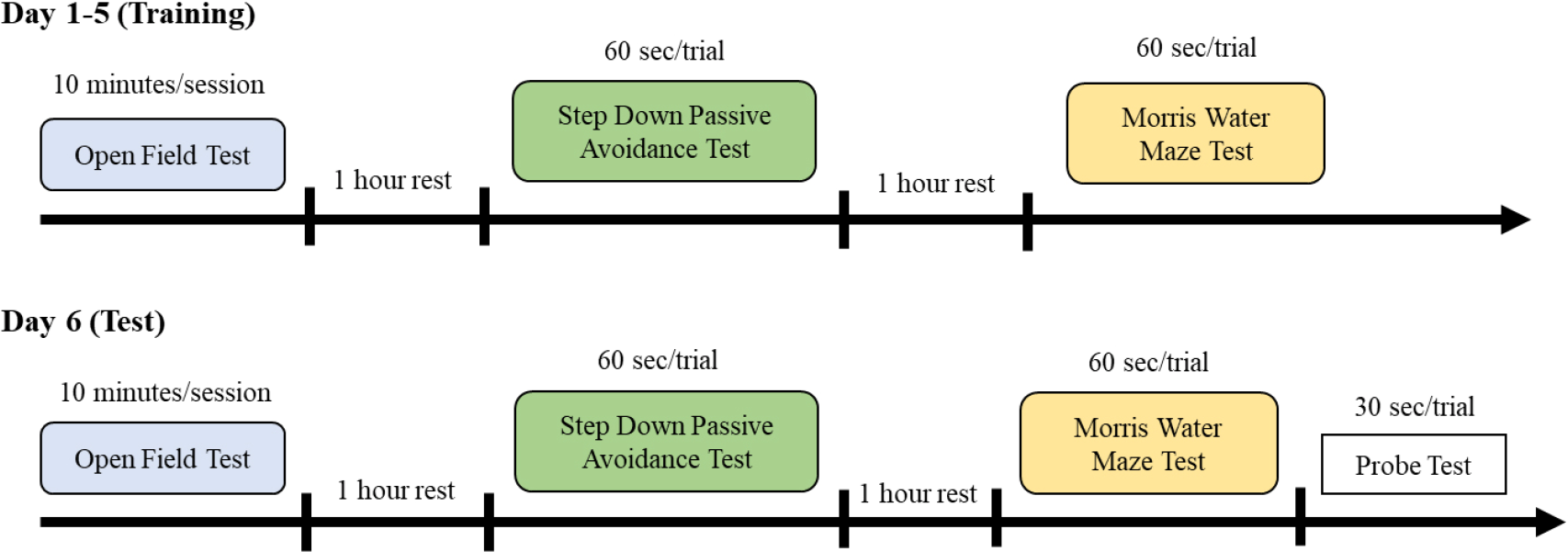
The timeline of the neurobehavioral test.
2.3.2 Step down passive avoidance test
A passive avoidance test is used to evaluate the learning and memory of the animals. The test involves rats trained to avoid the aversive stimulus [23]. The memory retention test was conducted in a cage with an elevated platform (Ugo Basile) for 6 consecutive days. The training (day 1 to 5) and test (day 6) procedure was conducted for 60 s with 4 trials (Fig. 1). After the habituation period (1 min), the male rats were placed individually on an elevated platform, the platform was set to vibrate at 60 Hz and the floor with a steel bar was set with mild electric shock (0.4 mA). The response to foot shock was recorded by measuring the time taken for the rat to step down from the vibrating platform within 60 s.
2.3.3 Morris water maze test
Morris water maze test is designed to study the spatial memory of the animals where they will be trained to locate the hidden platform based on their pre-training memory [24]. The test was conducted in a circular tank filled with water with 150 cm diameter and 60 cm height with a camera installed above. The rats were given 60 s to find the platform with 4 trials based on 4 different quadrants. Each of the male rats was trained to find the platform at the centre area of the pool in clear water for 5 days (Fig. 1). After 5 days of training, the rats were individually tested to find the platform in opaque water (coloured by non-toxic paint). The time taken for the rats to find the hidden platform within 60 s was automatically recorded by ANY-Maze software (Stoelting Co.). Then the rats were subjected to the probe test for 30 s to determine the reference memory, where the platform was removed from the pool.
2.4 Statistical analysis
All data were represented as means ± SEM (standard error of mean). The data were tested for normality using a Shapiro-Wilk test. One-way ANOVA followed by post hoc Bonferroni analysis was used to determine the multiple comparisons of the synaptic proteins (GluN2A, GluN2B, PSD-95 and synapsin I) in the different dosages of BPA. All behavioural data were analysed using two-way ANOVA followed by post hoc Bonferroni analysis. A value of p < 0.05 was considered to be statically significant.
3 Results
3.1 The expression of GluN2A and GluN2B at the foetus and adolescent stage
The ELISA analysis showed that prenatal BPA exposure significantly changed the level of GluN2A (A and C) and GluN2B (B and D) in the foetal and adolescent hippocampus in a dose-dependent manner. According to Fig. 2 BPA treated group at 5 mg/kg/day and 50 mg/kg/day showed a significantly lower level of GluN2A in the foetus (p < 0.05) and adolescent (p < 0.01) hippocampus than the control group. Next, quantification of GluN2B in the foetus (p < 0.001) and adolescent (p < 0.001) hippocampus revealed BPA treated groups at 5 mg/kg/day and 50 mg/kg/day significantly reduced the level of GluN2B compared to the control.
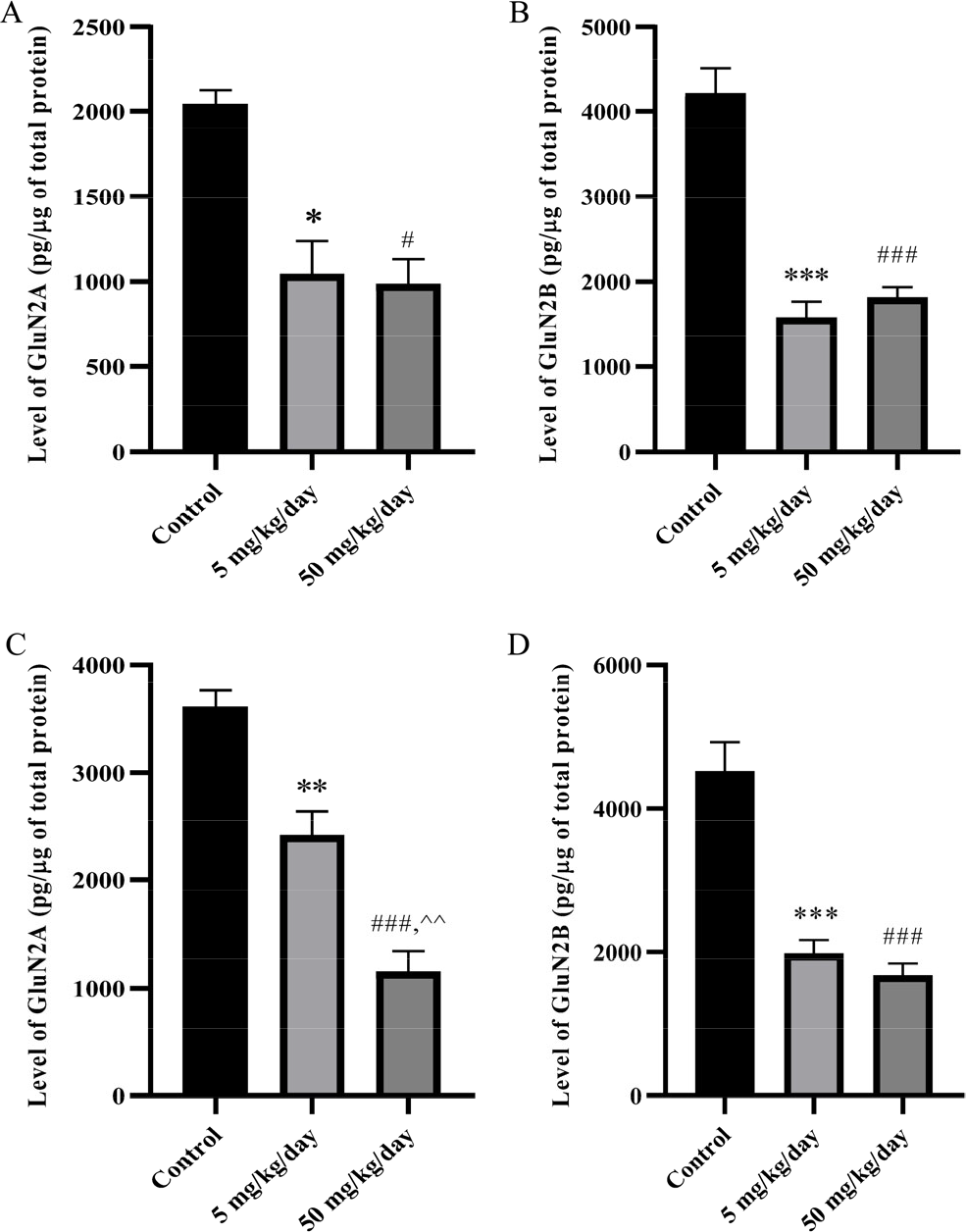
The level of GluN2A at foetus (A) and adolescent (C), GluN2B at foetus (B) and adolescent (D) in male rat hippocampus. The data were shown in pg/μg of total protein, mean ± SEM, n = 4. *, p < 0.05; **, p < 0.01; ***, p < 0.001 (5 mg/kg/day vs. control); #, p < 0.05; ###, p < 0.001 (50 mg/kg/day vs. control); ^^, p < 0.01 (5 mg/kg/day vs. 50 mg/kg/day) by one-way ANOVA followed by Bonferroni analysis.
3.3 The expression of synaptic markers
Besides essential aspects of the development of NMDA receptor subunits, synapsin I and PSD-95 are also crucial in synaptic mechanisms during learning and memory function. The findings from ELISA analysis revealed that prenatal BPA exposure reduced the expression of synapsin I [Fig. 3(A)] and PSD-95 [Fig. 3(B)] in the adolescent rat hippocampus. BPA treatment reduced PSD-95, where 50 mg/kg/day has a significantly (p < 0.01) lower level of PSD-95 in the hippocampus than in the control. The level of synapsin I showed no significant differences between all groups, yet a trend showed BPA treated groups have lower levels of synapsin I than the control group.
3.4 Neurobehavioural tests
3.4.1 Open field test
There was no significant interaction of anxiety-related behaviour found in total distance travelled [Fig. 4(A)] between the groups on all days except for day 1 (D1) between 5 and 50 mg/kg/day groups (p < 0.05). The prenatal BPA exposure demonstrated no significant effects in the number of crossing centre areas [Fig. 4(B)]. There was a significant difference between the groups at D1 (p < 0.01, p < 0.05), D4 (p < 0.05) and D5 (0.05) in the number or entry peripheral area [Fig. 4(C)]. When compared to the controls, BPA-treated groups showed a significant effect on D3 in the time taken for freezing episodes [Fig. 4(D); p < 0.001, p < 0.001]. The number of grooming [Fig. 4(E)] showed there were significant effects on D1 (p < 0.001), D2 (p < 0.05) and D5 (p < 0.01, p < 0.001) between the groups. Lastly, rats from BPA treatment excrete more than the control, as on D1 (p > 0.001, p < 0.01), they showed anxiety-related behaviour significantly. The track plot [Fig. 4(G)] report showed the activity of the rats in the arena during the 10 min test sessions. The centre square was denominated as “central zone” while the outer square was denominated as “peripheral zone”. The control rat [Fig. 4(G), (i)] was observed exploring the whole arena freely considered to have reduced levels of fear and danger. The BPA treated rats [Fig. 4(G), (ii) and (iii)] tended to avoid the centre area as they are frequently exploring the peripheral area showing anxiety-related behaviour.
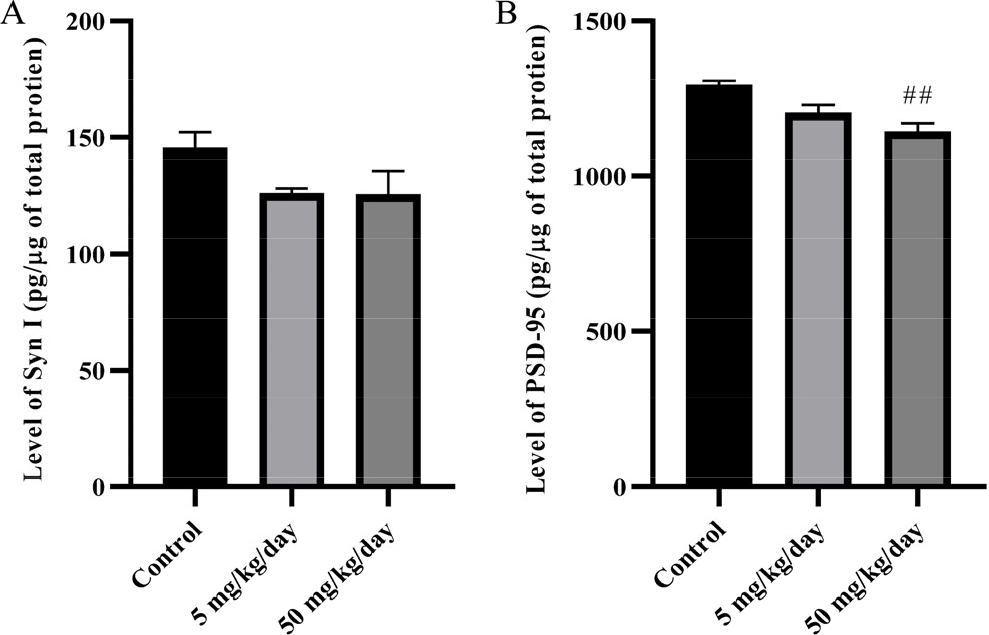
The level of synapsin I (A) and PSD-95 (B) in male adolescent rat hippocampus. The data were shown in pg/μg of total protein, mean ± SEM, n = 4. ##, p < 0.01 (50 mg/kg/day vs. control) by one-way ANOVA followed by Bonferroni analysis.
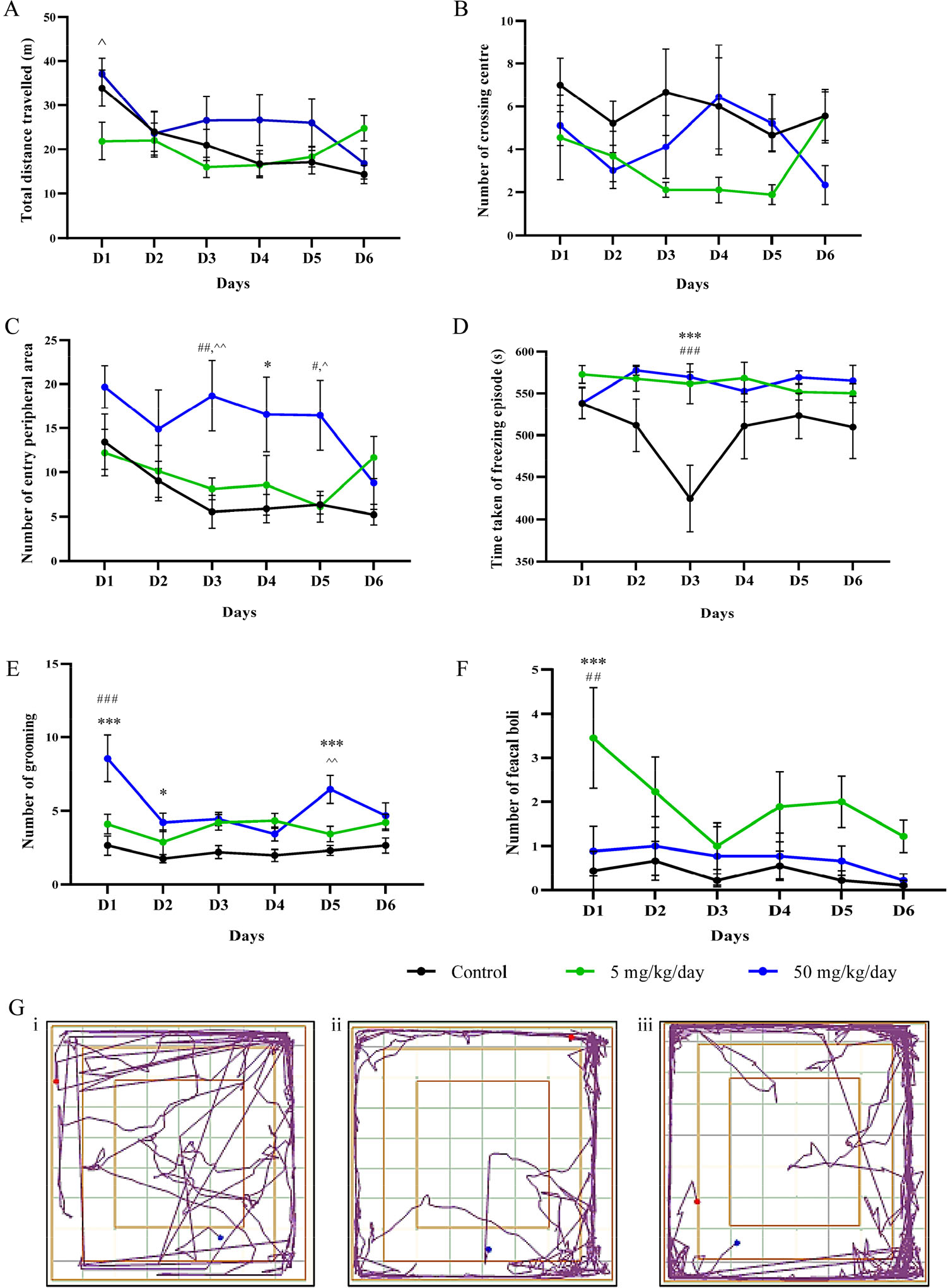
The effect of prenatal BPA exposure in exploratory and locomotor activity of the adolescent male rats using open field test. (A) Total distance travelled. (B) Number of crossing centre area. (C) Number of entry peripheral area. (D) Time taken of freezing episodes. (E) Number of grooming. (F) Number of feacal boli. Data were shown in mean ± SEM, n = 9. *, p < 0.05; ***, p < 0.001 (control vs. 5 mg/kg/day); #, p < 0.05; ##, p < 0.01; ###, p < 0.001 (control vs. 50 mg/kg/day); ^, p < 0.05; ^^, p < 0.05 (5 mg/kg/day vs. 50 mg/kg/day) by two-way ANOVA followed with post hoc Bonferroni analysis. (G) Track plot, (i) control, (ii) 5 mg/kg/day, (iii) 50 mg/kg/day.
3.4.2 Step down passive avoidance
In the training session, only D1 (p < 0.01, p < 0.05) showed significantly impaired aversive memory between the groups based on the latency to step down from the vibrating platform. In D6, the 5 mg/kg/day group significantly reduced the aversive memory performance (p < 0.05). Overall, the finding indicates that the 5 mg/kg/day group experienced higher memory error than 50 mg/kg/day and the control group (Fig. 5).
3.4.3 Morris water maze
The analysis of data showed BPA treated groups committed significantly more memory errors on trials D3 (p < 0.05) and test D6 (p < 0.01 and p < 0.05) compared to the control group [Fig. 6(A)]. The 5 mg/kg/day showed a progressive reduction in spatial memory than others. Other than that, the track plot [Fig. 6(B)] showed the navigation track of the rats in the swimming arena where the 4 boxes represent different quadrants (Q1, Q2, Q3 and Q4). Based on the plot, BPA treated rats [Fig. 6(B), (ii) and (iii)] demonstrated a longer distance travelled than control rats [Fig. 6(B), (i)] showing a reduction in spatial memory. Therefore, the further the distance of the swimming, the longer time takes for the rats to find the escape platform.
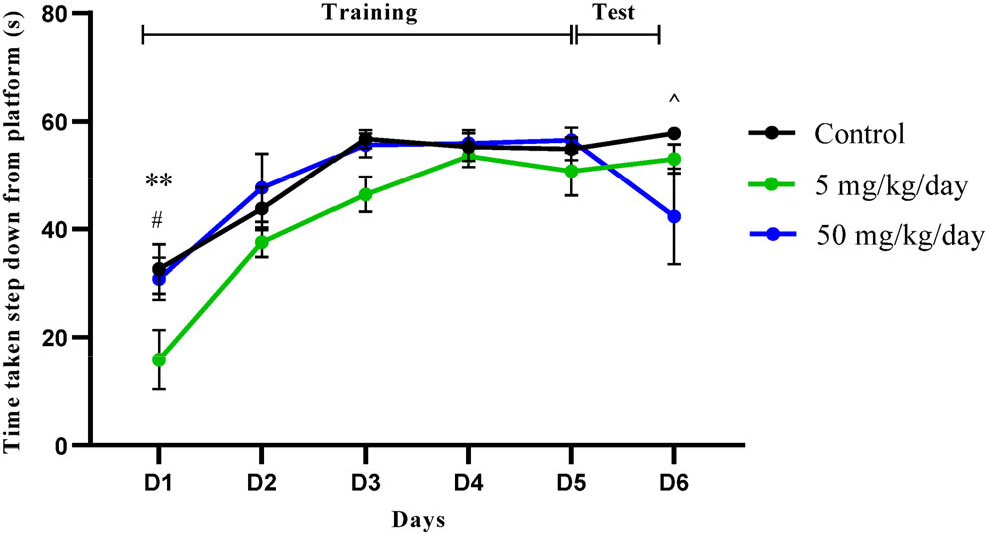
The memory retention test with step down passive avoidance test. Data were shown in mean ± SEM, n = 9. **, p < 0.01 (control vs. 5 mg/kg/day); #, p < 0.05 (control vs. 50 mg/kg/day); ^, p < 0.05 (5 mg/kg/day vs. 50 mg/kg/day) by two-way ANOVA with post hoc Bonferroni analysis.
4 Discussion
Neurogenesis of the foetus during pregnancy has been recognized as the most susceptible stage toward environmental perturbation. In the present study, BPA is mainly selected as an environmental toxicant suspected to disrupt the related receptor signalling in the hippocampus during the foetus and adolescent stages. The first study was done to determine the effects of prenatal BPA exposure on the level of NMDA receptor subunits (GluN2A and GluN2B) and synaptic plasticity marker (synapsin I and PSD-95) in the hippocampus of the foetus and adolescent male rats. Secondly, to understand how modifications at the protein levels by BPA influence synaptic signalling, behavioural studies were done to evaluate hippocampal-related behavioural changes in terms of anxiety state with learning and memory functions of the rats.
The daily BPA dosage was chosen at 5 and 50 mg/kg to represent the NOAEL and LOAEL of BPA exposure on health effects. The present study revealed that continuous prenatal BPA exposure at dosages 5 and 50 mg/kg/day for 21 days of gestation reduced the levels of GluN2A and GluN2B subunits at the foetus and adolescent stage. The levels of PSD-95 and synapsin I at the adolescent stage were also reduced causing increased memory errors in the neurobehavioural test. The findings of current studies suggested the outcomes of the study may be related to the response of BPA metabolism in maternal where BPA has been demonstrated to cross the placenta in an active unconjugated form [9, 25] The immature liver of the foetus has a lower level of the enzyme UDP-glucuronosyltransferase (UGT), which limits the metabolism of BPA and leads to bioaccumulation of BPA in the foetus where the brain has been suspected as a susceptible organ toward BPA exposure [26]. In the developing brain, BPA reportedly can bind strongly with the oestrogen related receptor γ (ERRγ). This interference of ERRγ might lead to misregulation of genes translations in neurodevelopment [27]. Then, this might alter the epigenetics expression where the gene is converted into functioning proteins or inherited by the subsequent generation [28]. These molecular mechanisms mediating the effects of BPA on brain development, specifically the hippocampus, might be the fundamental cause of the findings of this study.
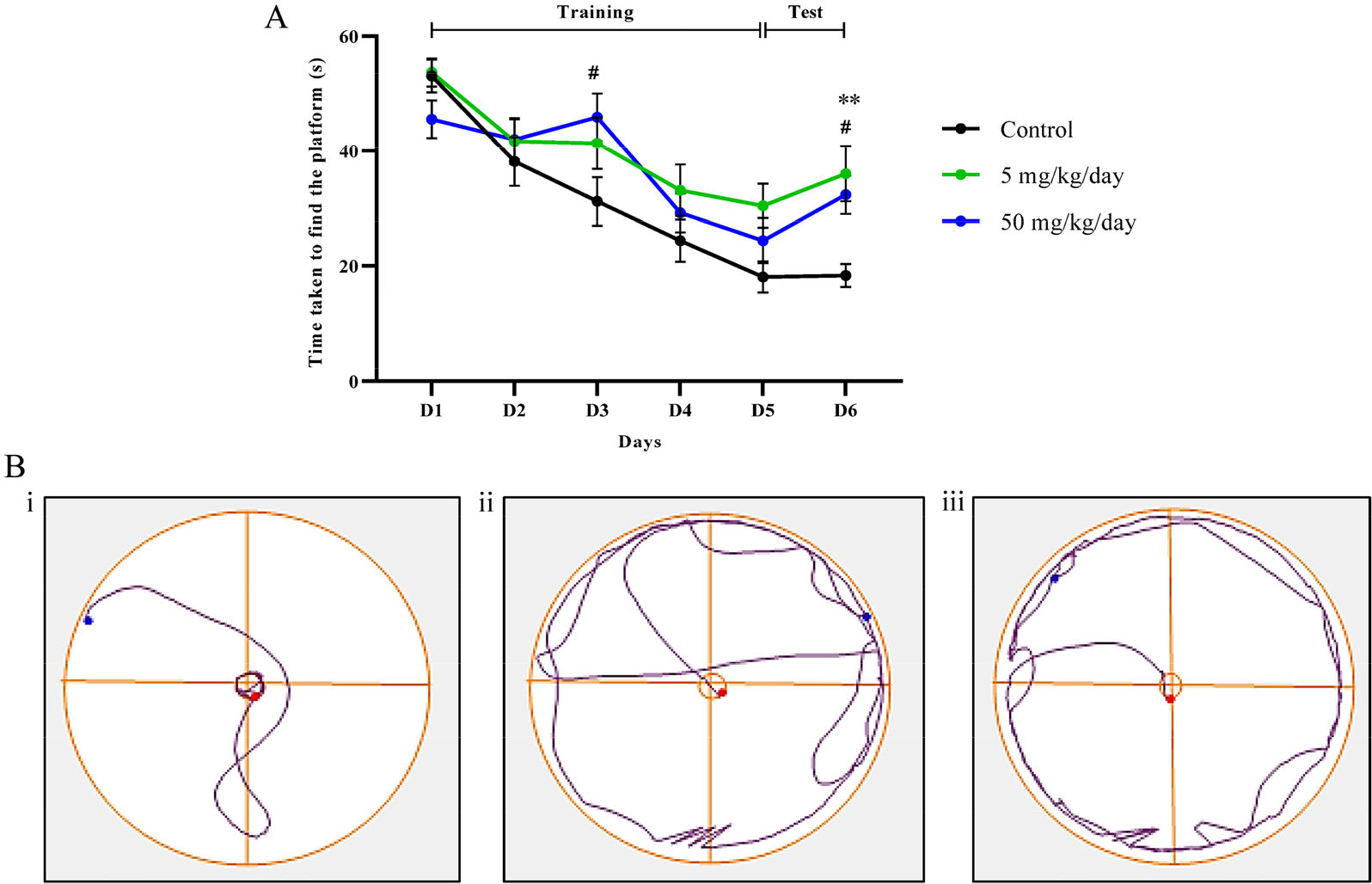
The memory retention test with Morris water maze. (A) Data were shown in mean ± SEM n = 9. **, p < 0.01 (control vs. 5 mg/kg/day); #, p < 0.05 (control vs. 50 mg/kg/day) by two-way ANOVA with post hoc Bonferroni analysis. (B) Track plot, (i) control, (ii) 5 mg/kg/day, (iii) 50 mg/kg/day.
In the subsequent study, the inhibition of NMDA receptor subunits expression which is the GluN2A and GluN2B in the adolescent male rat hippocampus, significantly inherited by the expression of GluN2A and GluN2B in foetus male rat hippocampus. The present findings were consistent with the previous study where prenatal BPA exposure at 0.05, 0.5, 5, 50 and 200 mg/kg/day inhibit the level of GluN2A and GluN2B in the hippocampus of male rat offsprings [15, 29]. In the NMDA receptor, these subunits consist of a glycine-binding site which contributes to Ca2+ permeability into the postsynaptic terminal [15]. The involvement of BPA in modulating these subunits may influence the threshold of LTP thereby impairing the cellular basis in learning and memory retention of an individual [30 –33].
Furthermore, prenatal BPA exposure at 5 and 50 mg/kg/day also decreased the level of synapsin I and PSD-95 in adolescent male rat hippocampus. Synapsin I is stably expressed in adult presynaptic terminals and its phosphorylation is important in regulating the synaptic vesicle release of neurotransmitters and synaptic formation [19]. The development of PSD-95 is also prominent in the adult stage, where it binds to the Ca2+/Calmodulin-dependent protein kinase II (CaMKII). It results in phosphorylation of GluN2B which enhances the NMDA receptors’ signalling [34]. These findings logically related to the controlled study of prenatal BPA exposure were at lower doses than the present study (0.04, 0.4 and 4 mg/kg/day), revealed to down-regulate the expression of synapsin I in the adult rodent’s hippocampus [14, 35]. A study by Wang et al. also showed that prenatal BPA exposure at 0.05 to 50 mg/kg/day reduced the expression of PSD-95 in adult rat hippocampus [36].
To determine the effect of prenatal exposure to BPA on cognitive behaviours, the rat will first validate the exploratory and emotional state by an open field test. In the open field test, the rats were released to explore by following their natural tendency to travel in the field. The analysis of anxiety-related behaviour in dim light showed BPA treated groups (5 and 50 mg/kg/day) increased the total distance travelled and several points of entry peripheral area. The results might consider positive atypical behaviour due to anxiety. Memory impairment might be caused the rat to recognise the surroundings and increase their exploratory activity, where the outcome has been observed in rodents with Alzheimer’s disease (AD) and familial frontotemporal dementia (FTDP-17) [37].
Conversely, the rat from BPA treated group has a lower number of crossing centre areas than the control group. It was found that the increase in the number of crossing centres inhibits anxiety-related behaviour in low light conditions [35]. Next, BPA-treated groups have a slightly higher frequency of freezing episodes than the control group, where increased time in immobility or resting episodes can indicate the level of anxiety or depressive state in rodent behaviour [38]. Lastly, higher emotional response of the rats reflecting anxiety at 5 and 50 mg/kg/day groups showed there is an increment in the number of grooming and number of faecal boli, which is consistent with the previous anxiety-related studies [14, 35, 39]. Hence, the current study demonstrates that prenatal BPA exposure regulates anxiety-related behaviour in adolescent male Sprague Dawley rats.
A test to evaluate fear or aversive memory was conducted using a step-down passive avoidance test to determine the state-dependent learning of the subjected animal. We found that the early exposure to BPA at 5 mg/kg/day impaired the passive avoidance memory in adult male rats from day 1 to day 6. However, the 50 mg/kg/day group showed no difference in latency to stay on the vibrating platform during the training session, yet a decrease in latency during the test session (day 6). Similarly, several BPA studies on adult male rats also showed that BPA exposure at 0.05, 0.4, 0.5, 4, 5 and 50 mg/kg/day decreased the time taken to step down from the platform than the control group [14, 15]. Next, the Morris water maze is an approach model which most widely accepted and used to evaluate hippocampal-dependent spatial learning and memory in rodents [40]. The result showed that BPA at 5 and 50 mg/kg/day impaired spatial memory by taking more extended time and distance to find the platform at the centre area of the pool. The studies by Zhang et al. [20], Hass et al. [21], and Liu et al. [41], also demonstrated that male rodents were receiving BPA (0.025, 0.04, 0.15, 0.25, 4, 5, 7.50, 40 and 50 mg/kg/day) exhibit the spatial memory deficits in Morris water maze test. Overall, it has been believed that the resulting memory impairment indicates that BPA can disrupt dendritic development and neurotransmitter homeostasis in the hippocampus [20]. The error in navigation ability is related to the decrement of the NMDA receptor subunits in the hippocampus of the rodents [15, 16, 42].
5 Conclusion
In conclusion, the administration of prenatal BPA exposure at 5 mg/kg/day (NOAEL) and 50 mg/kg/day (LOAEL) impaired learning and memory in an adolescent. The inhibition of NMDA receptor subunits (GluN2A and GluN2B) and synaptic markers (synapsin I and PSD-95) in the hippocampus during the foetus stage play a critical role in the synaptic development as their expression is fate to be inherited when reaching the adolescent. The findings suggested that the risk assessment of BPA exposure at NOAEL and LOAEL should be revised, especially for pregnant women, as the toxicity of BPA can influence the neurological function of the offspring as early as the foetus stage.
Footnotes
Ethical approval
This work is approved by the Committee on Animal Research and Ethics (UiTM CARE) under ethical approval number UiTM CARE: 351/2021 (3 Sept 2021).
Conflict of interests
All contributing authors have no conflict of interest in this work.
Funding
This project was funded by the Fundamental Research Grant Scheme, Ministry of Higher Education, Malaysia (Grant No. FRGS/1/2018/ SKK08/UITM/02/9) and Geran Penyelidikan Khas, Universiti Teknologi MARA [Grant No. 600-RMC/GPK 5/3 (191/2020)].
Authors’ contribution
Norazirah Mat Nayan: Literature research, experimental studies, data acquisition, data analysis, statistical analysis, manuscript preparation, manuscript editing. Rosfaiizah Siran: Study concepts, study design, definition of intelectuall content, experimental studies, data acquisition, data analysis, statistical analysis, manuscript preparation, manuscript editing, manuscript review. Siti Hamimah Sheikh Abd Kadir: Study concepts, study design, data analysis, manuscript editing, manuscript review. Musalmah Mazlan: Study concepts, study design, data analysis, manuscript editing, manuscript review. Andrean Husin: Study concepts, study design, data acquisition, manuscript editing, manuscript review. Che Badariah Abd Aziz: Study concepts, study design, data acquisition, manuscript editing, manuscript review.