
Abstract
In the Colombian Caribbean, in addition to landscape modifications due to changes in land use, marked periods of drought and rainfall affect the survival of organisms. These changes modulate the supply of resources for scarabs with broad feeading habits. The spatio-temporal variation of scarab beetle guilds in a fragment of Tropical Dry Forest (Bs-T) and a surrounding crop was evaluated in six samplings; in each zone, four light traps were placed between18:00 to 6:00 and temperature, humidity and precipitation variables were measured. In the forest 5,184 individuals distribuited in 19 species were collected, and in the crop 1,919 individuals belonging to 16 species of subfamilies Dynastinae, Melolonthinae and Rutelinae. The phytophagous scarab beetles from forest and crop were represented by four guilds: Rhizo-Phylophagous (RP), Sapro-Caulophagous (SC), Sapro-Meliphagous(SM) and Xilo-Philophagous (XP); the predominant guild corresponded to the RP with ten species. In forest and crop, the dominant guild was RF with ten species, and the least number of species and guilds were collected in March, July and August, while May presented the highest diversity of species and guilds. The above coincides with the increase of precipitation, that showed positive correlation with richness and abundance in forest (p = 0.77, both cases) and crop (p = 0.84 and p = 0.66, respectively) during May. The guilds of phytophagous fluctuated in composition, richness, and abundance mainly in response to climatic seasonality, rather than the characteristics of each of the area.
Introducción
La distribución espacial y temporal de los organismos se explica principalmente por los recursos disponibles para sus actividades vitales, por las interacciones que se dan entre ellos y por el régimen climático a través del tiempo (Zobel 1997); lo que define la manera como se estructuran las comunidades en los ecosistemas. Sin embargo, la transformación de los bosques es uno de los impactos ambientales que más consecuencias trae a la biodiversidad local, ya que genera cambios drásticos en los hábitats, alterando los atributos de los ensambles establecidos (Martino 2007).
En el departamento del Atlántico los bosques son sometidos a una gran presión antrópica por extracción de madera y uso agrícola (Martínez-Hernández et al. 2010). Debido a esto, las coberturas boscosas presentan cambios que propician la dominancia de pocas especies con mayores capacidades de soportar condiciones de perturbación y que desplazan a otros grupos de insectos que cumplen importantes roles ecológicos, tales como escarabajos fitófagos de la familia Scarabaeidae. La mayoría de sus integrantes, tienen hábitos fitófagos, y pocos son depredadores. Estos organismos se caracterizan por tener amplia variedad de funciones en los ecosistemas como degradadores de materia orgánica, consumidores primarios y secundarios, polinizadores, así como alimento de otros insectos y varios grupos de vertebrados (Morón 1994). Por lo tanto, en ambientes forestales tienen un impacto en el ecosistema tanto por su participación en los procesos de fragmentación y reciclaje de la madera y la hojarasca como por su posible papel como plagas (Morón 1985; Aragón et al. 2013).
En Colombia las investigaciones en este grupo se han incrementado los últimos años. Actualmente se cuenta con importante información sobre inventarios locales y nacionales (Restrepo-Giraldo et al. 2003; Suárez y Amat-García 2007; Gasca-Alvarez y Amat-García 2010; Sanabria-García et al. 2012; Vallejo y Wolff 2013; García-Atencia et al. 2015; García-Atencia y Martínez-Hernández 2015; López-García et al. 2015a, b) e identificación de especies y fenología de reconocidas plagas agrícolas (Pardo-Locarno et al. 2012; Vallejo y Wolff 2013).
Las investigaciones sobre diversidad de escarabajos fitófagos son escasas. Sin embargo, se resaltan los realizados por Alcázar-Ruíz et al. (2003), Reyes-Novelo y Morón (2005), García-de Jesús et al. (2016), Ibarra y Damborsky (2017) y Serrano y Barro (2017) para el neotrópico, y las investigaciones de García-Atencia et al. (2015) y Villalobos-Moreno et al. (2018) para Colombia. En los bosques secos del Caribe colombiano las contribuciones en este campo han obtenido importantes resultados para el entendimiento del papel de estas comunidades en los ecosistemas (Martínez-Hernández et al. 2010; Pardo-Locarno et al. 2012; García-Atencia et al. 2015; García-Atencia y Martínez-Hernández 2015). Si bien, García-Atencia et al. (2015) estudiaron previamente la variación de las comunidades de estos escarabajos en un fragmento de Bs-T, no tuvieron en cuenta la importancia de los policultivos como elementos que integran la matriz de estudio. Por lo tanto resulta necesaria la evaluación de la estructura de la comunidad en una matriz común en la región de estudio. Esto representaría un avance en el conocimiento ecológico de estas las comunidades y su comportamiento en un contexto de cambio por las condiciones naturales de ecosistema y antropogénico.
En el presente estudio se analizaron los cambios estacionales de los gremios de escarabajos fitófagos, en un bosque y un cultivo, y el cambio entre las comunidades de ambos tipos de vegetación, con el fin de aportar al conocimiento base para el uso y la conservación de los fragmentos de Bosque seco Tropical en el departamento del Atlántico.
Aunque los gremios de los Scarabaeidae fitófagos no están claramente descritos, la propuesta de Morón y Deloya (1991), modificada por Deloya et al. (2007), representa una contribución al entendimiento de cómo se diferencian las comunidades y cómo se asemejan en términos de fenotipo y de respuesta al ambiente (Colwell y Fuentes 1975). Es decir, debido a que las especies de una comunidad no están igualmente afectadas por los cambios en los ecosistemas, el análisis de gremios puede tener una mayor aproximación al entendimiento de respuesta de los organismos ante disturbios ambientales.
Materiales y métodos
Área de estudio.
La Reserva Campesina de La Montaña -RCM- (o Tierra Arena), se encuentra ubicada en el municipio de Juan de Acosta, departamento del Atlántico, Colombia; localizada a 10°46′2,6″N y 75°0,2′34″O, con altitudes entre 177 y 250 m (García-Atencia et al. 2015). La RCM forma parte del distrito Montes de María y Piojó, dentro de la región biogeográfica del Cinturón Árido Pericaribeño (Hernández-Camacho 1992). Los meses de mayor precipitación histórica corresponden a agosto y septiembre con promedios que van desde 100 mm/mes hasta 580 mm/mes. Enero, febrero y marzo corresponden a los meses con menores precipitaciones, que van desde 0 hasta 30 mm/mes y la temperatura oscila entre los 25°C y 33°C (García-Atencia et al. 2015).
El presente estudio se realizó en un fragmento de Bs-T según lo descrito por Holdridge et al. (1971) y un policultivo adyacente separado a 1,5 km aproximadamente. El bosque, cuenta con una extensión de alrededor 47 ha y altitud máxima de 220 msnm. La vegetación predominante es la típica de BsT, descrita por Pizano y García (2014) en la que se presentan tres estratos vegetativos de dosel, arbustivo y sotobosque, y están presentes especies vegetales como Aphelandra pulcherrima (Jacq.) Kunth., Aspidosperma polyneron Müll. Arg., Astronium graveolens Jacq., Bursera simaruba (L.), Calycophyllum candidissimum (Vahl) DC., Capparis frondosa Jacq., Capparis odorattisima Jacq., Clavija sp., Hura crepitans L., Meliococcus oliviformis (Kunth), Ruelia macrophylla Vahl., Spondias mombin L., y Trichilia acuminata (Humb. & Bonpl. Ex Roem. & Schult.).
El policultivo, corresponde a un área de 30 ha con altura máxima de 241 msnm, dedicada a la siembra de Capsicum annuum L. (ají), Citrullus lanatus (Thunb.) Matsum. & Nakai (sandía), Cucumis sativus L. (pepino), Cucurbita moschata Duchesne (ahuyama), Manihot esculenta Crantz (yuca), y Zea mays L. (maíz). También pueden encontrarse árboles de Crataeva tapia L. (naranjuelo), Bactris sp. (palmas de corozo), Maclura tinctoria (L.) (dinde), Platymiscium pinnatum (Jacq.) (trevol), y Sterculia apetala (Jacq.) H. Karst. (camajón), dispersos por el área a distancias que no sobrepasan los 30 m y cercas vivas de troncos de Gliricidia sepium (Jacq.) Kunth ex Walpers (matarratón), que rodean la zona.
Recolecta de individuos.
Se realizaron seis muestreos entre marzo y agosto del 2013. Se colocaron ocho trampas de luz tipo embudo separadas aproximadamente 150 m entre sí, cuatro en bosque y cuatro en cultivo en forma de cuadrilátero para evitar el marcado efecto borde. Las trampas operaron una noche por mes desde las 18:00 hasta las 6:00, para un total de 72 horas de inversión y 48 muestras analizadas. Los días de recolecta se seleccionaron según el calendario lunar, lo que aseguro un muestreo nocturno de máxima oscuridad.
En cada área se tomaron las variables de temperatura y humedad ambiental con ayuda de Dataloggers marca GSI Super Quality. Para los valores de precipitación, se instaló un pluviómetro marca Hellman en el borde del fragmento, para obtener registros diarios de este parámetro en la RCM.
Fase de laboratorio.
Los ejemplares recolectados se preservaron en etanol al 70 % con su respectivo rótulo. Posteriormente, se determinaron con las claves propuestas por Endröd (1985), Ramírez-Ponce y Morón (2009), Sanabria-García et al. (2012) y Vallejo y Wolff (2013). Adicionalmente, para cada especie se tomaron datos de longitud corporal para la estimación de la biomasa y se clasificaron según los gremios propuestos por Deloya et al. (2007). El material producto de este trabajo, está depositado en la Colección de Entomología del Instituto de Ciencias Naturales (ICN) en la ciudad de Bogotá.
Análisis de los datos.
Se determinó la riqueza de especies y gremios capturados por muestreo y sitio; así como la abundancia (número de individuos) por especie. La biomasa se estimó como la cantidad de miligramos (mg) aportado por especies y gremios. Esta última fue calculada mediante la ecuación:
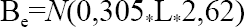
Donde, Be es la biomasa en mg, N es la abundancia y L es la media geométrica de la longitud corporal para las clases de tamaño relevantes. Se adoptaron las clases de tamaño citadas por Stork y Blackburn (1993) citadas en Sánchez y Amat (2005).
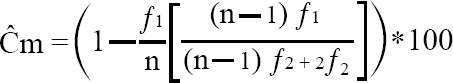
Para evaluar la representatividad de los muestreos, se estimó la cobertura de la muestra (Ĉm) para cada sitio (Chao y Jost 2012) que es calculada por la fórmula:
Donde n es la abundancia de la muestra y f1 y f2 son los números de singletons (especies con un individuo en las muestras) y doubletons (especies con dos individuos en las muestras), respectivamente. Ésta, es una medida de completitud, dada la porción del total de individuos en una comunidad que pertenece a las especies representadas en la muestra (Chao y Jost 2012)
Se estimó la diversidad alfa, basada en la diversidad verdadera, donde: 0D es igual a la riqueza de especies, por lo que es insensible a las abundancias de las mismas; 1D (exponencial de Shannon), representa al número efectivo de especies de una comunidad teniendo en cuenta las abundancias de las especies comunes; y 2D (inverso de Simpson) tiene en cuenta las abundancias de las especies que dominan la comunidad (Jost 2006). Los valores se calcularon con el programa iNEXT Online (Hsieh et al. 2020). Con estos se realizaron perfiles de diversidad por sitio y muestreos, acompañados de curvas de rango abundancia.
La diversidad beta fue estimada según lo propuesto por Carvalho et al. (2012), donde la disimilitud total (βcc) incluye los componentes recambio de especies (β-3) y la disimilitud debida a las diferencias en la riqueza (βrich). Según este enfoque particionado, la máxima disimilitud se obtiene con un valor igual a 1. Todo fue calculado con el paquete BAT de R desarrollado por Cardoso et al. (2015).
Se utilizó la prueba pareada no paramétrica de U Mann-Whitney para evidenciar si existían diferencias significativas entre abundancia de especies, biomasas de especies y abundancia y biomasa de gremios entre áreas. Así mismo una prueba de Kruskall-Wallis para saber si existían diferencias significativas entre la relación de la biomasa de especies y de gremios con el número de individuos en bosque y cultivo. Como complemento se realizaron correlaciones de Spearman entre las abundancias de gremios y las variables ambientales. Todo esto con ayuda del software R versión 1.2 5042 (Rstudio Team 2020).
Resultados y discusión
Se recolectaron 7.018 individuos agrupados en 23 especies pertenecientes a las subfamilias Dynastinae, Melolonthinae y Rutelinae y en cuatro gremios gremios: Rizo-Filófago (RF), Sapro-Caulófagos (SC), Sapro-Melífagos (SM) y Xilo-Filófagos (XF) (Tabla 1). La mayor riqueza la presentó la subfamilia Rutelinae con once especies, mientras que la mayor abundancia la presentó la subfamilia Melolonthinae con 5.180 especímenes. Las especies Phyllophaga pachypyga (Burmeister, 1855) y Astaena sp. aportaron un 29,2 % y 29,6 % a la abundancia total, respectivamente. Para ambos usos del suelo estudiados, la cobertura del muestreo estuvo en el 99 % de confiabilidad.
Abundancia y biomasa (mg) de especies de los ensambles de escarabajos (Coleoptera: Scarabaeidae) fitófagos en bosque y cultivo de la RCM, departamento del Atlántico, Colombia. Abreviaturas: SM, Sapro-Melífagos; RF, Rizo-Filófagos; SC, Sapro-Caulófagos; XF, Xilo-Filófagos; S, riqueza; N, abundancia.
En el bosque se recolectaron 19 especies y 5.184 especímenes, y en el cultivo 16 especies y 1.919 individuos. La abundancia total de los fitófagos en las dos áreas no presentó diferencias significativas según la prueba pareada no paramétrica U Mann-Whitney (U = 170; P > 0,05).
De acuerdo con los resultados obtenidos, el Bs-T estudiado alberga el 3,9 % de los escarabajos fitófagos (Scarabaeidae) reportados para Colombia y el 41,7 % de los descritos para el Caribe colombiano por Restrepo-Giraldo et al. (2003). Las altas abundancias se explican en gran medida por la presencia de Liogenys quadridens y P. pachypyga, de la subfamilia Melolonthinae y Paranomala inconstans de la subfamilia Rutelinae.
La mayor riqueza y abundancia la presentó el gremio RF con 16 especies y 6.897 individuos, respectivamente. Este gremio aportó el 97,5 % a la abundancia total de los gremios encontrados en la RCM. En bosque y en cultivo, el gremio predominante correspondió al RF con diez especies en cada sitio.
Los RF que, por su biología, pueden corresponder a potenciales plagas agrícolas, se caracterizan por tener estrategia de reproducción r con gran tolerancia a las perturbaciones de los hábitats. Estas especies aprovechan los recursos que abundan en sistemas homogéneos, donde en los últimos años han predominado épocas prolongadas de sequía y perturbación antropogénica que induce a la homogeneidad en los recursos.
El aporte de D. dubius al gremio fue insignificante y sólo estuvo presente en cultivo, hábitats donde se les ha recolectado como hospederos de plantas de maíz (Z. mays), arroz (Oryza sativa) y malanga (Xanthosoma sagittifolium) (Joly y Escalona 2010).
Variación espacial.
Los mayores valores de la diversidad verdadera corresponden al bosque, mientras que el cultivo presentó un patrón similar con menor diversidad (Fig. 1). Los valores de las diversidades 1D y 2D reflejan la alta dominancia por pocas especies de la comunidad evidenciando que la estructura de los ensambles presentes en cada sitio se caracteriza por tener muchas especies con abundancias muy bajas y pocas especies dominantes. Por su parte βcc arrojó una disimilitud de 0,58, es decir que las especies no compartidas representan el 58% del número total de especies. De esto, el 45% se le atribuye al recambio en la riqueza de especies (β-3) que indica que ambos sitios presentan comunidades similares con pocas especies exclusivas; mientras el 13% a la diferencia en riqueza (βrich), que evidencia que hay pocas únicas de cada sitio.
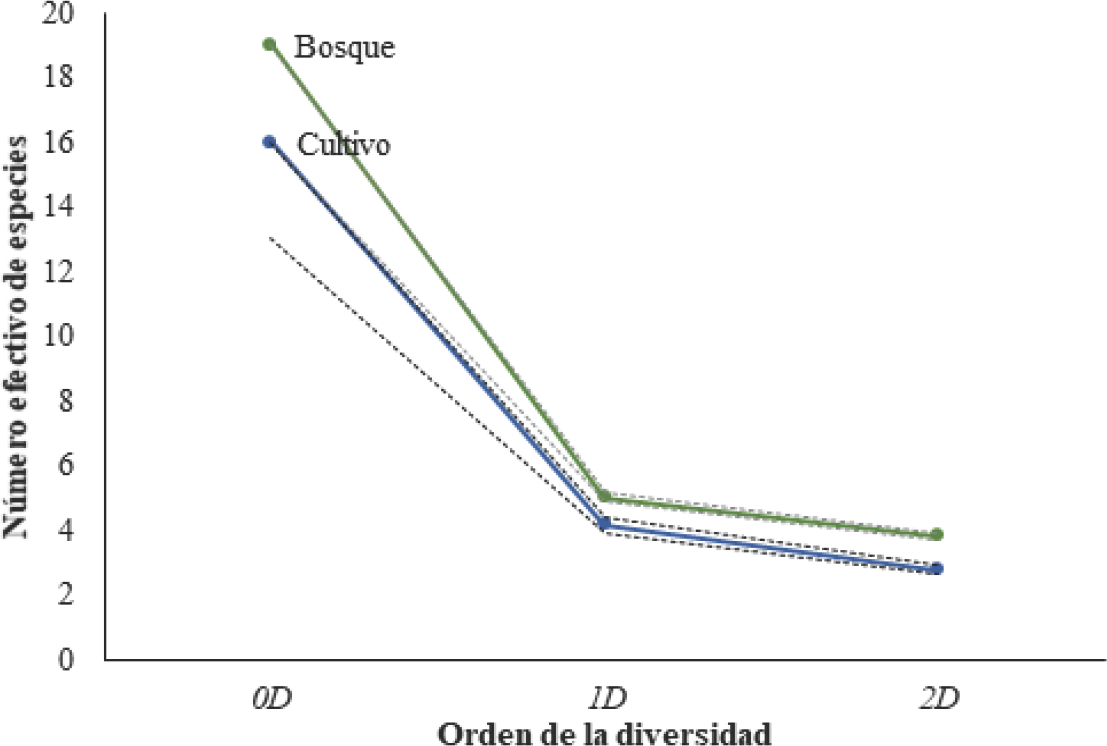
Perfil de diversidad de escarabajos fitófagos (Coleoptera: Scarabaeidae) para bosque y cultivo de la RCM. Líneas punteadas indican intervalos de confianza.
La mayor abundancia y riqueza de escarabajos del Bosque, corresponde a la mayor heterogeneidad de recursos que permiten un mayor establecimiento y reproducción de las poblaciones. Por su parte, en el cultivo se presentan condiciones más homogéneas con permanencia muy limitada, pues los pobladores de la zona intercalan las áreas de cultivo cada año para evitar la pérdida en la calidad del suelo, lo que hace que sea aprovechado por aquellas especies capaces de explotar los recursos que allí se ofrecen garantizando su éxito reproductivo reflejado en sus altas abundancias. Sin embargo, aunque no existen diferencias significativas en la riqueza y la abundancia en los dos sitios, si se demuestra una diferencia en la composición de especies.
En el bosque Astaena sp., P. inconstans y P. pachypyga, del gremio RF, representan el 84% de la abundancia total mientras que E. humilis presentó el menor número de individuos. En el área del cultivo, las especies dominantes corresponden también al gremio RF y son L. quadridens, P. inconstans y P. pachypyga y (Fig. 2). Además, P. pachypyga comprende más del 50% del total con más de 1.000 individuos y sumado con L. quadridens y P. inconstans, representan el 85 % de la abundancia total. Esta alta dominancia de especies con roles similares, pero con distintas abundancias en cada sitio, puede ser el resultado de hábitats degradados que, aunque albergan un número considerable de especies, las características intrínsecas de los sistemas y el estrés ambiental agravan las condiciones. Sin embargo, la estructura de la comunidad presenta diferencias entre hábitats.
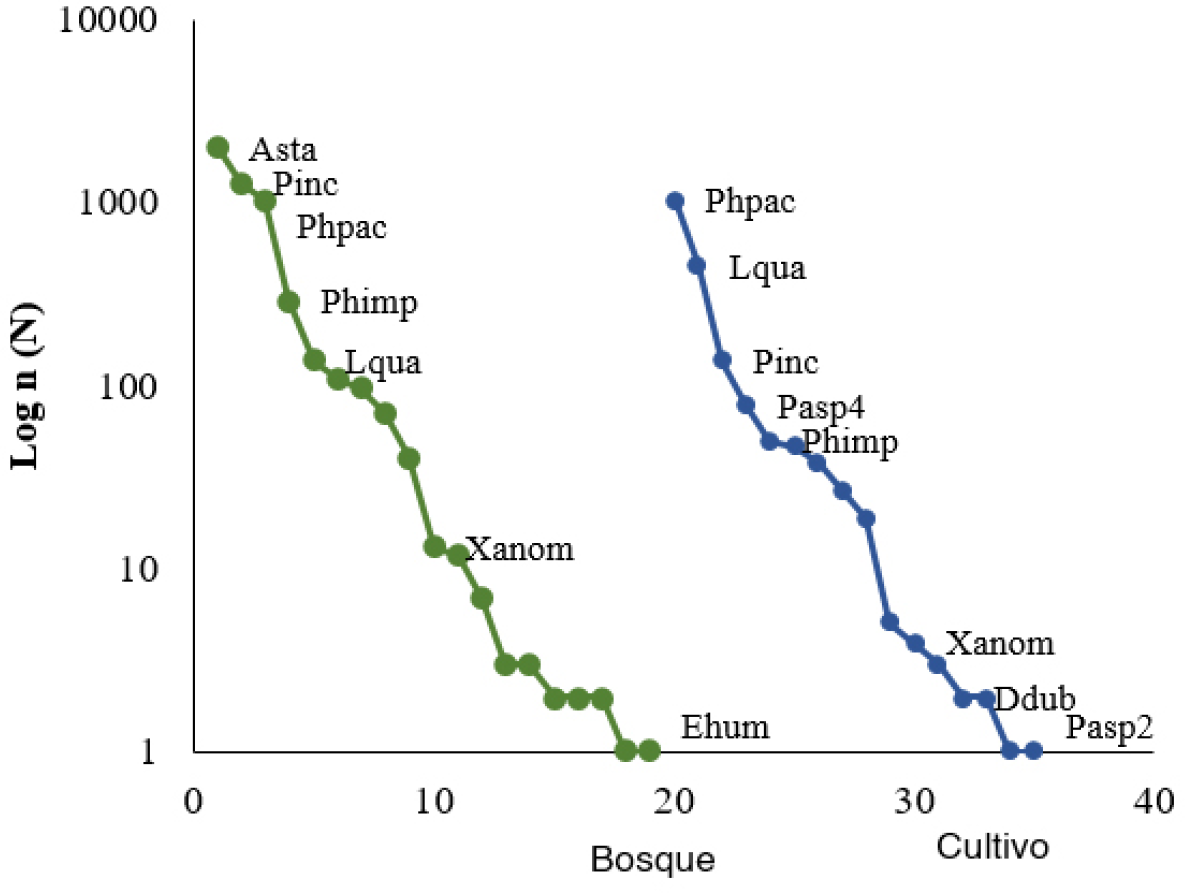
Curvas rango-abundancia en Bosque y cultivo de la RCM. Abreviaturas: Asta, Astaena; Pinc, Paranomala inconstans; Phpac, Phyllophaga pachypyga; Phimp, Phyllophaga impressipyga; Lqua, Liogenys quadridens; Xanom, Xenopelidnota anomala; Ehum, Euetheola humillis; Pasp4, Paranomala sp. 4; Ddub, Dyscinetus dubius; Pasp2, Paranomala sp. 2.
El análisis no paramétrico, mostró que no hay diferencias significativas entre la biomasa de las especies en las áreas muestreadas (W = 167; P > 0,05). Aunque, el aporte de P. pachypyga a la biomasa en ambos sitios fue similar, siendo ligeramente mayor en el cultivo. Por su parte, Phyllophaga impressipyga (U = 205,5; P < 0,05), Astaena sp. (U = 140; P <0,05), Paranomala sp. 3 (U = 216; P < 0,05) y P. menetriesi (U = 204; P < 0,05) presentaron diferencias significativas en cuanto a la biomasa aportada en cada sitio. En cuanto a las especies que dominaron por biomasa, se encontraron para el bosque Phyllophaga sp. 1 y P. inconstans, y para el cultivo, Phyllophaga sp. 1, siendo estas especies las que aportaron más de 150 mg en cada hábitat. Con estos datos, se observó que algunas de las especies abundantes, no dominaron en términos de biomasa.
Con respecto a la biomasa de los gremios de Scarabaeidae fitófagos, los valores no presentaron diferencias significativas entre bosque y cultivo (W = 1,5; P > 0,05) (Fig. 3). Todos los gremios recolectados en el presente estudio se registraron en las dos áreas muestreadas, con mayor biomasa en el Bosque. Los RF en ambas áreas se caracterizaron por ser los que más aportaron a la biomasa, seguido por los XF. El aporte de los SM y SC fue menor de 1mg. Este último presentó un aporte insignificante con respecto a los demás.
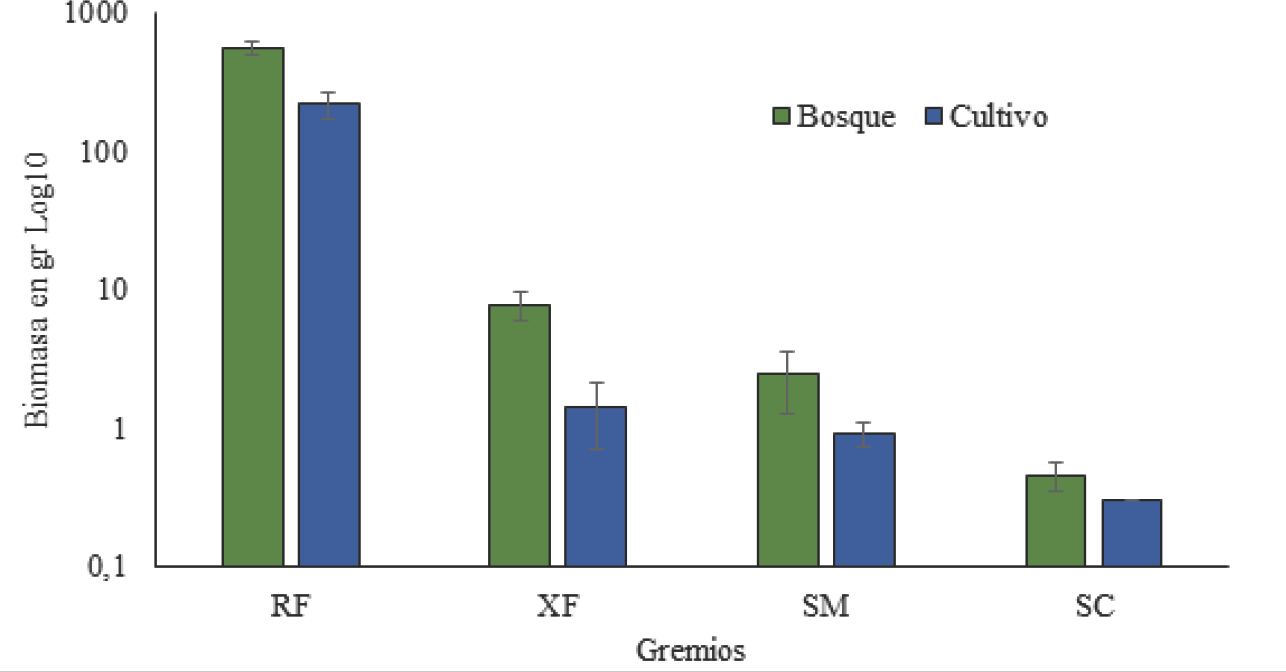
Aporte de la biomasa en miligramos de los gremios de escarabajos fitófagos (Coleoptera: Scarabaeidae) en bosque y cultivo de la RCM, departamento del Atlántico, Colombia. Abreviaturas: RF, Rizo-Filófagos; XF, Xilo-Filófagos; SM, Sapro-Melífagos; SC, Sapro-Caulófagos.
Para el bosque, la biomasa está dominada por P. inconstans que además de ser abundante presentó tallas mayores a 15 mm. Esta especie ha sido considerada invasora que frecuenta pastizales inducidos y cultivos de gramíneas (Ramírez-Salinas et al. 2004). Por su parte en el cultivo, L. quadridens, presentó la mayor biomasa, por lo que la ausencia y/o poca abundancia de especies de tallas más grandes representaron este hábitat donde la vegetación es homogénea, el estrato arbóreo está ausente y los arbustos muy separados entre sí. Adicionalmente, las especies de talla pequeña tienen un metabolismo más rápido, lo que las obliga a estar más concentradas en los sitios que brindan recursos energéticos de manera permanente. Lo anterior refleja que aunque ambos sitios presenten condiciones de perturbación, en la comunidad de insectos fitófagos del cultivo abundan las formas pequeñas, o bien “oportunistas” (Clarke y Warwick 2001).
En cuanto a la relación entre la biomasa de gremios y el número de individuos, no se presentaron diferencias significativas en el bosque (H = 1,33; P > 0,05) ni en el cultivo (H = 2,08; P > 0,05). Sin embargo, existen diferencias significativas entre la biomasa de especies y la abundancia de las mismas, tanto en bosque (H = 5,08; P < 0,05) como en el cultivo (H = 7,78; P < 0,05).
En el hábitat de Bs-T, Cyclocephala spp., T. fossor y E. humilis se recolectaron en mayo y junio, pero la baja garantía de supervivencia dada por las condiciones ambientales no permitió el establecimiento de los gremios SM y SC. En el caso de E. humilis, Bernardi et al. (2008) lo reportaron para Brasil como plaga de cultivos de arroz, caña de azúcar, millo y eucalipto, al igual que T. fossor que ha sido encontrado en capítulos y raíces de arroz, caña de azúcar, girasol, y maíz (Escalona y Joly 2006). Algunas especies de Cyclocephala son frecuentes en inflorescencias de las Araceae que se caracterizan por presentar espata y espádice, lo que las hace atractivas y ventajosas para el resguardo de algunas especies; por otro lado, les brindan temperatura estable, alimento, refugio, posibilidad de hallazgo de pareja, retribuyéndoles con su condición de polinizadores (Morón et al. 1997; Gasca-Álvarez, 2013). Otras especies del género pueden estar relacionadas con flores actinomorfas y frutos de una gran variedad de plantas de la familia Araceae que pueden estar presentes en variados tipos de ecosistemas (Milet-Pinheiro et al. 2017). Sin embargo, las especies de estos tres géneros de escarabajos tienen en común que, aunque sus hábitos son poco conocidos, sus larvas podrían ayudar a la mineralización y algunos adultos a la polinización (Morón et al. 1997).
Según Morón (2004), la mayor parte de las especies de escarabajos edafícolas deben estar adaptándose a las condiciones de ecotono y de agro-ecosistemas, siendo también probable el aumento de especies originalmente asociadas a ambientes silvestres que puedan adaptarse a condiciones de monocultivo y que, por determinadas circunstancias ecológicas, puedan causar daños a las plantas de importancia económica. Por otro lado, la composición de la entomofauna usualmente se ve influenciada por las modificaciones en los hábitats naturales resultado de las actividades humanas, que propician la disminución de algunas especies y el éxito de otras a nivel local y regional.
Variación temporal.
Los escarabajos (Coleoptera: Scarabaeidae) fitófagos en los hábitats estudiados presentaron una marcada estacionalidad distribuida con baja densidad y corroborada por la correlación positiva de la riqueza y abundancia de especies y gremios con la precipitación; esto se refleja en los valores más altos de abundancia concentrados en el muestreo de mayo, en el que la intensidad de las lluvias se incrementó (Tabla 2). En general, la prueba de correlación de Spearman mostró una correlación positiva entre la precipitación con la riqueza y abundancia en bosque (p = 0,77 para los dos casos) y cultivo (p = 0,84 y p = 0,66, respectivamente). El comportamiento de la temperatura y la humedad fue similar para los dos sitios durante el periodo muestreado.
Variación temporal de la abundancia de gremios de escarabajos fitófagos y variables ambientales en bosque y cultivo de la RCM. RF, Rizo-Filófagos; SC, Sapro-Caulófagos; SM, Sapro-Melífagos; XF, Xilo-Filófagos; Temp., Temperatura; HR, Humedad Relativa; Precip., Precipitación.
Este patrón es similar a lo encontrado por García-Atencia et al. (2015) en el mismo lugar, en diferentes localidades de Colombia (Martínez-Hernández et al. 2010; Útima y Vallejo 2010) y en países como México (Alcázar-Ruiz et al. 2003; Reyes-Novelo y Morón 2005; Ramírez-Ponce y Morón 2009; Lugo-García et al. 2011), Puerto Rico (Martínez et al. 2009) y en la Florida, EE.UU. (Buss 2006). En estos estudios, se ha encontrado una estrecha relación entre la riqueza y abundancia con el inicio del período de lluvias, lo que sustenta la diversidad dada por los índices usados en este trabajo durante mayo y junio. Es en estos meses algunas especies salen de sus cámaras pupales, como el caso de los adultos de Phyllophaga spp. que se reproducen y aumentan sus actividades, por lo cual son comúnmente conocidos como los “cocuyos de mayo” (Morón 1994).
Para el bosque, los mayores valores de diversidad 0D, 1D y 2D se presentaron durante mayo que coincide con un ligero aumento de la precipitación, momento en el cual probablemente se incrementaron los recursos a explotar. Aunque los dos hábitats no presentaron diferencias en cuanto a 0D, en el cultivo, los mayores valores de 1D y 2D se registraron para M4 (Fig. 4). Los menores valores de diversidad se presentaron en el M5 y M6 en los que se presentaron valores bajos de precipitación. Esto refleja las pocas abundancias recolectadas en los hábitats, y en el caso particular del bosque, la no obtención de individuos. Aunque ambos sitios presentan patrones similares, la composición y abundancia de las especies es diferente, lo que quiere decir que hay una variación bosque-cultivo, que corresponde a un posible reemplazo de los taxones que cumplen funciones ecológicas equivalentes. Estos cambios se presentan cuando existen perturbaciones intensas o cambios radicales en el uso del suelo, dando lugar a la introducción de nuevos integrantes (tal vez generalistas o con amplio espectro ecológico), al cambio en el dominio de especies presentes e inclusive a la desaparición de algún taxón (Morón 2001).
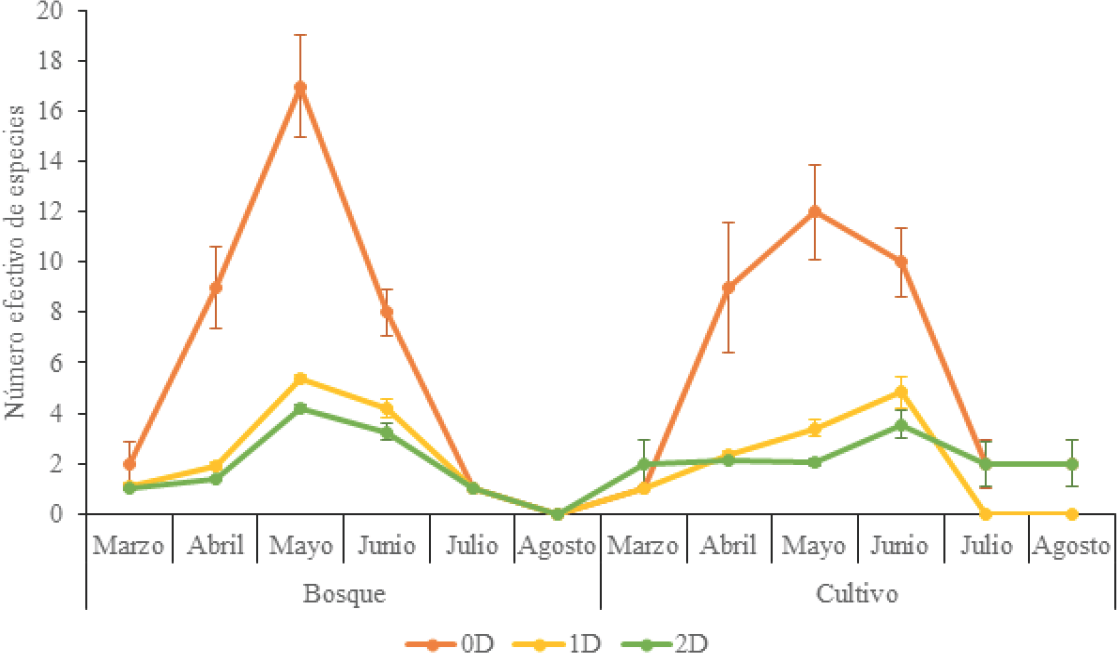
Perfil de diversidad °D, 'D y 2D de escarabajos fitófagos (Coleoptera: Scarabaeidae) recolectados en bosque y cultivo durante los meses de muestreo en la RCM, departamento del Atlántico, Colombia.
Según beta temporal total (βcc), en el bosque los muestreos fueron altamente disimiles entre ellos, teniendo valores por encima del 85 % de disimilitud. Al aparecer las lluvias en mayo, en los BsT producen la emergencia de plántulas y demás recursos verdes que constituyen la principal fuente de alimentos de los escarabajos fitófagos (Scarabaeidae). Sin embargo, la disminución abrupta y atípica de la precipitación impide el normal paso a una nueva temporada lluviosa en estos ecosistemas, lo que disminuye la calidad y cantidad de los recursos a consumir y nuevos microhábitats que le sirven a los herbívoros como refugio ante posibles amenazas de sus depredadores (Huston 1994, Barraza et al. 2010).
Para el cultivo, la tendencia fue similar a excepción de la disimilitud de mayo con junio que fue del 44%. Si bien, la dominancia es mayor en los meses con bajas precipitaciones, especialmente durante marzo en las dos áreas de estudio, dada la alta abundancia de Astaena sp. en bosque y L. quadridens en cultivo. En general, la tendencia en la variación temporal correspondió a que la diversidad beta estuvo más influenciada por la pérdida de especies que por el reemplazo. Esto, por la sincronización de la aparición de especies con el período de máximas lluvias, lo que permitió un aumento en la riqueza de especies en pocos meses de muestreo.
En el bosque la biomasa de gremios por muestreo presentó diferencias significativas (H = 9,68; p = 0,03), siendo mayo el que presenta la mayor disimilitud con respecto a marzo (p = 0,05) (Fig. 5). Para el cultivo, no se presentaron diferencias significativas, pero mayo marcó mayor diferencia con respecto a los dos últimos muestreos (p = 0,02). Esta alta diferencia con respecto a los otros muestreos obedece a la presencia de las especies más grandes muestreadas en toda la RCM, y algunas de ellas exclusivas de ese mes y de bajo porte, como P. polita recolectada sólo en el bosque y X. anomala con bajas abundancias en cultivo. Estas especies conforman el gremio XF. Aunque los dos sitios muestreados presentan el mismo patrón, en el bosque los cambios de la biomasa durante los meses de muestreo son más abruptos, lo que demuestra una mayor sensibilidad de la comunidad de insectos fitófagos de la familia Scarabaeidae a los fuertes cambios ambientales que se presentaron durante el año de muestreo.
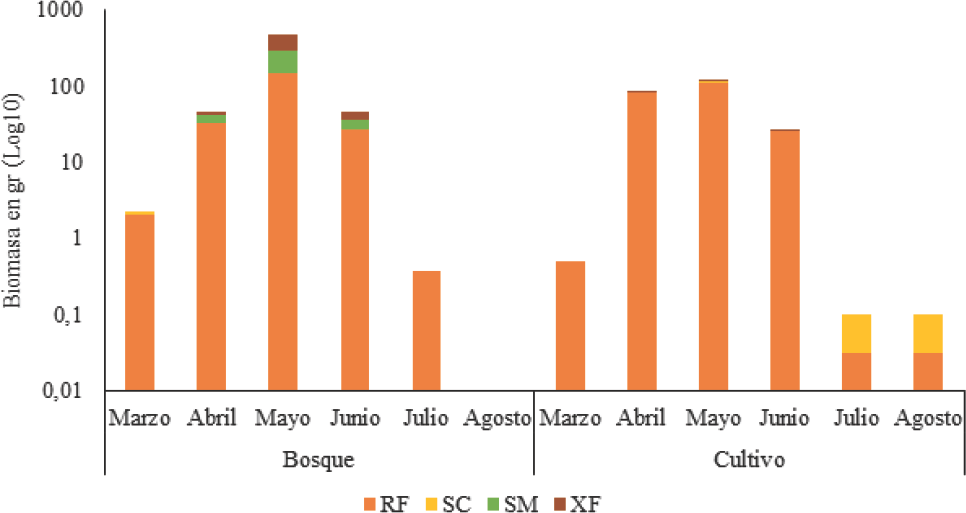
Variación de la biomasa por gremios de escarabajos fitófagos (Coleoptera: Scarabaeidae) en la RCM, departamento del Atlántico. Sm, Sapro-Melífagos; RF, Rizo-Filófagos; SC, Sapro-Caulófagos; XF, Xilo-Filófagos.
En ambos sitios, los gremios presentaron una correlación positiva con algunas variables ambientales, siendo la precipitación la más importante en términos de influencia en la fenología (Tabla 3). Por su papel como degradadores primarios y secundarios, los gremios de Scarabaeidae fitófagos pueden ser bioindicadores de ecosistemas importantes para la conservación (Morón et al. 1997; Deloya et al. 2007), considerando la diversidad de éstos y relacionándolos con la variedad de recursos disponibles.
Rangos de correlación de Spearman (valores r) en bosque (b) y cultivo (C). Valores que presentaron significancia (p > 0,05) aparecen en negrita.
Teniendo en cuenta que la composición gremial de escarabajos (Scarabaeidae) fitófagos es similar en bosques y cultivos, se infiere una baja oferta de recursos durante el tiempo de muestreo por las modificaciones del paisaje y las características intrínsecas del ecosistema que afectan la sincronía de algunas especies de esta familia con la aparición de recursos temporales en el Bs-T. Lo anterior, genera periodos cada vez más críticos de supervivencia que provocarían la desaparición de especies con nichos restringidos.
Conclusiones
La composición del bosque difirió a la del cultivo por tres especies exclusivas del primer sitio, lo que representa un bajo valor de diversidad beta dado por las características similares de los sistemas en cuanto a la disponibilidad de recursos. Las comunidades de escarabajos fitófagos y sus gremios fluctúan en composición, riqueza y abundancia principalmente como respuesta a la estacionalidad climática, más que a la homogeneización de las áreas por el uso agrícola.
En el caso de la biomasa, reflejó que las especies de tallas pequeñas dominaron más en el hábitat del cultivo que en bosque, lo que es una característica propia de sitios intervenidos, y para este caso, por condiciones antropogénicas y naturales. Si se considera que en la costa Caribe colombiana se pueden presentar tres épocas climáticas marcadas se concluye que el 2013 correspondió a un año atípico. Durante el período de muestreo el mayor valor de precipitación correspondió a 99 mm, lo que pudo haber impedido la continuidad en el incremento del número de especies e individuos en la RCM. Adicional a las intervenciones de los ecosistemas por la explotación agrícola y ganadera, los tiempos prolongados de sequía pueden repercutir en una posible alta mortalidad de adultos y disminución de la emergencia de pupas.
Contribuciones de los autores
SGA participó en la concepción y diseño del artículo; SGA y GAG definieron el contenido intelectual y la búsqueda de literatura; realizaron los estudios experimentales y la obtención de datos; participaron en el análisis de los datos experimentales; realizaron el análisis estadístico; fueron responsables de preparar el manuscrito y someterlo a publicación.
Conflictos de interés
Los autores declaramos no tener conflictos de intereses.
Origen y financiación
La investigación hace parte de la tesis de maestría de la primera autora, la cual recibió financiación otorgada al Grupo Insectos de Colombia mediante la Convocatoria Nacional apoyo para el Fortalecimiento de Grupos de Investigación que soporten programas de posgrado de la Universidad Nacional de Colombia-2012.