
Abstract
Determining the spatial distribution of populations of Heilipus lauri and Stenoma catenifer, insects of economic and quarantine importance in avocado, is a useful information for designing and implementing integrated pest management plans. Aggregate spatial distribution is considered one of the most common patterns in nature, however, it has not been documented for these insect pest species. We propose through spatial statistical methods the analysis of data from periodic monitoring where the number of fruits with damage (FWD) caused by the two insect pests was recorded. The study was conducted between 2019-2020 in four commercial orchards of avocado cv. Hass in the department of Cauca, Colombia. A sampling design was established based on the recognition of damage on previously geopositioned trees. The spatial distribution was determined through Taylor's power law and Ripley's K function. Taylor's aggregation coefficient (b) was 1.47 and 1.22 for H. lauri and S. catenifer, respectively, indicating aggregation. Ripley's K function identified aggregation points of FWD in a radius range of 5-35 meters, with the aggregation sites being consistent across the two productive cycles. The damage was mapped indicating aggregation sites that will allow the implementation of management strategies for quarantine importance insect pests of avocado in specific sites of the crop, showing the precise location of the greatest concentration of damage and populations of H. lauri and S. catenifer.
Introducción
El aguacate Persea americana Mill. (Lauraceae), es considerado una de las frutas tropicales de mayor valor comercial e incremento del consumo mundial, resultado de su alto contenido nutricional, propiedades organolépticas y su potencial agroindustrial (Hurtado-Fernández et al., 2018; Montoya et al., 2018; Sandoval et al., 2010). En Colombia el sistema productivo de aguacate ha mostrado un crecimiento superior al 120% en la producción desde el 2015 hasta el 2021; representando un 65% del área sembrada, siendo el cultivar Hass el de mayor área con 34 % del total sembrada (Ministerio de Agricultura y Desarrollo Rural, 2021).
Entre las principales limitantes fitosanitarios que afectan la producción de aguacate con destino al consumo interno y exportación de fruta fresca, se destacan los insectos plaga de importancia económica y cuarentenaria; la polilla de la semilla del aguacate, Stenoma catenifer Walsingham (Lepidoptera: Depressariidae) y los picudos Heilipus lauri Bohemann y Heilipus trifasciatus (Fabricius) (Coleoptera: Curculionidade) (Carabalí Muñoz et al., 2021; ICA, 2016). En Colombia, la presencia y daño de H. trifasciatus se restringe a frutos de materiales nativos, en contraste, S. catenifer y H. lauri, se alimentan de la pulpa y la semilla en distintos genotipos y cultivares de P. americana; la biología de las dos especies está estrechamente relacionada con la disponibilidad de frutos de aguacate en diferentes estados fenológicos. Sobre pulpa y semillas de P. americana cultivar Hass el ciclo biológico de S. catenifer tiene una duración promedio de huevo a adulto de 61 días y sobre semillas de frutos del mismo cultivar, H. lauri tiene una duración promedio de huevo hasta la emergencia del adulto de 76 días (Carabalí-Muñoz et al., 2021; Diaz-Grisales et al., 2017). Por sus hábitos alimenticios y reproductivos, están ampliamente distribuidos en diferentes zonas productoras del país (Carabalí Muñoz et al., 2021); sin embargo, la fluctuación, presencia y daño de las poblaciones de S. catenifer y H. lauri, presenta variabilidad entre hospederos (genotipos, cultivares, variedades) en el espacio y tiempo.
La Corporación Colombiana de Investigación Agropecuaria (Agrosavia, Palmira, Valle del Cauca, Colombia) ha avanzado en estudios para el manejo de S. catenifer y H. lauri desde el año 2010, en el desarrollo y evaluación de métodos para monitorear la aparición de adultos de la polilla (trampas de luz) y el picudo, usando trampas tipo piramidal y lona piso (adultos presentes en ramas de aguacate) (Carabali-Muñoz, 2020). Sin embargo, los resultados mostraron que el método de monitoreo denominado “reconocimiento y seguimiento al daño en fruto” (método directo del reconocimiento de los síntomas de daño en frutos ocasionados por estados inmaduros de H. lauri y S. catenifer, que incluye también los daños de las hembras adultas de H. lauri durante la oviposición), se considera como el más fiable para la toma de decisiones en la implementación de estrategias de manejo de poblaciones (Carabali-Muñoz, 2020; Carabali-Muñoz et al., 2021). En este mismo sentido, los estudios también han revelado que en ciertas áreas específicas del cultivo el daño a frutos ocasionado por S. catenifer y H. lauri, se repitió en los ciclos productivos, demostrando así que las poblaciones de los dos barrenadores exhiben un patrón de distribución espacial agregado (Carabali-Muñoz et al., 2023), resultados que son consistentes con los reportados (Moral, 2004), quienes sustentan que las poblaciones de insectos se concentran en algunas áreas con mayor preferencia y continuidad en el tiempo; este comportamiento sugiere que las complejas interacciones que se establecen entre factores bióticos y abióticos influyen en la concentración de poblaciones de insectos y daños (Hassell et al., 1991; Schowalter, 2016). En este sentido, se ha determinado que las poblaciones de insectos en la búsqueda de satisfacer sus necesidades de alimentación y reproducción exhiben comportamientos propios de las especies que deriva en la generación de zonas de hábitats con características diferenciales (Roslin & Kotze, 2005).
Los avances en sistemas de información geográfica (GIS) y herramientas de análisis espacial como la geoestadística, han favorecido el desarrollo de investigaciones para determinar las distribuciones espaciales de especies con relación a su entorno físico y ambiental. Así mismo, la información y datos obtenidos pueden ser analizados y asociados con niveles de infestación, preferencias de agregación y la estructura de la población de insectos (Castrignano et al., 2012; Moral, 2004). El resultado de la aplicación de los métodos y análisis espaciales es una imagen o mapa que muestra la distribución de una población de insectos, permitiendo la identificación de “sitios calientes”; el conocimiento de estos sitios y variaciones de las poblaciones de insectos pueden ser estudiadas con el fin de identificar las áreas con mayor riesgo, optimizar el monitoreo de plagas y su control (Castrignano et al., 2012).
Esta ubicación en el espacio de las poblaciones de insectos y daño puede tener comportamientos de tipo agregado, uniforme o aleatorio, los cuales, pueden ser determinados por métodos de interpolación y visualizados en mapas. No obstante, el comportamiento de agregación de poblaciones de insectos puede ser determinado y calculado a través de diferentes modelos estadísticos como los basados en regresiones (Ley de potencia de Taylor e Iwao), u otros métodos dentro de la estadística espacial (Índice de Moran, función K de Ripley y semivariogramas) (Duarte et al., 2015; Nestel et al., 2004; Taylor, 2019; Vinatier et al., 2011). Diversas investigaciones han utilizado metodologías para identificar la distribución espacial de insectos plaga (Fortin et al., 2002; Rahbek, 2005). Por ejemplo, la distribución espacial de plagas como la chinche apestosa marrón Halymorpha halys Stål (Hemiptera: Pentatomidae) en durazno, plagas forestales no nativas en Norte América y poblaciones de plantas arvenses han utilizado técnicas como la Ley de potencia de Taylor, algunos índices de dispersión y la función K de Ripley (Hahn et al., 2017; Wang et al., 2019; Ward et al., 2019), combinando métodos cuantitativos, que relacionan la media y varianza, junto con la estadística espacial.
La distribución espacial de los insectos plagas de importancia cuarentenarias en los cultivos de aguacate no ha sido determinada y la aplicación de métodos cuantitativos en su identificación se constituye en una herramienta adicional que representa una etapa clave sobre la cual se pueden estructurar nuevos esquemas de monitoreo y manejo integrado de plagas (MIP) basados en los patrones de distribución del daño. El objetivo del presente trabajo fue caracterizar la distribución espacial y temporal de la afectación causada en frutos por H. lauri y S. catenifer en cultivos comerciales de aguacate cv. Hass en el departamento del Cauca, Colombia, mediante análisis de patrones puntuales combinando los métodos de Ley de potencia de Taylor y la función K de Ripley.
Materiales y métodos
Área de estudio, plantas y diseño de experimentos
La investigación se llevó a cabo en cuatro parcelas experimentales (PE) ubicadas en huertos comerciales de aguacate (P. americana Mill. cv. Hass) de aproximadamente cuatro años, establecidas en los municipios de Sotará y Timbío (Cauca, Colombia) (Tabla 1). Las parcelas se encuentran ubicadas en zonas de reconocida infestación y con registros de presencia de poblaciones y daño en frutos ocasionados por diferentes estados de desarrollo de S. catenifer y H. lauri. En cada una de las parcelas se tiene establecido un programa de manejo agronómico basado en podas, fertilizaciones periódicas provenientes de fuentes químicas y orgánicas. El manejo fitosanitario incluye la aplicación de fungicidas e insecticidas.
Características y ubicación de las parcelas experimentales evaluadas.
PE: Parcela experimental;
ha: Hectárea
Monitoreo de daños en frutos por Heilipus lauri y Stenoma catenifer
El método de monitoreo utilizado en los muestreos correspondió al método directo de reconocimiento del daño sobre frutos de aguacate cv. Hass ocasionado por poblaciones de H. lauri y S. catenifer. A pesar de que ambos insectos tienen entre sus hábitos a) barrenar el fruto, b) oviposición y alimentación asociada a la etapa fenológica de fructificación, c) preferencia de alimentación de pulpa y semilla y d) desarrollo de estados biológicos en el interior de fruto, estas especies presentan diferencias en la sintomatología de los daños resultado de las interacciones que establecen entre sus estados biológicos y su hospedero. En campo, el daño producido por H. lauri (Figura 1a) se reconoce en frutos entre tres y ocho meses de desarrollo, los cuales exhiben una perforación circular sin presencia de residuos de alimentación (preferencia de alimentación por semilla) (Figura 1b). En S. catenifer (Figura 1c) los daños son observables con mayor generalidad sobre frutos a partir de los tres meses de edad hasta la cosecha y a diferencia de H. lauri sobre el fruto se reconocen residuos de excretas, expulsadas por la larva joven producto de su preferencia de alimentación por la pulpa del fruto (Figura 1d).
El monitoreo de la población de adultos de H. lauri en campo se dificulta debido al comportamiento del insecto (tanatosis y preferencia de forrajeo en estratos medios y superiores del árbol) (Carabali et al., 2023). Sin embargo, se tiene ampliamente documentado que la fluctuación de su población y desarrollo biológico de los estados de huevo, larva, pupa y adulto, guardan una estrecha relación con la disponibilidad y estados fenológicos de frutos de P. americana cv. Hass (Carabali-Muñoz et al. 2021; Diaz et al., 2017). Este conocimiento permite validar la eficiencia del método directo y sustenta la aplicación de los frutos con daño como una unidad de muestreo consistente e informativa.
En H. lauri se realizaron observaciones periódicas de frutos para identificar los orificios circulares que realiza la hembra para depositar el huevo en un intento por ocultarlo y acercarlo a la semilla. La primera unidad de observación, está representada en una pequeña perforación oval en la epidermis del fruto (diámetro 4,4 ± 0,8 mm y profundidad de 2,0 ± 0,5 cm) (Carabali-Muñoz, 2020). Una segunda unidad de observación son los residuos de la excavación con los cuales la hembra cubre las horadaciones y el huevo. En ocasiones, las dos unidades de observación puede ser identificadas por la presencia de resina blanca o presencia de costra circular, resultado de la cristalización de los azúcares presentes en los exudados del fruto como respuesta al daño producido por el insecto (Caicedo et al., 2010).
En S. catenifer el método de monitoreo utilizado en los muestreos corresponde al reconocimiento del daño sobre frutos de aguacate cv. Hass que ocasionan los diferentes estadios larvales sobre la superficie y en el interior del fruto, donde las larvas en su recorrido hasta alcanzar la semilla consumen los tejidos del mesocarpio y endocarpio del fruto. Una primera unidad de observación la constituye una pequeña y casi imperceptible perforación (1,0 ± 0,2 mm) que realiza la larva de primer estadio. La segunda está asociada a la presencia de pequeños montículos de residuos de excretas (color café), que son depositadas por la larvas de estadios superiores al alimentarse de los tejidos internos del fruto (Carabalí-Muñoz et al., 2021). Al igual que en H. lauri, las unidades de muestreo de la polilla, en ocasiones pueden ser observadas por la presencia de resina blanca que las recubre.
Diseño de muestreo de frutos afectados por H. lauri y S. catenifer
Se definió un diseño de muestreo generalizado que incluía la totalidad de árboles y frutos de aguacate que conformaban cada una de las parcelas experimentales incluidas en el estudio (Tabla 1). Un total de 1676 árboles que se encontraban sembrados en las cuatro parcelas fueron marcados y geoposionados utilizando un GPS (por sus siglas en inglés) Garmin Map 62Sc (Garmin International, Inc., Olathe, Kansas, USA). El muestreo consistió en rodear el perímetro del árbol, revisar todos los frutos, seleccionar y registrar el número de frutos que presentaban daños causados por H. lauri y S. catenifer. La frecuencia del muestreo fue cada dos semanas durante dos ciclos productivos, iniciando en el estado fenológico 715 (50 % del volumen final del fruto) hasta el 719 (90 % del volumen final del fruto) (Alcaraz et al., 2013).
Los frutos que presentaban daños por H. lauri y S. catenifer se removieron y llevaron al laboratorio de entomología de AGROSAVIA (Palmira, Valle del Cauca, Colombia), donde se conservaron en tarrinas de plástico transparentes con dimensiones (35 x 25 x 13 cm) y papel toalla en el fondo. Las tarrinas fueron acondicionadas previamente para favorecer la aireación al interior del recipiente con un reemplazo del 80% de la superficie de la tapa por muselina blanca. Diariamente se revisaron los frutos hasta obtener los adultos. Para la confirmación de la especie se realizó un análisis comparativo con las poblaciones de las crías de H. lauri y S. catenifer establecidas desde 2015 en AGROSAVIA (C.I. Palmira) y descripciones morfológicas reportadas por Diaz et al. (2017) y Manrique et al. (2014).
Análisis de datos
La información registrada en las evaluaciones del comportamiento y distribución del daño ocasionado por H. lauri y S. catenifer en frutos de aguacate se realizó entre los ciclos de producción de los años 2019 y 2020. En total se efectuaron 48 monitoreos (seis/parcela/ciclo productivo) cada 15 días entre monitoreos por parcela en los estados fenológicos 715 (50% del volumen final) hasta el 719 (90% del volumen final) (Alcaraz et al., 2013). En los monitoreos se registró el número de frutos con daño por especie (H. lauri y S. catenifer) por árbol y se efectuaron análisis descriptivos de los daños entre las parcelas experimentales. Los datos se consideraron como Patrones Puntuales Espaciales (PPP) de naturaleza aleatoria para cada parcela experimental. Para el análisis estadístico de los patrones se utilizó el modelo de Poisson homogéneo, el cual asume un patrón teórico con Aleatoriedad Espacial Completa (CSR, por sus siglas en ingles) que permite determinar si la respuesta (patrón del daño) se identifica con un modelo de distribución aleatoria, uniforme o agregada en el espacio (Baddeley, 2008). Además, su utilización posibilita la visualización de los patrones en mapas, extrapolar las densidades de los puntos, y evaluar la dependencia espacial entre los puntos (Hahn et al., 2017; Ward et al., 2019).
En la determinación de la distribución del daño se utilizó la Ley de potencia de Taylor (Taylor, 1961). La ley de potencia Taylor (TPL por sus siglas en inglés) o ley de la media, es la relación entre la variancia observada (s2) y el promedio aritmético (m) mediante el uso de una regresión lineal para estimar dos parámetros: Log (s2) = log(a) + b.log (m). Donde el coeficiente b representa el índice de agregación y el tipo de distribución espacial (Taylor, 1961). La ley establece una distribución aleatoria (Poisson) cuando b = 1, uniforme cuando b<1 y b>1 indicará una distribución agregada (Taylor, 2019). Para este análisis se usó la función powers_law de la paquete de R epiphy en Rstudio (R Core Team, 2021).
La función K de Ripley se utiliza para evaluar la distribución espacial de un patrón, en este caso, la presencia de daño en los árboles causado por H. lauri y S. catenifer. Esta función compara las distancias entre pares de puntos del patrón observado con las que se esperarían en un patrón de aleatoriedad espacial completa (CSR).
El método de Ripley (Lefort et al., 2013; Ripley, 1981; Walter, 2006) se basa en el número de puntos “semis de puntos”. Estos pueden ser individuos, árboles en una población, o cualquier objeto contado a una cierta distancia o clase de distancia. La función K(r) se define como:

La función K de Ripley, compara el valor calculado de K(r) con el valor teórico esperado bajo CSR que es K(r) = (πr^2). Si los valores de K(r) son mayores que el valor teórico, indica una distribución espacial agregada del daño. Si son menores, sugiere una distribución uniforme, mientras que si son iguales, el patrón es aleatorio (Baddeley et al., 2015). K(r) se interpreta con la intensidad de población (λ) que hay por unidad de área λ = n/A (Mavunda et al., 2023).
Los gráficos de los valores K(r) fueron generados mediante la función Kest del paquete Spatstat en R. Para aumentar la confiabilidad de los resultados se aplicó la prueba de Monte Carlo que estima los intervalos de confianza. Solo se incluyeron en el análisis los monitoreos que reportaron un daño mínimo de 10 árboles, lo que permitió identificar posibles tendencias espaciales entre los puntos.
Resultados

Adultos de insectos plaga de importancia cuarentenaria y sintomatología externa de daño en frutos de P. americana cv. Hass. a. Adulto de H. lauri. b. Daño de H. lauri. c. Adulto de S. catenifer. d. Daño de S. catenifer.
Distribución temporal del daño de S. catenifer y H. lauri
En general, durante los dos ciclos productivos del 2019 y 2020 las poblaciones de H. lauri y S. catenifer, fueron identificadas como las especies que ocasionaron daños de manera generalizada en las parcelas experimentales. Sin embargo, produjeron variabilidad en los niveles de daño (Tabla 2). En el ciclo productivo del 2019 se registraron 1409 frutos afectados producidos por las dos especies, siendo 81 % mayor el daño por H. lauri comparado con S. catenifer. En este periodo, los mayores daños se presentaron en las parcelas I y II, aportando el 85,2% del daño total de las dos especies, siendo 25,36 % y 10,5%, superiores comparados con los daños registrados en la parcela III y IV, respectivamente (Tabla 2). Cuando se comparan los valores de número de frutos con daño (FCD) entre las parcelas I y II los resultados mostraron que las poblaciones de H. lauri, producen mayores pérdidas de frutos (77%), con relación a los daños que producen las poblaciones de S. catenifer. Los análisis del daño ocasionado por S. catenifer entre parcelas, muestran que la parcela I, presentó el mayor número de frutos afectados, siendo mayor 35%, 100% y 25,3%, comparados con las parcelas II, III y IV, respectivamente (Tabla 2).
Los análisis del ciclo productivo del año 2020 revelaron un incremento del 40,3 % en el número de frutos afectados por la polilla de semilla de aguacate S. catenifer con respecto al 2019. Entre parcelas el mayor daño ocasionado por H. lauri se presentó en la parcela II, siendo superior, 27,9 %, 25,7 % y 5%, con respecto a las parcelas, IV, III y I, respectivamente (Tabla 2). En el caso de las poblaciones de S. catenifer, el número de FCD de la parcela II, fueron 2 y 2,9 veces mayores, con respecto a las parcelas III y IV (Tabla 2). En el ciclo productivo del año 2020, un total de 1452 frutos presentaron síntomas de daño, siendo superior en 3%, comparado con el 2019.
En general la fluctuación de frutos con daño para el año 2019 en las cuatro parcelas para las dos especies mostró un comportamiento similar. Este se caracterizó por un mayor número de frutos afectados en las primeras etapas de evaluación y desarrollo del cultivo que fueron registrados en el primer monitoreo. Para H. lauri, se presentó un rango entre (50-370 frutos con daño) en el primer monitoreo y menor a 30 frutos con daño en los registros del último monitoreo. Así mismo, los análisis para S. catenifer mostraron en el primer monitoreo un rango de número de frutos afectados (0-180 frutos con daño) y sin presencia de frutos afectados en el registro del último monitoreo (Figura 2). En el año 2020, se registró un comportamiento similar de la fluctuación del daño entre monitoreos y parcelas (I, III, IV), sin embargo, un comportamiento diferencial fue registrado en la parcela II, donde las dos especies exhibieron un incremento del número de frutos afectados durante todos los monitoreos para H. lauri y entre el monitoreo 2 y 4 para S. catenifer (Figura 2). Los daños registrados en los tres primeros monitoreos en las cuatro parcelas experimentales se presentaron en los estados fenológicos del desarrollo del fruto de 715, 716 y 717 (correspondiente al 50 % del tamaño final del fruto y el 70% o más del tamaño final del fruto) que corresponden a los primeros tres (3) monitoreos (daños realizados en los meses de julio, agosto y septiembre). Estos resultados, permiten concluir que los programas de monitoreo deben estar dirigidos a estos estados fenológicos y validan como unidades de muestreo y observación a los frutos con daño que ocasionan las dos especies de insectos.
Daño en frutos de P. americana cv. Hass ocasionado por H. lauri y S. catenifer en cuatro parcelas experimentales durante dos periodos productivos.
FCD; frutos con daño.
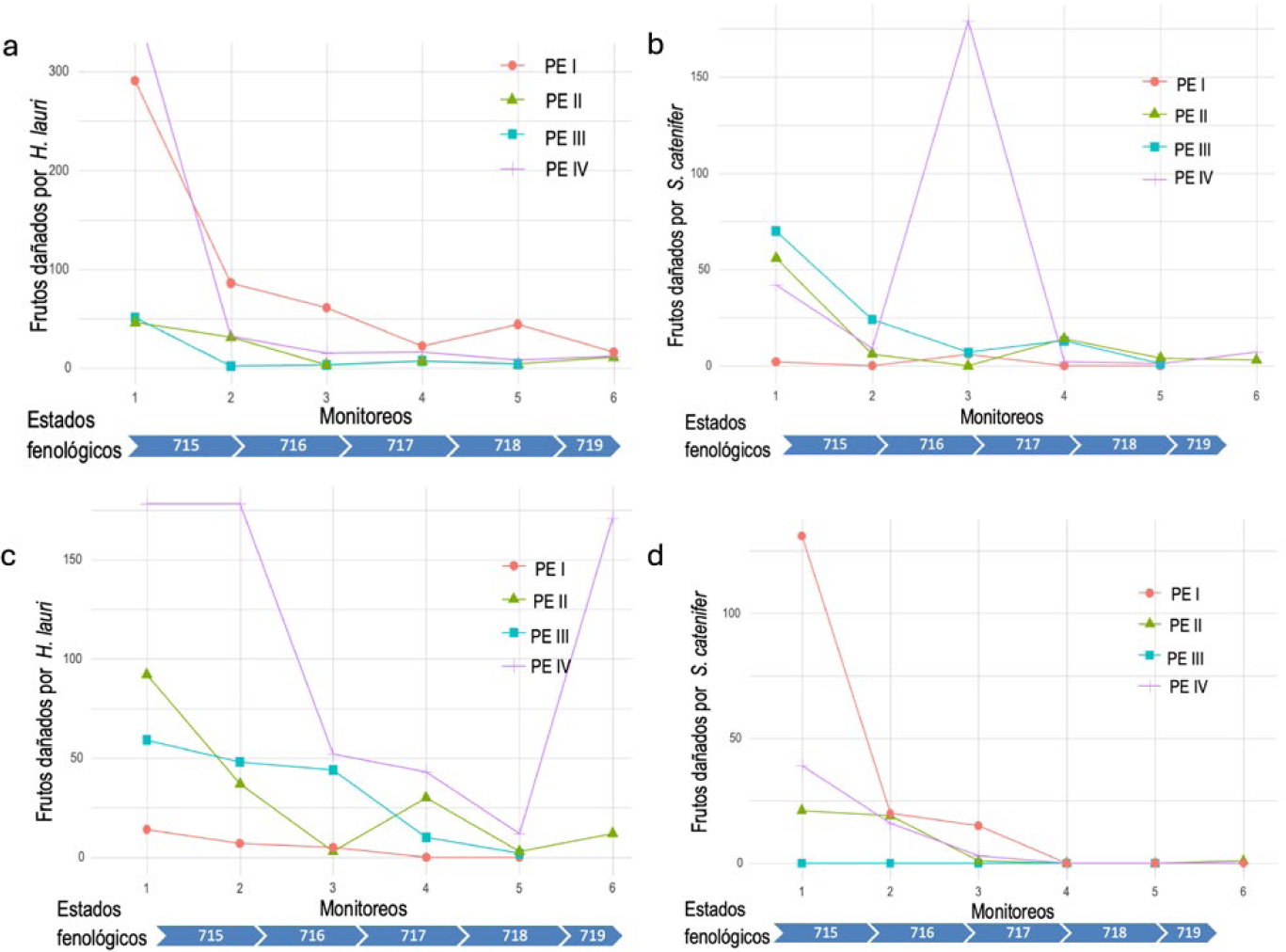
Daño en frutos por H. lauri y S. catenifer en los periodos 2019 y 2020. a. Daño en frutos por H. lauri monitoreos periodo 2019. b. Daño en frutos por S. catenifer monitoreos periodo 2019. c. Daño en frutos por H. lauri monitoreos periodo 2020. d. Daño en frutos por S. catenifer monitoreos periodo 2020.
Distribución del daño de H. lauri y S. catenifer mediante la ley de potencia de Taylor
Los resultados de los registros de frutos con menor daño obtenidos en los muestreos para cada especie de insecto-plaga por parcela (dos ciclos productivos) se utilizaron para ajustar los parámetros a considerar en la ley de potencia de Taylor. Los datos obtenidos en cada monitoreo forman un punto en el sistema de coordenadas. A la vez que se verifica la existencia de correlación linear entre los puntos de datos se identifica la relación entre la varianza observada [log (varianza observada)] de cada sub-muestra representada por la línea sólida y la varianza correspondiente a la presunción de distribución aleatoria [Loge (varianza binomial)] representada por la línea punteada (Figura 3).
Los análisis mostraron un valor del coeficiente de determinación (0,94 y 0,97) y el parámetro de agregación (b = 1,47 y 1,22) para los datos de monitoreos y registros del daño de H. lauri y S. catenifer, respectivamente. Los resultados obtenidos revelaron que se cumplen los supuestos de ley de potencia de Taylor, así mismo, revelan la tendencia de agregación de puntos correspondientes a los diferentes monitoreos y registros de frutos con daño ocasionados por S. catenifer y H. lauri (Figura 3). En los análisis se identificaron algunos periodos en los cuales no se observa la tendencia a la agregación dentro de las parcelas experimentales, sin embargo, cuando se realizan análisis conjuntos de datos se identifica una relación que se ajusta a una distribución estadística agregada. Los resultados encontrados son consistentes con los reportados por (Fortin et al., 2002; Rahbek, 2005), quienes utilizaron el método para identificar la distribución espacial de otra especie de insecto, a la vez que, permiten sugerir que su uso puede ser aplicado a la determinación del patrón de distribución de las poblaciones de H. lauri y S. catenifer. El patrón de distribución agregado que exhiben las poblaciones de insectos de importancia cuarentenaria en P. americana cv. Hass, será clave en la definición de los planes de monitoreo y programas de manejo.
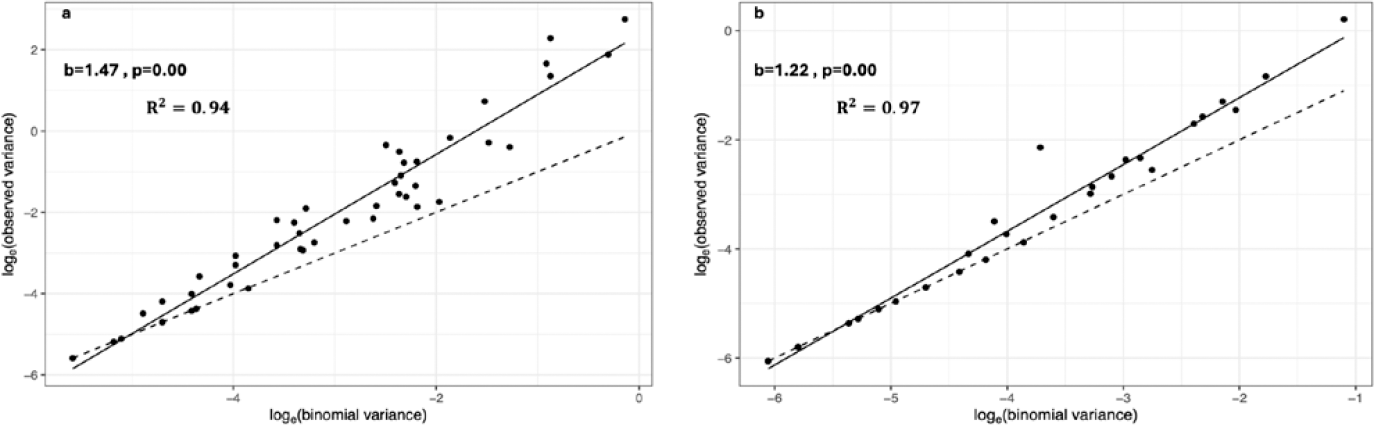
Ajuste de la Ley de potencia de Taylor de la distribución del daño causado por H. lauri y S. catenifer en los monitoreos de las parcelas I, II, III y IV para los periodos 2019 y 2020. a. Helipus lauri. b. Stenoma catenifer.
Función K de Ripley
El análisis de los resultados de los registros de los daños en los monitoreos cuando se utiliza la función K de Ripley permitió determinar la agregación, explicar el comportamiento y las distancias en las que ocurre la agregación del daño producido por las poblaciones de H. lauri y S. catenifer en las parcelas experimentales de aguacate. En la Figura 4 se presentan los resultados que se generan cuando se utiliza este método; la línea de color rojo (punteada) representa el patrón de Aleatoriedad Espacial Completa (CSR, teórico) y el área en gris representa los límites con un nivel de confianza (95%) simulados a partir de esta distribución teórica. Así mismo, la línea de color negro representa los datos observados y muestra el grado de agregación del daño a diferentes distancias. En general, los análisis revelaron que la agregación del daño se presentó a una distancia mínima de 0,00005 unidades de grados (r), que equivale a 5 m (conversión de arcos de grados a metros).
Los resultados de los análisis de la parcela experimental I (PE-I) mostraron la existencia de patrones de agregación del daño en frutos de aguacate ocasionado por H. lauri registrado en los monitoreos (M1, M2, M3 y M5) en el periodo 2019 (Figura 4a). Entre los monitoreos se presentó variabilidad, encontrándose, un mayor nivel de agregación en el monitoreo 1 (M1), comparado con el M2, M3 y M5. Entre los registros de los monitoreos 1 y 5 se presentó un descenso del nivel de agregación, resultado de la reducción del número de FCD, que disminuye entre monitoreos, por la implementación de la práctica de remoción de los frutos afectados. En el monitoreo 4 (M4), se observó una pérdida del nivel de agregación por la reducción del número de frutos con daño, seguido de un periodo de reinfestación en el M5, que desaparece en el M6 (Figura 4a). Los análisis del comportamiento del daño causado por S. catenifer, revelaron la presencia de agregación en el primer monitoreo (Figura 4b). A partir del monitoreo 2 (M2) y continuando hasta el M3, se registró una desaparición del nivel de agregación por la reducción de la incidencia del daño. Estos resultados sugieren que el daño en frutos de P. americana cv. Hass que producen las poblaciones de S. catenifer y H. lauri, mostraron mayor preferencia a alimentarse y desarrollarse en determinadas áreas, hechos que demuestran que la selección de árboles y frutos esta mediada por factores bióticos y abióticos que no fueron objetivo de este estudio.
Los resultados obtenidos con la prueba de K de Ripley revelaron que los daños producidos por poblaciones de H. lauri y S. catenifer durante el periodo 2019 ocurrieron en un área comprendida entre los 5 m y 35 m, de la parcela experimental I. Esto demuestra que la concentración o agregación del daño se presentó a una distancia (≤ 35 m) de radio con relación al sitio (árboles) que presentaron el mayor daño. En el periodo 2020 los análisis con K de Ripley de los registros del daño de la parcela I no permitieron identificar niveles de agregación por la baja incidencia de las dos especies plaga causado por la remoción de frutos con daño que se realizó durante el monitoreo.
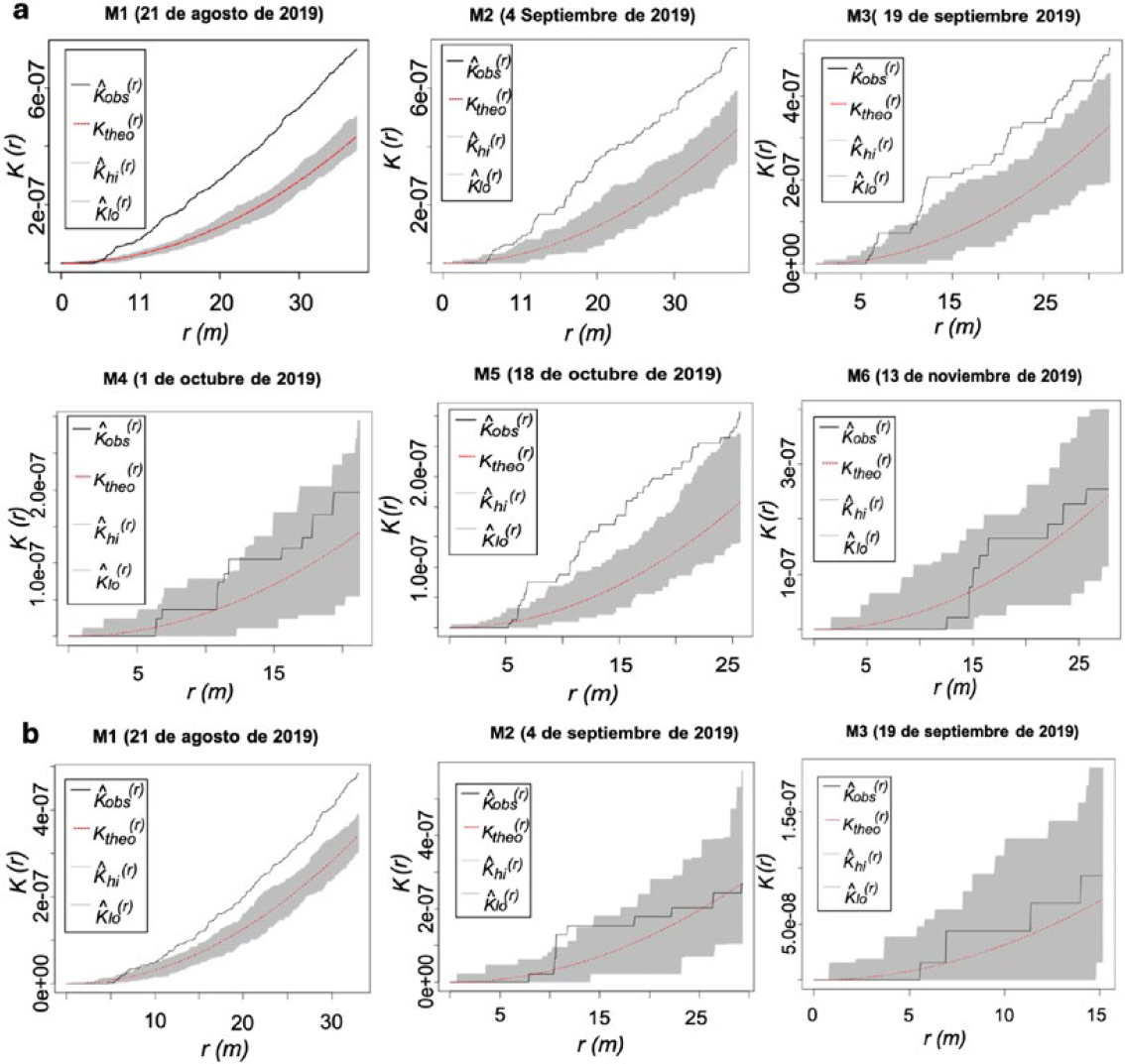
Parcela experimental (PE I): Función K de Ripley de la presencia de daño de plagas de aguacate en frutos de aguacate (Persea americana cv. Hass) para cada monitoreo en el 2019. a. H. lauri. b. S. catenifer
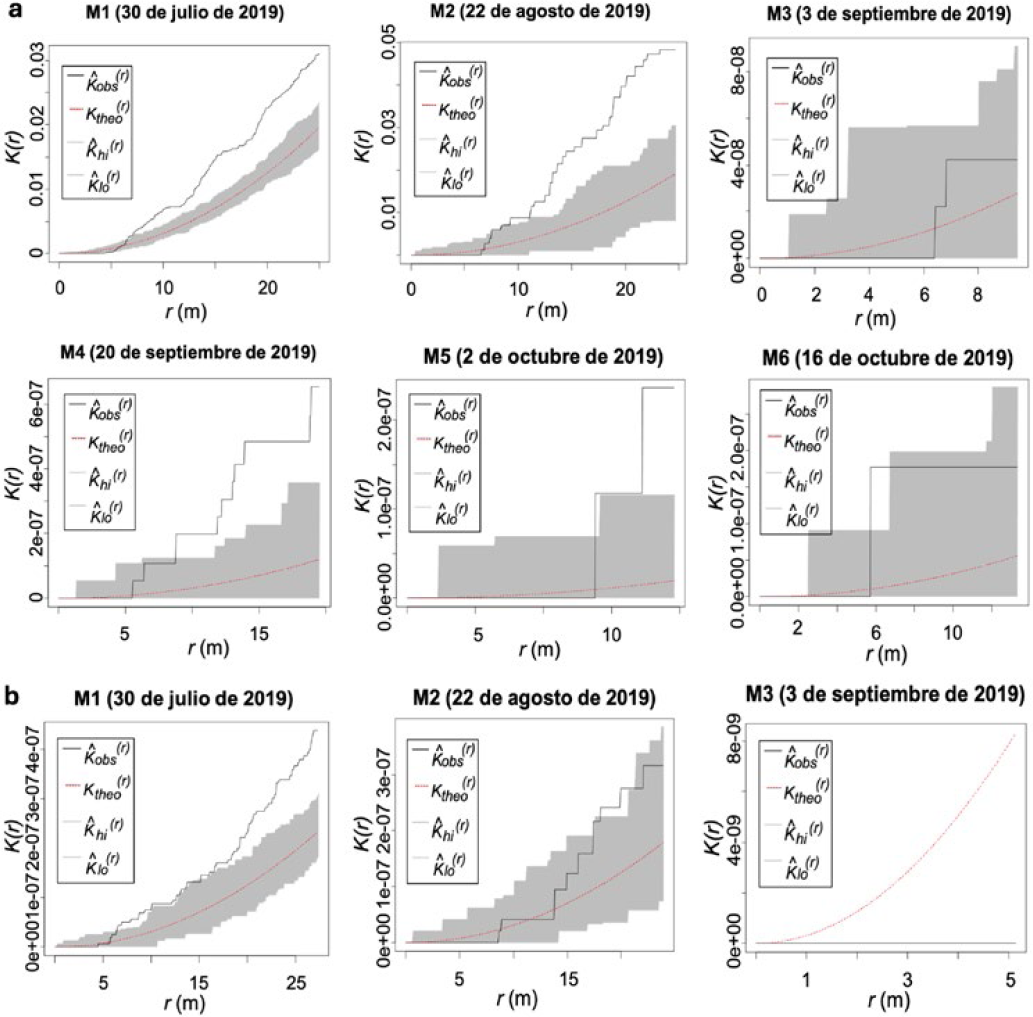
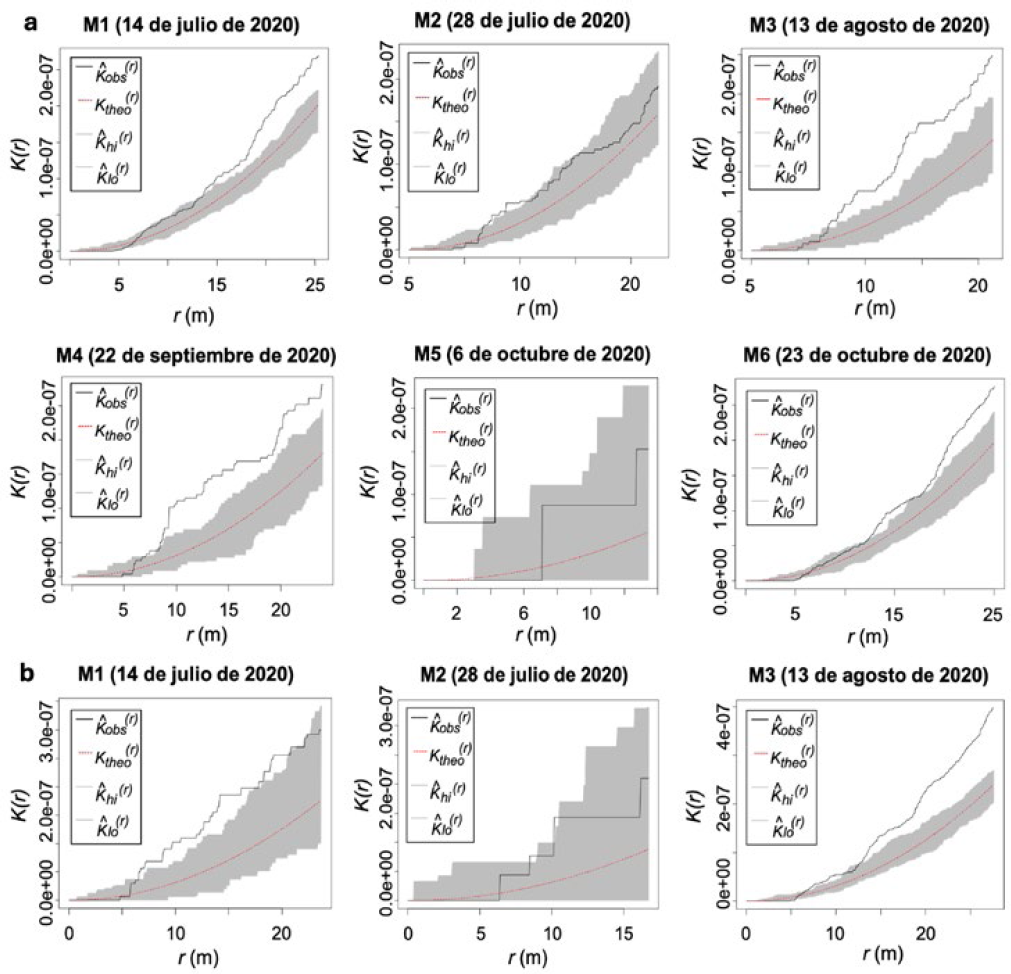
Los análisis de los registros (2019) de la parcela II, mostraron la presencia de patrones de agregación del daño en frutos ocasionado por H. lauri registrados en los monitoreos (M1, M2 y M4) (Figura 5a). Cuando los análisis se realizaron entre monitoreos se presentó variabilidad en los resultados. Se encontró una mayor agregación en el monitoreo 1 (M1) y monitoreo 2 (M2), comparado con M3, M4 y M6. Los resultados para S. catenifer, mostraron agregación en el registro del daño obtenidos en el monitoreo 1, sin embargo, los análisis efectuados con los registros de los monitoreos siguientes (M2 a M6), no presentaron una agregación sobresaliente debido a la disminución del número de frutos con daño (Figura 5b).
Los resultados de los análisis de la parcela II (año 2020) mostraron la presencia de agregación del daño ocasionado por H. lauri en los monitoreos (M1, M3, M4 y M6) siendo más pronunciada en los monitoreos 3 y 4 (Figura 6a). En los registros de los monitoreos 2 y 5 se presentó un menor nivel de agregación, resultado de la disminución de los frutos con síntomas de daño, debido a reducción de unidades de muestreo (frutos con daño) en el monitoreo 5 y la concentración del daño en un menor número de árboles de aguacate registrado para el monitoreo 2, obteniendo como resultado menor agregación definida por un mayor números de árboles con daño, no obstante, la agregación se da por la concentración del daño en pocos árboles que presentaron altos niveles de infestación (≥ 150 frutos afectados) (Figura 6a). En el monitoreo 6, los análisis muestran incrementos en los niveles de daño como consecuencia de reinfestaciones por incrementos de las poblaciones de insectos plaga que produjeron un aumento en el nivel de agregación del daño. En el periodo 2020 los resultados del comportamiento del daño causado por S. catenifer, mostraron patrones de agregación en los registros del monitoreo 1 (M1), posteriormente, en los monitoreos siguientes, se observa una reducción del daño y bajos niveles de agregación en los siguientes monitoreos con excepción del monitoreo 3 (M3), en el cual se presentó una reinfestación y agregación del daño (Figura 6b).
La función de K de Ripley reveló que la agregación espacial del daño producido por las poblaciones de H. lauri y S. catenifer en la PE II durante los años 2019 y 2020 se presentó a una distancia enmarcada en un radio entre los 5 m y 30 m, donde se encuentran los árboles registrados en los monitoreos como los de mayor número de frutos afectados que en conjunto representan las áreas en las que se concentró el mayor daño.
De acuerdo con el daño reportado (Figura 1) y la función K de Ripley se puede observar que hay una relación entre el número de frutos afectados y la agregación espacial. Cada monitoreo con daño mayor a 50 frutos presentó una marcada agregación de los frutos con síntomas de daño para ambas especies, con excepción del monitoreo 2 (M2) de la parcela II (periodo 2020) (Figura 6a), en el cual se encontraron 178 frutos con daño. Sin embargo, la agregación del daño no superó los intervalos de confianza debido a que el daño se concentró en un menor número de árboles y la función K de Ripley lo representa como un punto independientemente de la cantidad del daño asociado.
PE II: Función K de Ripley de la presencia del daño de H. lauri y S. catenifer en frutos de aguacate (Persea americana cv. Hass) para cada monitoreo en el 2019. a. Heilipus lauri. b. Stenoma catenifer.
Los resultados de los análisis para la parcela III mostraron la presencia de agregación espacial del daño en frutos de aguacate ocasionado por H. lauri en el monitoreo 1 (M1) (periodo 2019), a partir del cual la incidencia del daño se redujo, a causa de la remoción de los frutos con síntomas de daño (Figura 7). Para el caso de S. catenifer no se presentó daño para este periodo.
PE II: Función K de Ripley de la presencia del daño en frutos de aguacate (Persea americana cv. Hass) para cada monitoreo en el 2020. a. Heilipus lauri. b. Stenoma catenifer.
Los resultados de los análisis de los monitoreos en la parcela III (periodo 2020) presentaron agregación espacial del daño causado por H. lauri de manera significativa en el monitoreo 1(M1) (Figura 8a). En los siguientes monitoreos el daño se redujo de forma gradual, presentado niveles de agregación en el monitoreo 2 (M2), a una distancia entre 25 m a 30 m y en el monitoreo 3 (M3) a 15 m, desapareciendo el patrón de agregación para los siguientes monitoreos (Figura 8a). Para poblaciones de S. catenifer relacionadas con el daño en frutos de P. americana cv. Hass, los análisis mostraron agregación espacial del daño en la parcela III y monitoreo 1 (M1) a una distancia de los 5 m hasta los 35 m. En los siguientes monitoreos los niveles de daño se redujeron y como consecuencia no se presentó agregación (Figura 8b).
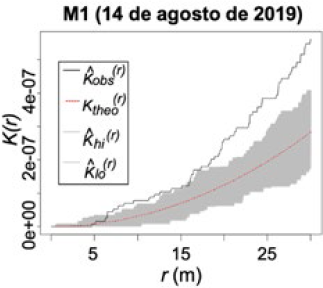
Parcela experimental III: Función K de Ripley de la presencia del daño de H. lauri en frutos de aguacate en los monitoreos del 2019.
Los resultados de los análisis de los registros de los monitoreos obtenidos en la parcela IV (periodo 2019) presentaron agregación del daño en frutos ocasionado por poblaciones de H. lauri en el monitoreo 1 (M1). A partir de los monitoreos siguientes los registros de número de frutos con daño fueron menores al igual que los niveles de agregación, siendo identificada una concentración del daño en el monitoreo 2 (M2) a una distancia (≥ 15 m y ≤ 20 m) (Figura 9a). Las poblaciones de S. catenifer produjeron un mayor daño agregado en el monitoreo 1 (M1) identificado a una distancia entre (10 m y 25 m). Los análisis de los monitoreos siguientes no mostraron niveles de agregación (Figura 9b). Los análisis del periodo 2020 mostraron concentración del daño en frutos, ocasionado por las dos especies en los registros obtenidos en los monitoreos 1 (M1), 2 (M2) y 4 (M4) que presentaron un nivel de agregación similar (Figura 10a). Las poblaciones de S. catenifer, produjeron menores niveles de daño y no hubo agregación significativa entre los monitoreos donde se registró daño (Figura 10b).
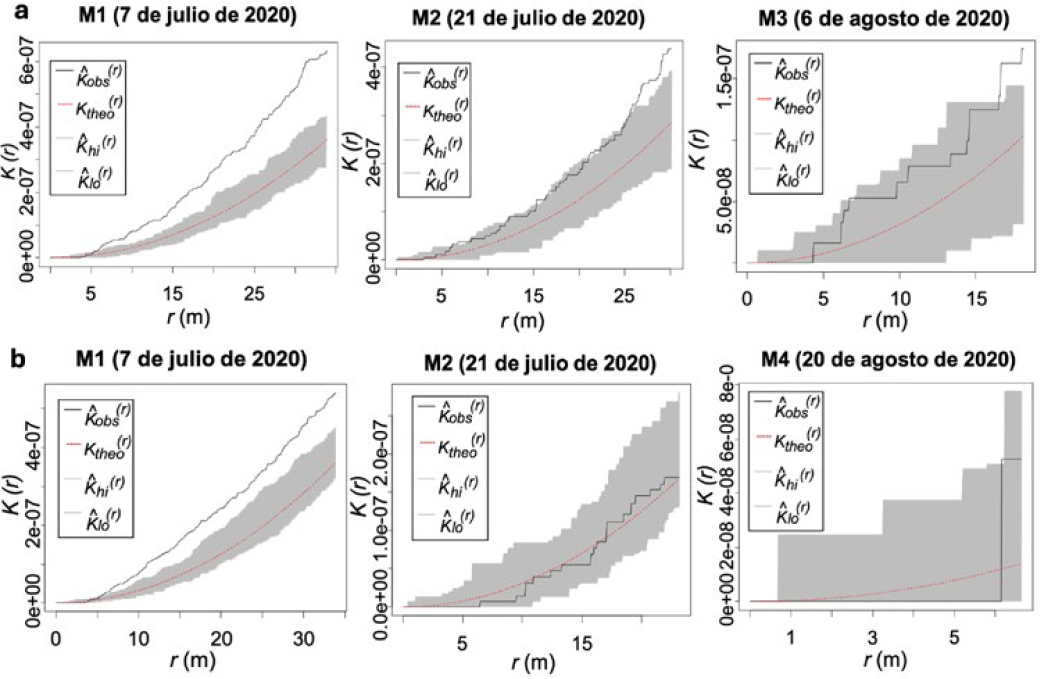
Parcela experimental III: Función K de Ripley de la presencia del daño de H. lauri y S. catenifer en frutos de aguacate (Persea americana cv. Hass) monitoreos periodo 2020. a. Heilipus lauri. b. Stenoma catenifer.
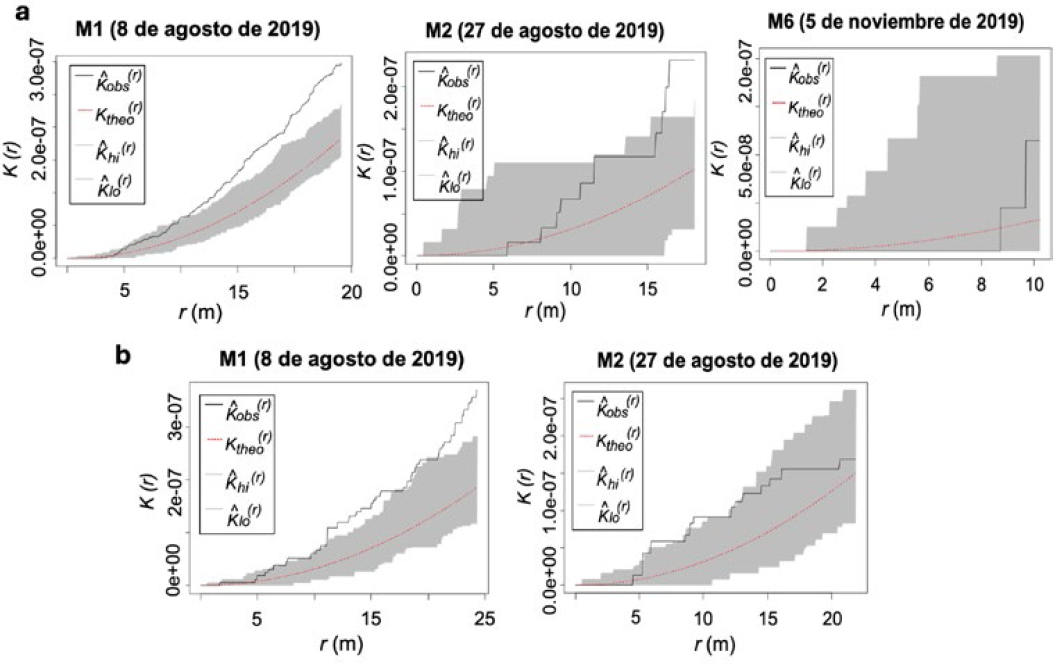
PE IV: Función K de Ripley de la presencia del daño de H. lauri y S. catenifer en frutos de aguacate (Persea americana cv. Hass) monitoreos periodo 2019. a. Heilipus lauri. b. Stenoma catenifer.
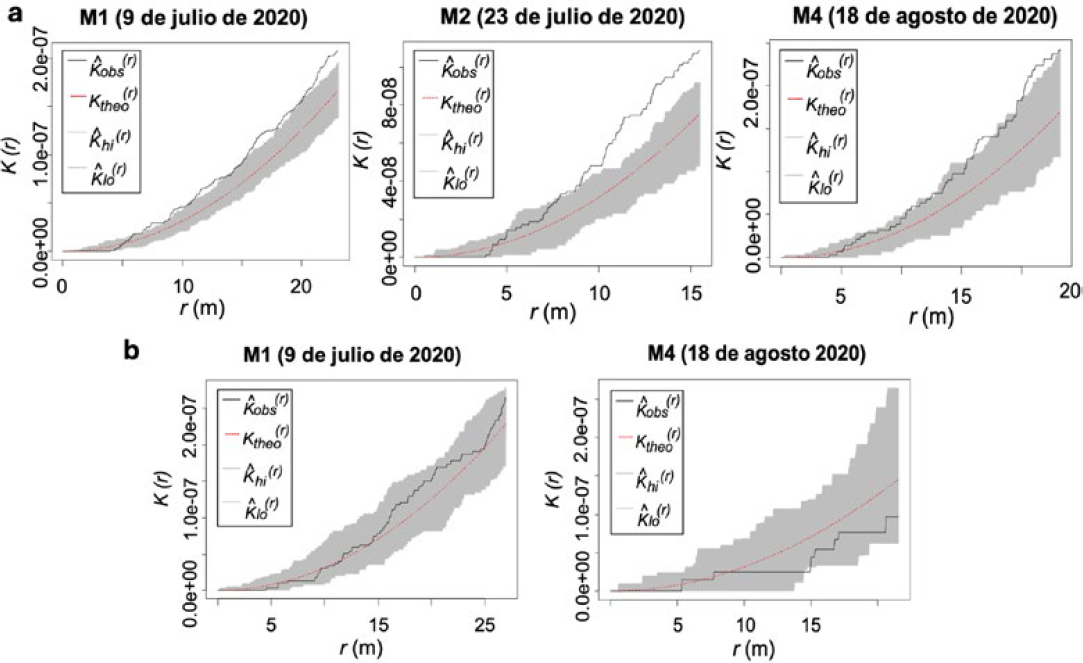
PE IV: Función K de Ripley de la presencia del daño de H. lauri y S. catenifer en frutos de aguacate (Persea americana cv. Hass) en los monitoreos del 2020. a. Heilipus lauri. b. Stenoma catenifer.
Estimación de densidad de Kernel (KDE)
Los análisis a través del método de interpolación de Kernel (KDE) representan una forma alternativa de visualizar en mapas el comportamiento de agregación de la variable frutos con daño. A partir de los árboles georreferenciados en las parcelas experimentales se encontraron puntos (sitios) de agregación del daño en frutos de P. americana cv. Hass, causado por S. catenifer y H. lauri. Los resultados encontrados con el método de Kernel son consistentes con la tendencia a la agregación registrada en los monitoreos iniciales identificados con los métodos K-Ripley (Figura 3-9). Adicionalmente, con la aplicación del método de Kernel, se logran identificar en el espacio los sitios (zonas del cultivo) donde se presenta la concentración de árboles con mayor número de frutos afectados en las diferentes parcelas evaluadas y su comportamiento en el tiempo.
Algunas de las zonas con agregación identificadas en las cuatro parcelas persistieron en el tiempo y en la mayoría de las ocasiones se repitieron en el siguiente ciclo productivo. En este mismo sentido, los resultados de la distribución espacial del daño encontrados en la parcela I (periodo 2019), muestran, que la concentración de los frutos afectados por S. catenifer y H. lauri, fue la misma para las dos especies, lo que demuestra similitud en la preferencia de árboles como sustrato de alimentación y oviposición. En general, se presentaron dos sitios con agregación “focos” ubicados en la zona norte y sur de la parcela I, los cuales se registraron con mayor intensidad durante los dos primeros monitoreos (M1 y M2) con un registro de daño (≤ 25 FCD por árbol), a partir de los cuales se redujo la intensidad del daño y sitios de agregación “focos” que coincide con los resultados encontrados cuando los datos se analizaron con la función K de Ripley (Figura 11).
Los resultados de la distribución espacial del daño en la parcela II (2019), revelaron la presencia de dos sitios con daño (focos) ubicados en la zona norte y sur de la parcela (cercanos al borde). Los sitios de agregación coinciden con los registrados en ciclos productivos anteriores. El daño ocasionado por H. lauri presentó variabilidad en la distribución e intensidad a través del tiempo. En el monitoreo 1 (M1) se presentó un foco principal en la zona norte y otro con ubicación noroeste de la parcela II, con registro de daño (≥ 50 FCD por árbol) (Figura 12a). Para S. catenifer en el monitoreo 1 (M1) se registró un foco en la ubicación noreste y otros daños menores distribuidos en la zona centro-sur de la parcela II con un máximo de seis frutos afectados por árbol. En los monitoreos iniciales (M1-M2) fue donde se lograron registrar las principales agregaciones obtenidas con los métodos de los análisis de agregación descritos anteriormente (Figura 12b).
La distribución espacial del daño producido por poblaciones de H. lauri en el periodo 2020 (parcela II), muestra la ubicación de un sitio con mayor agregación en la zona central con incidencia de 30 frutos con daño por árbol; el cual presentó un incremento registrado para el monitoreo 2 (M2) presentando hasta 60 frutos con daño por árbol (Figura 13a). Posteriormente, los análisis revelan una reducción en la agregación del daño. Los frutos con daño producidos por S. catenifer se presentaron principalmente en la zona norte registrado en el monitoreo 1 (M1) y en el sector sur para el monitoreo 2 (M2) (incidencia: ≤ 6 FCD por árbol), coincidiendo los focos de daño en las mismas zonas durante los periodos 2019 y 2020 (Figura 13b).
Los mapas obtenidos de las interpolaciones para la parcela III (periodo 2019), mostraron que el daño de H. lauri se registró específicamente en el monitoreo 1 (M1), con un sitio de agregación (foco) en la zona suroeste (incidencia: ≤7 FCD por árbol) y sin presencia de daño ocasionado por S. catenifer. En el 2020, la distribución espacial del daño presentó una mayor dispersión en la parcela III con distribuciones agregadas similares de frutos con daño causado por H. lauri y S. catenifer. En este periodo se presentó un surgimiento de focos en el monitoreo 4 (M4) (incidencia: ≤ 7 FCD por árbol) y monitoreo 3 (M3) (incidencia: ≤ 16 FCD por árbol) para H. lauri y S. catenifer, respectivamente, coexistiendo poblaciones y daño en el mismo sitio (Figura 14).
Los resultados de la distribución espacial del daño en la parcela IV (2019), muestran un sitio de agregación principal en la zona norte causado por poblaciones de H. lauri (Figura15a) que alcanzaron mayores niveles de infestación durante los periodos correspondientes a los monitoreos 1 y 2 (M1 y M2) con una incidencia (≤ 10 frutos afectados por árbol). Por el contrario, en los monitoreos 3, 4 y 5 (M3, M4 y M5) no hay presencia de daño, reapareciendo nuevamente en los registros del monitoreo 6 (M6) los síntomas de daño producido H. lauri, que coincide con los sitios donde anteriormente se registró el daño (M1 y M2) (Figura 15a). Los resultados de la parcela IV (2020) mostraron un foco principal del daño ocasionado por H. lauri y S. catenifer en el borde este de la parcela correspondiente a los registros obtenidos en el monitoreo 1 (M1) (incidencia: ≤ 7 FCD por árbol) y otro foco con menor daño en la zona oeste. Estos sitios de concentración identificados en los M1, M2, reaparecieron en el monitoreo 6 (M6) en la zona oeste para H. lauri y en el monitoreo 4 (M4) en la zona este para S. catenifer (Figura 16).
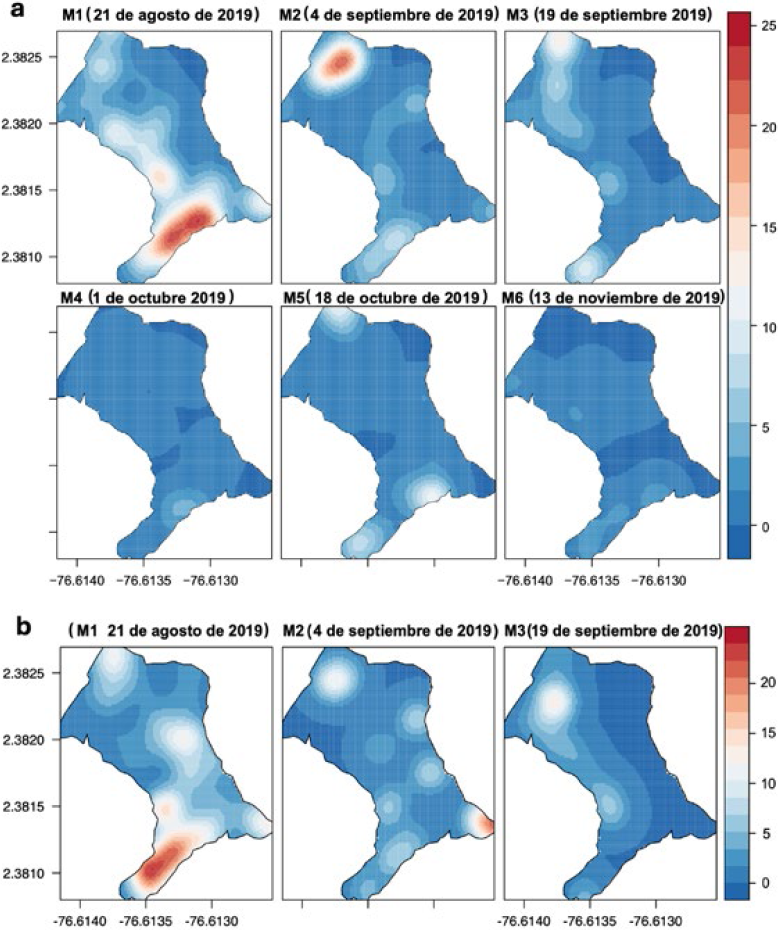
PE I: Interpolación KDE de la distribución espacial de FCD de H. lauri y S. catenifer para monitoreos año 2019. a. Heilipus lauri. b. Stenoma catenifer. En las Figuras a y b, las áreas diferenciadas en color rojo muestran la concentración del daño ocasionado por las dos especies.
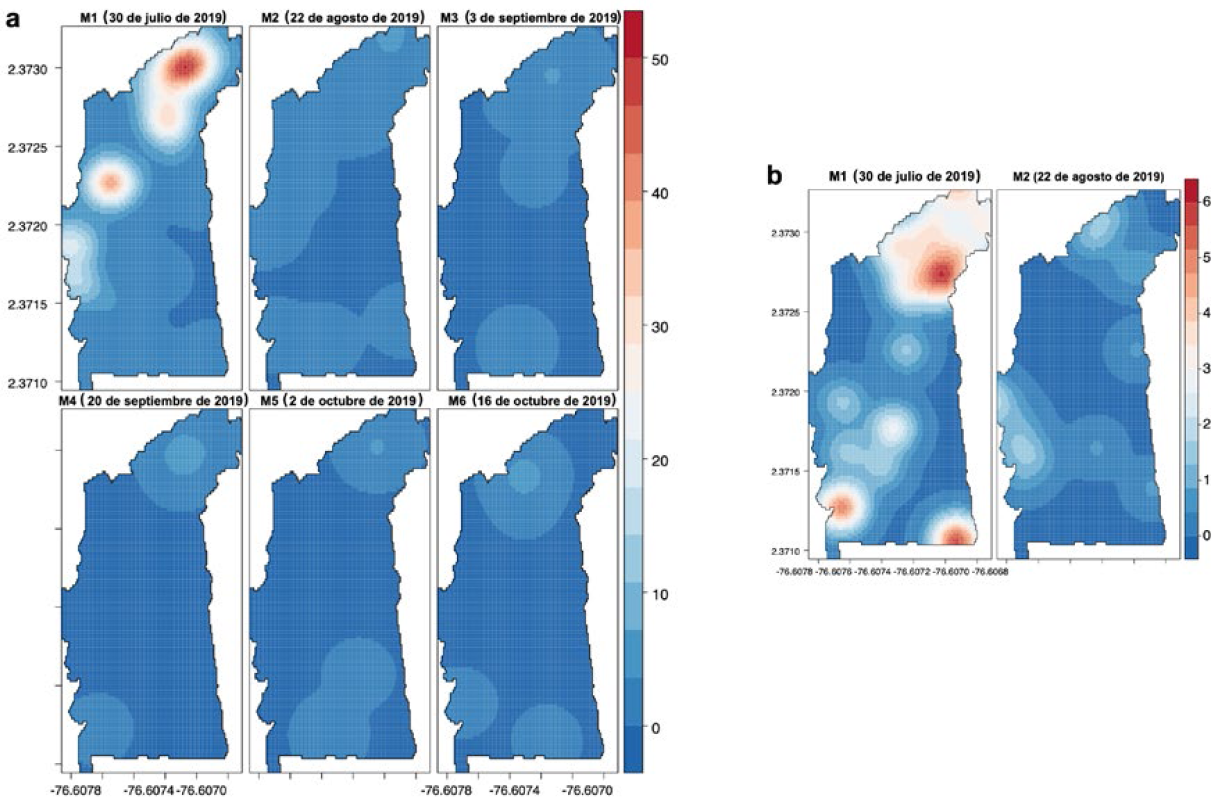
PE II: Interpolación KDE de la distribución espacial de FCD de H. lauri y S. catenifer monitoreos año 2019. a. Heilipus lauri. b. Stenoma catenifer.
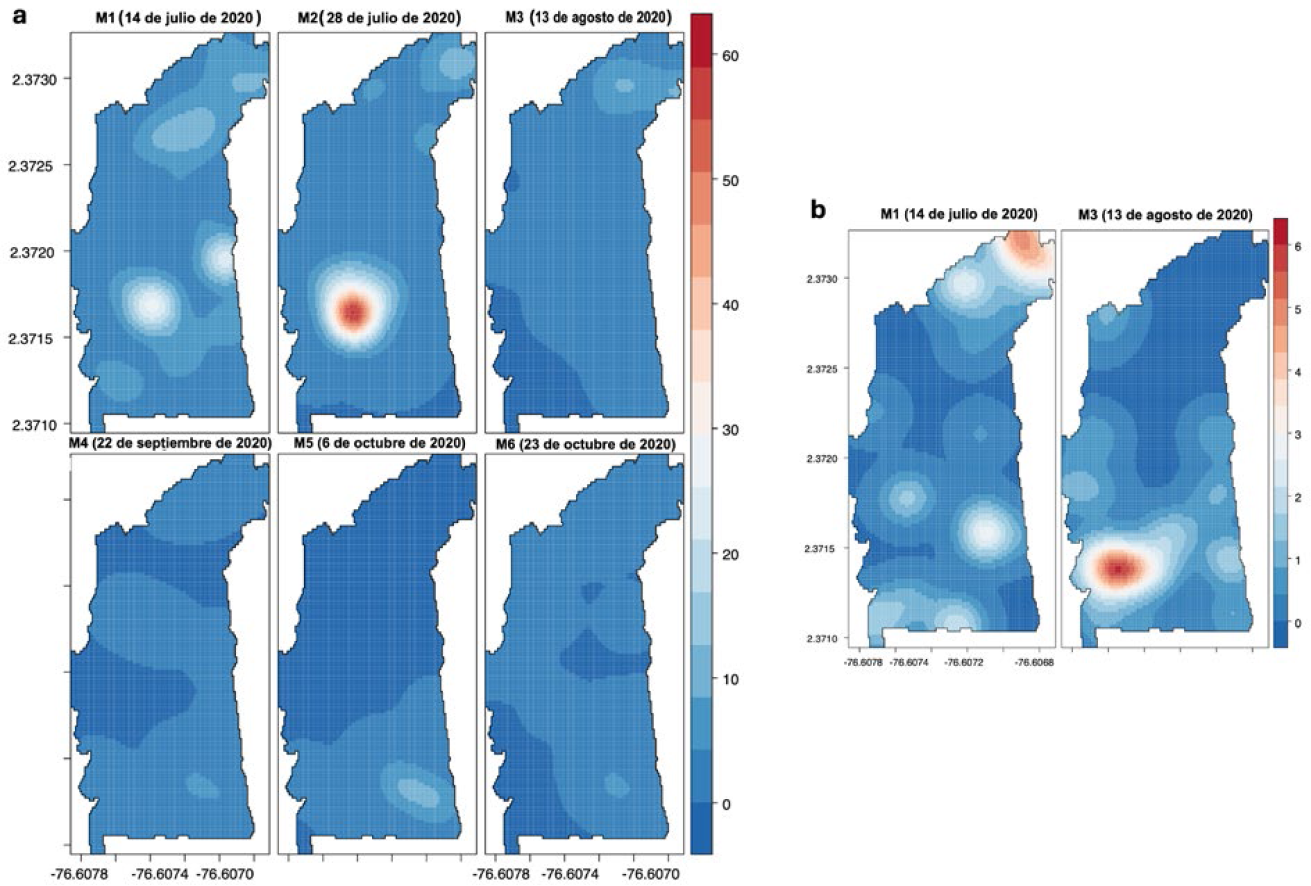
PE II: Interpolación KDE de la de la distribución espacial de FCD de H. lauri y S. catenifer en monitoreos año 2020. a. Heilipus lauri. b. Stenoma catenifer.
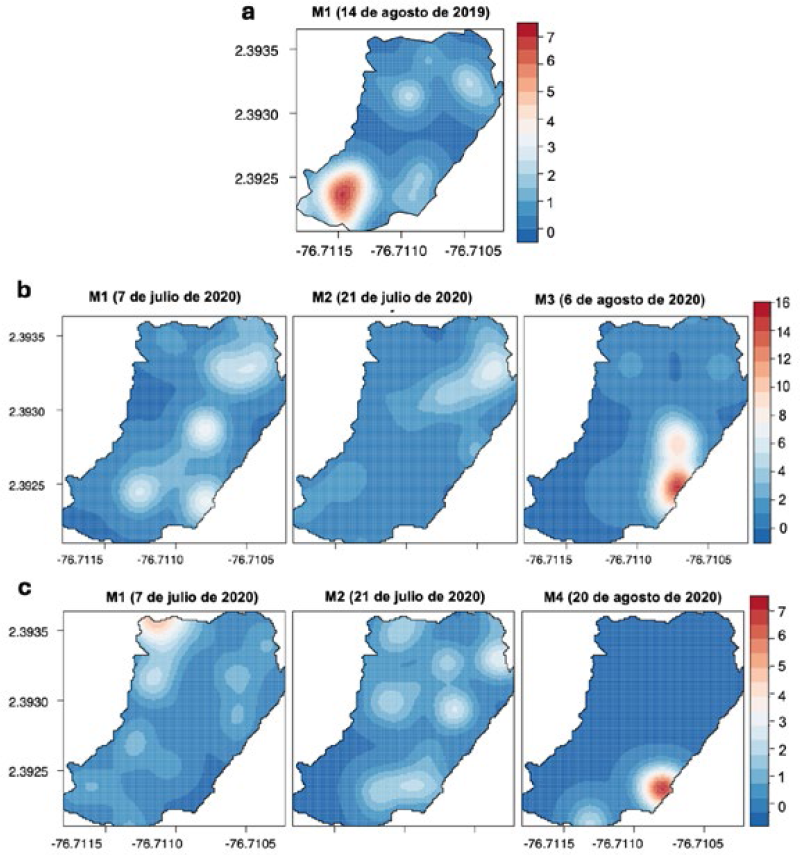
PE III: Interpolación KDE de la distribución espacial de FCD de H. lauri para los monitoreos durante 2019 y 2020 y de S. catenifer para el 2020. a. Heilipus lauri año 2019. b. H. lauri año 2020. c. Stenoma catenifer año 2020.
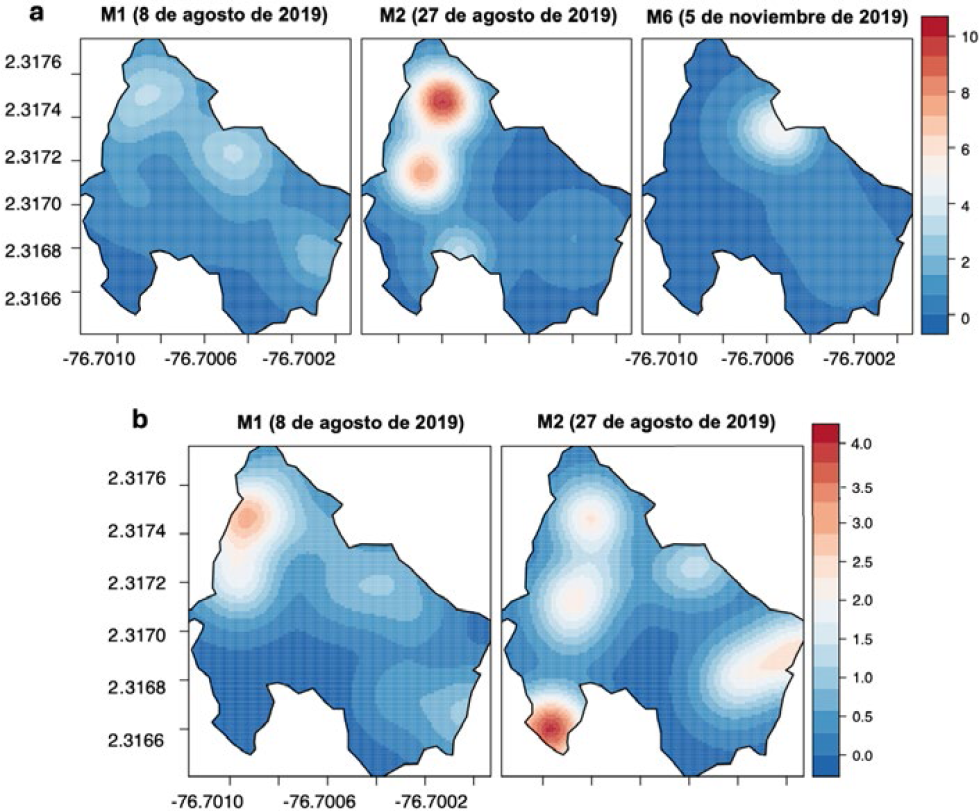
PE IV: Interpolación KDE de la distribución espacial de FCD de H. lauri y S. catenifer en monitoreos del 2019. a. Heilipus lauri. b. Stenoma catenifer.
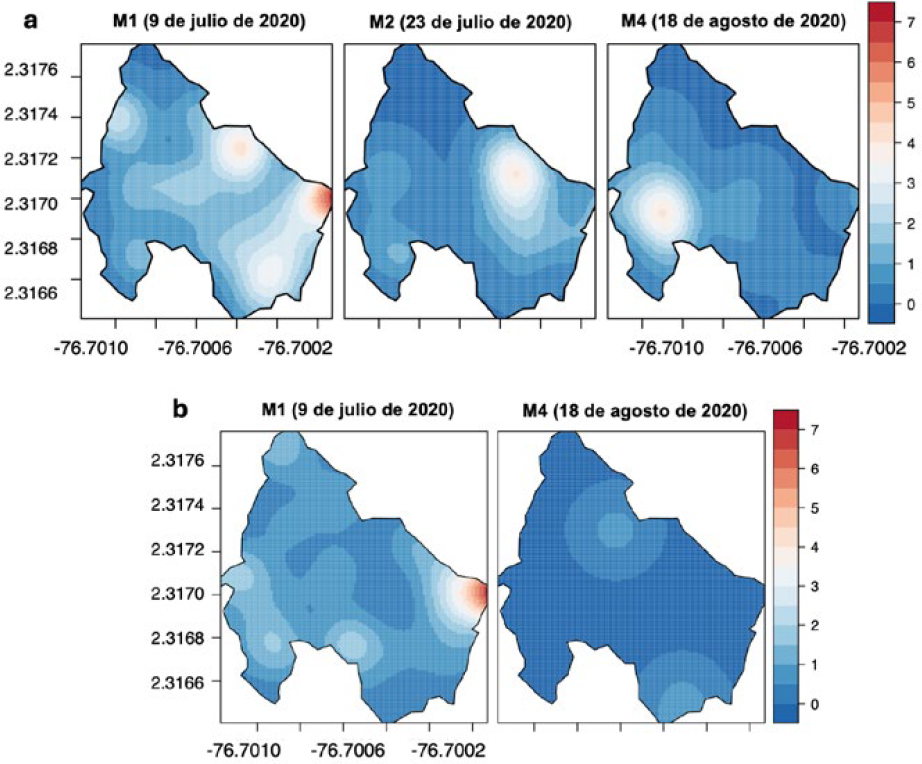
PE IV: Interpolación KDE de la distribución espacial de FCD de H. lauri y S. catenifer en monitoreos del 2020. a. Heilipus lauri. b. Stenoma catenifer.
De manera general se observa que el daño presentó una agregación principalmente en los registros obtenidos en el primer monitoreo (M1), a partir de los cuales se presentaba una reducción en la mayoría de los casos, debido a la remoción del árbol de los frutos con daño que se implementa durante el plan de monitoreo y muestreos.
Discusión
Los resultados del estudio revelan la presencia de patrones espaciales agregados en la distribución del daño causado por S. catenifer y H. lauri en las parcelas y cultivos de P. americana aguacate Hass. Estos patrones, identificados mediante métodos estadísticos y geoestadísticos, sugieren una correlación significativa en la concentración de daño en ciertas zonas específicas que fueron recurrentes en los diferentes periodos de evaluación.
Las parcelas experimentales se encontraban separadas por distancias de más de 10 km entre ellas, con la excepción de las parcelas establecidas en el municipio de Sotará, que estaban a 1 km de distancia. Además, a pesar de que las parcelas de estudio eran menores a 2 ha, se ubicaban en zonas aguacateras amplias, lo que las hace extrapolables a áreas más grandes debido a la amplia distribución de las plagas y sus preferencias a presentarse por focos.
Los insectos son organismos ectodermos, con temperaturas corporales variables y dependen de las condiciones del ambiente para su desarrollo (Gilbert & Raworth, 1996). Según los resultados del estudio, la distribución espacial agregada que exhibe el daño esta posiblemente relacionada con las diferenciadas condiciones ambientales y estados fenológicos que presentan los sitios preferidos por las poblaciones de insectos para alimentarse y reproducirse (Dawar et al., 2022). A pesar, que el presente estudio no tenía como propósito determinar los factores que influyen en la selección de los sitios por las dos especies de insectos, resultados de evaluaciones han mostrado que las variables ambientales (temperatura, humedad relativa, dirección del viento), cuando se conjugan e interrelacionan, establecen diferenciales condiciones ambientales en los sitios de agregación, que representan áreas (focos) donde las tasas intrínsecas de crecimiento de las poblaciones de S. catenifer y H. lauri, se favorecen (Carabali-Muñoz et al., 2023). Adicionalmente, este comportamiento y la selección de sitios se presentó con mayor regularidad en áreas cercanas a los bordes de cultivos que representan microhábitats donde predominan árboles de aguacates “nativos” que funcionan como hospederos de mayor preferencia para la reproducción y alimentación de H. lauri y S. catenifer, si se comparan con las variedades de uso comercial. Estos hallazgos son consistentes con investigaciones anteriores que han destacado la influencia de factores como la exposición a la luz solar y la proximidad a otros árboles infestados en la distribución de poblaciones de insectos plaga (Hahn et al., 2017; Martins et al., 2018; Pinchao & Carabali-Muñoz, 2019; Vacari et al., 2021).
La agregación espacial del daño alcanzó una mayor definición en las primeras etapas de desarrollo fenológico de formación de frutos, que fueron registrados en los primeros monitoreos, y determinaron la presencia de áreas con concentración del daño producido por S. catenifer y H. lauri hasta 35 m de radio. Los resultados encontrados en el estudio concuerdan con los reportados en cultivos de aguacate en México, donde las mayores capturas de poblaciones de S. catenifer se presentaron en las etapas de fructificación (del Carmen Velázquez-Martínez et al., 2022). Así mismo, la aplicación del método de la Ley de potencia de Taylor (LPT) en este estudio, confirma y fortalece las observaciones realizadas, al revelar una tendencia significativamente positiva de la agregación del daño de H. lauri y S. catenifer en los cultivos de aguacate cv. Hass. Este enfoque ha demostrado ser un complemento efectivo a otras técnicas para evaluar la distribución espacial de especies de insectos plaga (Deh-pahni et al., 2020; Jamshidi et al., 2022).
Existe consenso generalizado sobre la importancia y utilidad de la estadística espacial en la identificación de patrones y variaciones, no obstante, para las poblaciones de S. catenifer y H. lauri su aplicación no había sido estudiada. Los resultados obtenidos, además, de ampliar el conocimiento sobre la determinación e interpretación de la agregación del daño, contribuyen a generar nuevas investigaciones que den respuestas sobre los factores e interacciones que favorecen la presencia de poblaciones y daños en sitios específicos de los cultivos. Estudios sobre las distribuciones espaciales de las plagas en cultivos agrícolas han encontrado patrones y distribuciones que contribuyen en los programas de monitoreos y de manejo de plagas demostrando la predominancia en sitios de infestación, su distribución espacial y su evolución en los diferentes cultivos (Dionisio et al., 2020; Fatemi et al., 2023). Estudios han demostrado la asociaciones que presentan las incidencias de las diferentes plagas con los estados fenológicos y las condiciones microclimáticas que presentan cada sitio de agregación (Dawar et al., 2022; Mosedale et al., 2024). Los resultados analizados de manera integral sugieren la necesidad de formular estrategias de manejo específicas para estas áreas.
La aplicación de métodos como la función K de Ripley, la ley del potencia de Taylor y la interpolación de densidad de Kernel han permitido comprender las dinámicas de daño en tiempo y espacio a pequeñas escalas (Jiménez & Corredera, 2015; Rozas & Camarero, 2005; Teixeira Roth & Sánchez Infantas, 2006). Estos hallazgos sugieren que adicional a la identificación de patrones, los métodos representan una herramienta útil e informativa de aplicación en programas de manejo integral de plagas específicos por sitio, donde se consideren los resultados de las distancias de los sitios de agregación, estados fenológicos dentro de cultivo y zonas recurrentes de infestación. Se han utilizado estas herramientas para obtener información de la distribución espacial de plagas de cultivos con el objetivo de utilizar métodos apropiados de control y monitoreo de plagas (Bakry & Shakal, 2020; Guimapi et al., 2020). Al considerar la agregación del daño, los resultados de este estudio tienen implicaciones prácticas para mejorar los programas de monitoreo y control, aspecto que tradicionalmente no se ha tenido en cuenta y que puede pasar desapercibido. Esta comprensión detallada de la distribución del daño y su fluctuación temporal contribuye directamente al desarrollo de estrategias efectivas de manejo de poblaciones, especialmente para cumplir con los estándares de calidad necesarios para los mercados de exportación.
En particular, los resultados sugieren que los nuevos esquemas de manejo donde se incorporen programas de monitoreo deben considerar el conocimiento de la agregación del daño y direccionar orientar los diseños y unidades de muestreo a los sitios con mayor riesgo de infestación. Los programas de manejo y estrategias de reducción de poblaciones también deben considerar la información de los sitios de mayor agregación del daño, utilizando estrategias con mayor especificidad en sitios con alta concentración de plagas, comúnmente conocidos como focos.
En síntesis, los resultados obtenidos en el estudio no solo proporcionan una visión detallada de los patrones espaciales del daño en frutos causado por S. catenifer y H. lauri, sino que también brindan información y herramientas adicionales para la construcción de esquemas fitosanitarios con mayor eficacia y eficiencia, destacando la importancia de considerar la ecología específica de estas plagas en los cultivos de aguacate Hass. Estudios adicionales son necesarios para evaluar los factores bióticos y abióticos que determinan la agregación de estas plagas en cultivos de aguacate Hass. Adicionalmente, se recomienda realizar estudios complementarios para confirmar los patrones espaciales identificados en este estudio en otras regiones aguacateras del país.
Conclusiones
Los resultados de este trabajo indican que existen sitios influenciados por factores que favorecen el establecimiento y desarrollo de poblaciones de S. catenifer y H. lauri en zonas productoras de aguacate. En la determinación de los sitios se pueden utilizar análisis que permitan la definición de zonas donde existe una mayor probabilidad de encontrar mayor daño en la fruta. Estos conocimientos en su conjunto pueden conducir a alcanzar una mayor eficiencia de los diseños de experimentos de manejo de plagas, reducción de la variabilidad de los ensayos de campo y un manejo localizado más eficiente del control de plagas.
Los resultados permiten concluir que S. catenifer y H. lauri exhiben una distribución espacial agregada que se caracteriza porque la probabilidad de ocurrencia de un individuo de cada especie aumenta la del otro en un rango de distancias de 5 a 35 m. Su aplicación es de alta importancia en la agricultura sostenible y su campo de investigación se encuentra en constante desarrollo. Las metodologías y resultados encontrados en el estudio tendrán implicaciones en los nuevos programas de manejo integrado de plagas de importancia cuarentenaria de aguacate.
Contribución de los autores
Juan Camilo Zapata participated in the data collection process, data processing, experiment design, analysis and discussion of the information in the manuscript and in the writing of the manuscript.
Arturo Carabali participated in the data collection process, data processing, experiment design, analysis and discussion of the information in the manuscript and in the writing of the manuscript.
David Arango Londoño participated in the data processing, experiment design, analysis and discussion of the information in the manuscript and in the writing of the manuscript.
Conflicto de intereses
The authors declare no conflict of interest.