
Abstract
The richness, abundance and composition of hunting ants in three fragments of tropical dry forests in the northern region of Tolima, Colombia and their matrices were assessed; in addition, three scrub patches of shrub were included due to their high frequency in the landscape. Two samplings were carried out between May to June and October to November 2011. In each habitat, a linear 150 m transect was established in which pitfall traps were installed and 1mβ leaf-litter samples were extracted in mini-Winkler sacks were extracted. The 17 hunting ant species of Ectatomminae and Ponerinae found represent 44.7% of all the species reported for Tolima State. There significative differences in richness: 13 species were found in forests, seven in the matrix and 10 in the scrub patches and 17.17, 7, 11.94 species for forest, matrix ans scrub for estimated richness Chao 2. Locally, the species richness between forests and their matrices differed significantly only in the Centro Universitario Regional del Norte (CURN-UT), while the species composition was different in all sites and, but similar between forests and shrub patches. It is possible that the conversion of natural habitats into agricultural land areas become a modifying factor of the structure of hunting ants communities, that although in some sites the species richness is similar in forests and matrices, the species composition differs between them and in this way, the community responds to such changes
Introducción
De acuerdo con Kattán (2002), una de las causas más importantes de la pérdida global de diversidad, es la transformación de los bosques tropicales debido a actividades humanas. Esta transformación de hábitats naturales conlleva a la alteración de sus condiciones físicas, situación que puede incidir en la disponibilidad de los recursos y, en consecuencia, algunas especies de animales y plantas pueden resultar afectados de manera negativa y otros ser favorecidos (Didham et al. 1998; Desouza et al. 2001; Kattán 2002). Como resultado de la transformación, los fragmentos de bosque quedan inmersos en matrices modificadas, las cuales pueden representar diferentes calidades para la fauna.
Los hábitats que componen la matriz pueden llegar a contener una diversidad propia significativa que también interactúa con los fragmentos (Gascon et al.1999; Kattán 2002) y juegan un papel importante en el mantenimiento de la diversidad de los fragmentos de bosque que rodea, ya que su calidad puede permitir o bloquear la movilidad de organismos y, por tanto, procesos de migración entre parches (Perfecto 2003). De esta manera, las interacciones entre los elementos del paisaje pueden afectar la diversidad y abundancia de especies (Kattán 2002). En este sentido, el estudio de los componentes del paisaje adquiere importancia por cuanto su estructura puede determinar los patrones de distribución y movilidad de las especies.
En Colombia el bosque seco tropical (bs-T) es considerado uno de los ecosistemas más degradados, fragmentados y menos conocidos, debido a que los bosques se encuentran localizados en áreas de intenso uso ganadero y agrícola en las que se concentra gran parte de la producción del país. Específicamente, la región del valle seco del río Magdalena (Tolima, Cundinamarca y Huila), es una de las más desconocidas y con menos trabajos publicados. No obstante, esta región, ocupa el segundo lugar en cobertura boscosa dentro de las tres regiones con bosque seco tropical en Colombia (IAvH 1998).
Con base en lo anterior, se requiere evaluar el efecto de la fragmentación y el cambio en el uso del suelo sobre la fauna asociada a los tipos de hábitats, su caracterización, migraciones locales, dinámica e interacción, lo cual permitirá enriquecer el planteamiento y desarrollo de planes de conservación y restauración en el bosque seco tropical (IAvH 1998). En este sentido, el uso de insectos como organismos indicadores se convierte en una herramienta importante debido a su uso potencial para medir el grado de alteración de los ecosistemas (McGeoch 1998). Particularmente, las hormigas pueden ser muy sensibles a la transformación y perturbación del hábitat, por lo que han sido ampliamente usadas como indicadoras (Andersen 2010). En Colombia, los estudios relacionados con hormigas y paisajes fragmentados se han desarrollado principalmente en el bosque seco tropical en la cuenca del río Cauca y la región Caribe, en los cuales se han observado efectos variables de la fragmentación sobre la diversidad y composición de las comunidades de hormigas en diferentes hábitats, usando tanto grupos focales de hormigas como ensamblajes completos. Domínguez et al. (2007), reportaron similitud entre las hormigas cazadoras de fragmentos de bosque y sus matrices, argumentando movilidad entre hábitats continuos. En contraste, otros estudios reportan diferencias marcadas en cuanto a riqueza y composición de hormigas en bosques y matrices adyacentes (Armbrecht y Ulloa-Chacón 1999; Dix et al. 2005; Arcila-Cardona et al. 2007; Chacón de Ulloa et al. 2007), así como con el aumento en la diversidad y grupos funcionales a través de gradientes de restauración en un sitio de explotación minera de carbón a cielo abierto (Domínguez y Armbrecht 2011).
Debido a que las hormigas cazadoras (clado Poneroide y subclado Ectatomminoide) han sido caracterizadas como especies propias de áreas con cobertura arbórea, además de la capacidad indicadora de algunas de ellas (Arcila-Cardona et al. 2007; Chacón de Ulloa et al. 2007; Abadía et al. 2010), se propuso el presente estudio con el objetivo de caracterizar la comunidad de hormigas cazadoras del suelo y hojarasca, asociada a fragmentos de bosque seco tropical y sus matrices, en paisajes fragmentados del norte del departamento del Tolima.
Materiales y Métodos
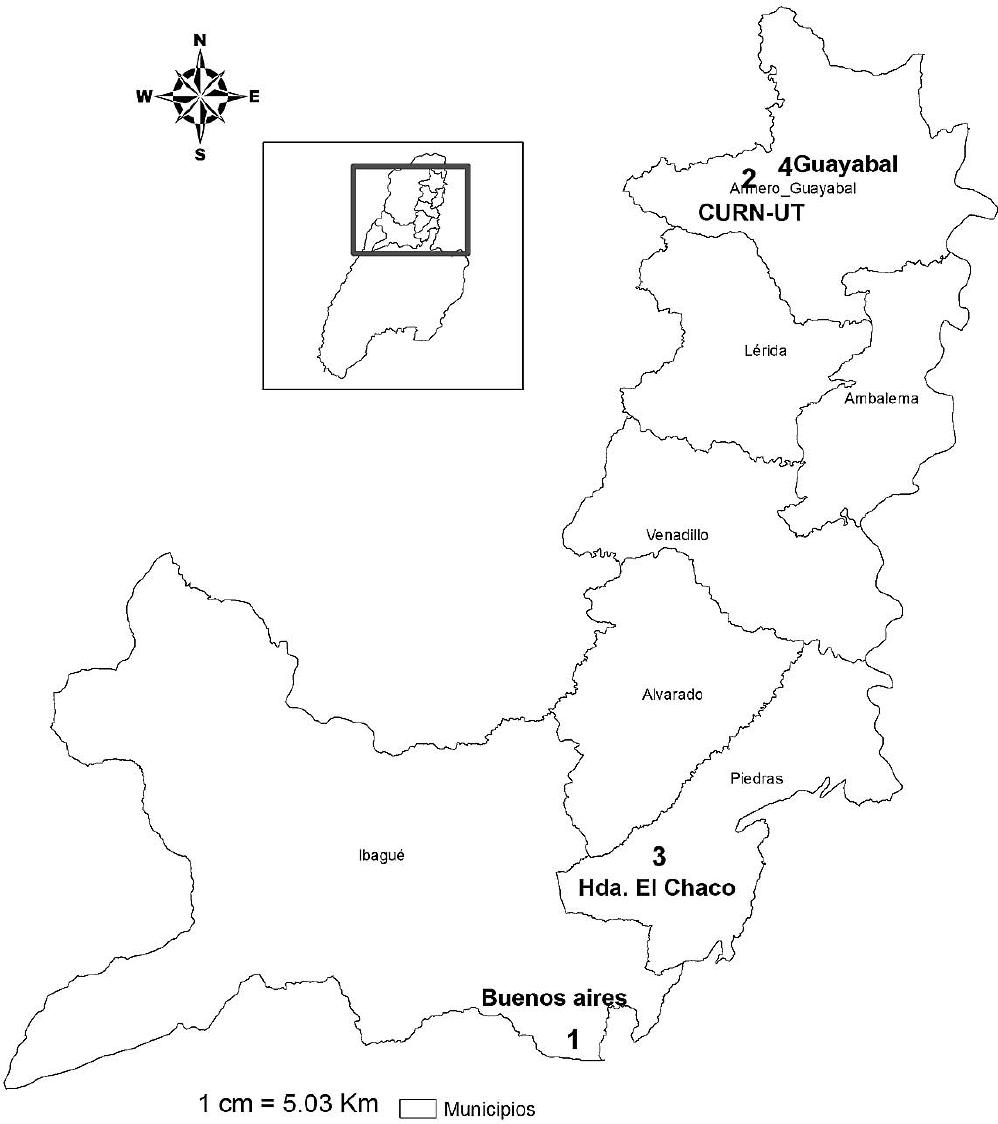
Ubicación de las localidades en los municipios que presentan la zona de vida bosque seco tropical en el norte del Tolima.
Sitios de muestreo definidos para la caracterización de las hormigas cazadoras del suelo y hojarasca en el bosque seco tropical del norte del Tolima.
Los fragmentos de bosque, con área aproximada de 10 ha, se consideran como bosques secundarios con poco grado de intervención, el dosel alcanza los 20 m, se distinguen dos estratos: el primero conformado por arbustos y árboles jóvenes y, el segundo, por los árboles del dosel con DAP > 10 cm. Las matrices, corresponden en todos los casos a suelos de uso agrícola con cultivos anuales o transitorios, principalmente de arroz y maíz. En la ventana de CURN-UT la matriz corresponde a lotes utilizados para las prácticas agrícolas docentes, en el que durante la realización del presente estudio se registró el cultivo de leguminosas, forrajes y de teca (Tectona grandis Linnaeus, 1781) hacia el final de la investigación. Los parches de matorrales se incluyeron ya que durante las visitas preliminares a las áreas de estudio se observó que conforman un tipo de hábitat frecuente en los paisajes de bosque seco en el Tolima. Estos parches, corresponden a áreas de regeneración natural que presentan doseles de hasta 7 m, en donde no se observa una estratificación vertical clara, es abundante la presencia de lianas y árboles jóvenes con DAP < 3 cm.
Se realizaron dos muestreos, uno entre mayo y junio y el otro entre octubre y noviembre de 2011. En cada tipo de hábitat, se estableció un transecto lineal de 150 m como lo sugiere Arcila-Cardona et al. (2007) y a una distancia mínima de 20 m, desde el borde hacia adentro del bosque y hacia la matriz. En cada transecto, cada 10 m, se utilizaron trampas de caída y sacos mini-Winkler. Las trampas se recogieron 48 horas después de su instalación y el contenido de los sacos mini-Winkler se tamizó previamente (hojarasca en 1 mβ). Un total de 216 trampas de caída fueron instaladas y 216 mβ de hojarasca procesados. El material recolectado se almacenó en alcohol al 70% y se procesó en el Laboratorio de Investigación en Zoología de la Universidad del Tolima. Para la determinación de los organismos se utilizaron las claves de Arias-Penna (2007), Fernández y Arias-Penna (2007), Mackay et al. (2007), Rodríguez (2007) y Lattke (2011). El género Hypoponera, fue agrupado en morfoespecies debido a la complejidad de su taxonomía, para lo que se tuvo en cuenta caracteres propuestos por Longino (2005) y se realizaron comparaciones con especímenes de la Colección Zoológica de la Universidad del Tolima (CZUT-Hm). Los organismos se encuentran en proceso de catalogación e ingreso a la CZUT-Hm.
Se obtuvo la lista de las especies de hormigas colectadas que fue comparada con los registros existentes para bosques del departamento del Tolima y bosques secos del país. Se evaluó la diversidad de los tipos de hábitat a partir de la riqueza específica ó número de especies (S) y la abundancia fue tomada como el número de eventos de captura para cada especie. Con el estimador no paramétrico Chao 2, calculado a través del programa Estimates 8.0 (Colwell 2006), se determinó la riqueza esperada y, a su vez, se estimaron los porcentajes de representatividad para los tres tipos de hábitats evaluados (general y discriminados por localidad). Se realizaron comparaciones de la acumulación de especies entre tipos de hábitats mediante análisis de rarefacción basada en muestras (Gotelli y Colwell 2001; Colwell et al. 2004). Se compararon los tres tipos de hábitats de manera general en términos de riqueza observada, abundancia y riqueza estimada mediante pruebas t. Para estas pruebas se utilizó el programa Statistica versión 7 (StatSoft 2007). Con el fin de determinar el cambio en la composición de la comunidad de hormigas entre tipos de hábitats a nivel regional y teniendo en cuenta cada localidad, se utilizó el índice de Bray-Curtis, que se presentó en dendrogramas de similitud. Además se realizó un análisis de ordenamiento (Nonmetric Multidimensional Scaling, NMDS), para representar los datos de acuerdo con sus distancias, seguido de un análisis ANOSIM, para evidenciar relaciones entre tipos de hábitats. Para estos análisis se empleó el programa PAST versión 2.14 (Hammer et al. 2001).
Resultados y Discusión
Composición y diversidad
Las hormigas capturadas sumaron 9.624 individuos en 1.775 eventos de captura, de los cuales 1.123 individuos (269 eventos) correspondieron a cinco géneros y 17 especies y morfoespecies de hormigas cazadoras de las subfamilias Ectatomminae y Ponerinae (Tabla 2). Lo cual representa un bajo registro comparado con otros estudios en bosque seco (Arcila-Cardona et al. 2007; Chacón de Ulloa et al. 2007; Domínguez et al. 2007), bosques subandinos (Abadía et al. 2010) y sistemas productivos (Sanabria-Blandón y Chacón de Ulloa 2011). Teniendo en cuenta que las localidades evaluadas no sobrepasan los 660 m de altitud, este resultado puede estar relacionado con el patrón encontrado en los Andes como consecuencia de las variaciones en la humedad y la pluviosidad, en el que el número de especies de hormigas es mayor entre 900 y 1700 m de altitud y disminuye por debajo de 900 m y por encima de 1700 m de altitud (Van der Hammen y Ward 2005). De otra parte, algunos estudios sobre biogeografía de hormigas cazadoras en Colombia, sugieren que la baja diversidad y similitud de las regiones secas del Caribe y el valle interandino del Magdalena en comparación con otras regiones del país, se debe probablemente a la falta de información y al predominio de ecosistemas transformados, lo que finalmente empobrece y homogeniza la fauna de hormigas cazadoras (Lozano-Zambrano et al. 2007; Lozano-Zambrano y Fernández 2007).
Lista de las especies de hormigas cazadoras y eventos de captura en tres tipos de hábitats, Bosque (B), Matriz (M), Matorrales (Mt), en el bosque seco Tropical del norte del Tolima.
Teniendo en cuenta los trabajos de Fernández et al. (1996), Sarmiento (2000), Jiménez et al. (2007a), Reinoso et al. (2009), Yara-Ortiz (2008; 2011), en los que se reportan especies de hormigas cazadoras para el Tolima, el registro de este estudio representa el 40% de las subfamilias y el 44,7% de las especies de hormigas cazadoras reportadas para el departamento. De esta manera, el presente trabajo contribuye al enriquecimiento del inventario regional con nuevos registros de las especies Leptogenys pubiceps (Emery, 1890), Leptogenys ritae (Forel, 1899), Odontomachus ruginodis (Smith, 1937) y Pachycondyla verenae (Forel, 1922).
La riqueza varió en los tres tipos de hábitat, 13 especies de hormigas cazadoras se registraron en bosques, siete en la matriz adyacente y 10 en parches de matorrales. Se evidenciaron diferencias entre tipos de hábitats al realizar comparaciones entre pares de ellos (Tabla 3). La riqueza fue mayor en bosque, seguido de matorrales y matriz. Este resultado concuerda con lo encontrado tanto en bosque seco como en otras zonas de vida en los estudios de Dix (2005), Chacón de Ulloa et al. (2007) y Chaves et al. (2007) en los que el número de especies de hormigas cazadoras registradas en los bosques representa más del 73% y alcanza un 94% del total encontrado, y en la matriz este valor no supera el 32%. A nivel local, también se observó variación en la riqueza observada (Tabla 2), sin embargo, al realizar comparaciones pareadas entre bosques y matrices correspondientes, la diferencia es significativa sólo en el CURN-UT, por lo que la diferencia hallada a nivel general puede estar determinada por esta localidad (Tabla 4). Es posible que la preparación del suelo en cada rotación de cultivo en las prácticas agrícolas en el CURN-UT, afecte la comunidad de hormigas cazadoras y, finalmente, algunas pocas especies logren establecerse y eludir este tipo de perturbación constante. En contraste, en las demás localidades, los cultivos son más estables en este sentido y permiten el establecimiento de un número no tan desigual de especies en relación con el bosque.
Comparación t para muestras independientes de la riqueza observada, eventos de captura (abundancia) y riqueza estimada entre tipos de hábitat, bosque (B), matriz (M) y matorrales (Mt), en el bosque seco tropical del norte del Tolima.
Comparación t-pareada de la riqueza y abundancia de hormigas entre bosques y matrices respectivas en el bosque seco tropical del norte del Tolima.
Entre Ponerinae y Ectatomminae, la subfamilia Ponerinae fue la más representativa en número de especies (88,23%), con los géneros Hypoponera y Pachycondyla que aportan el 41,17 % у 23,53 % de las especies, respectivamente. Estos géneros resultan frecuentes ya que son algunos de los más grandes, comunes y representativos en los bosques neotropicales (Fernández 2003; Fernández y Arias 2007; Mackay et al. 2007).
Algunas especies se presentaron únicamente en un tipo de hábitat, en el bosque el número de estas especies fue dos veces mayor en relación a la matriz y a los parches de matorrales (Fig. 2). Las especies L. ritae, P. verenae, Pachycondyla impressa (Roger, 1861) e Hypoponera sp.7, se presentaron únicamente en bosques; Hypoponera sp.1 e Hypoponera sp. 2 sólo en matriz y finalmente, Hypoponera sp.3 y L. pubiceps, se recolectaron únicamente en parches de matorrales. Estas especies, presentan también una baja abundancia (0,37 - 1,49%, Tabla 2), por lo que podrían representar casos de rareza demográfica y de vulnerabilidad (Armbrecht y Ulloa-Chacón 1999; Chacón de Ulloa et al. 2007), principalmente aquellas especies propias de bosques debido al deterioro avanzado de hábitats con cobertura boscosa en el paisaje del norte del Tolima. Algunos estudios reportan a P. impressa y especies del género Leptogenys en hábitats abiertos y en sistemas productivos, sin embargo su captura es más frecuente en áreas con mayor cobertura arbórea (Arcila-Cardona et al. 2007; Chacón de Ulloa et al. 2007; Domínguez et al. 2007; Mackay et al. 2007; Abadía et al. 2010; Sanabria-Blandón y Chacón de Ulloa 2011). En contraste, P. verenae, presenta hábitos de nidificación específicos debido a que utiliza sustratos presentes en áreas boscosas (Mackay et al. 2007).
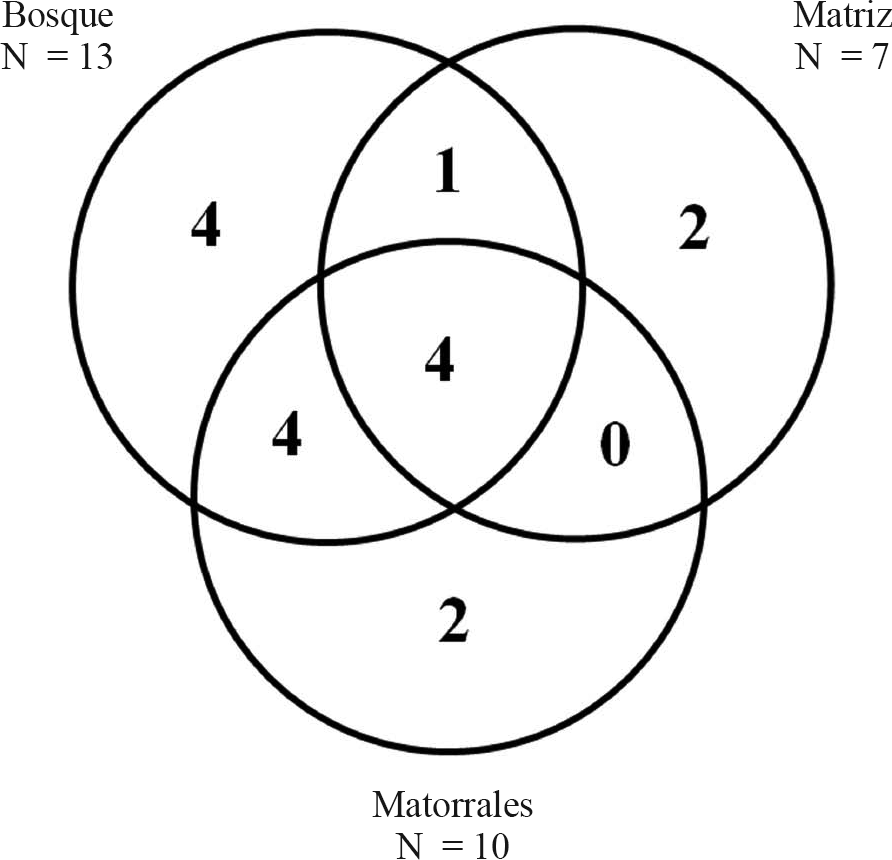
Número de especies exclusivas de hormigas de cada hábitat y compartidas entre ellos. Cada círculo representa un hábitat y los números, las especies de hormigas encontradas.
Estimación de riqueza por hábitats
El muestreo permitió obtener una representatividad de especies alta en los tres hábitats, superior al 70%. En general se logró el 93,46% y en los tipos de hábitats, los valores más altos se lograron en la matriz con el 100%, seguida de matorrales con 83,75% y, finalmente, en bosques con 75,71%. El estimador Chao 2 y las curvas de rarefacción regionales para los tres tipos de cobertura vegetal, reflejan diferencias entre la riqueza esperada y la tasa de acumulación de especies en cada tipo de hábitat, con valores más altos para bosques, seguidos por parches de matorrales y por último la matriz (Tabla 5, Fig. 3). La pérdida de especies desde el bosque hacia la matriz equivale al 59,23% de las especies esperadas. De acuerdo con lo anterior, se resalta el gran valor de los fragmentos de bosque y parches de matorrales, que pueden mantener una parte de la biodiversidad regional de hormigas cazadoras en el paisaje fragmentado del bosque seco del norte del Tolima y, por ende, su conservación debe ser parte fundamental en las estrategias para el mantenimiento de dicha biodiversidad.
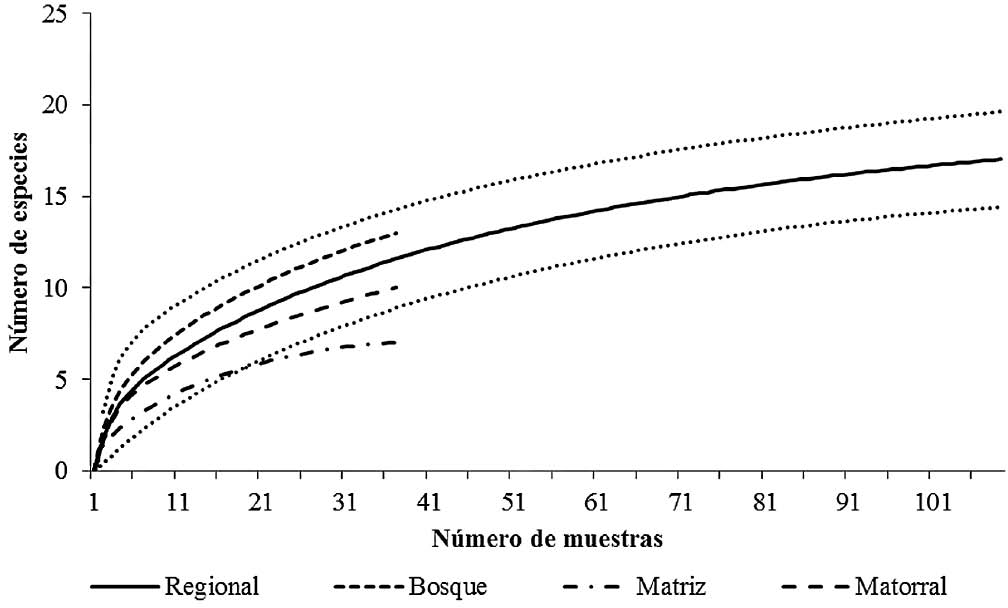
Curva de rarefacción regional (línea continua), intervalos de confianza (IC 95%, líneas discontinuas), y para las hormigas encontradas en bosque, matriz y parches de matorrales, en el bosque seco tropical del norte del Tolima.
Riqueza estimada general de hormigas para tres tipos de hábitats en el bosque seco tropical en el norte del Tolima.
Abundancia
En cuanto al número de eventos de captura en cada hábitat, no se halló diferencia significativa al compararlos de manera general (Tabla 3) como tampoco entre bosques y matrices correspondientes (Tabla 4). Sin embargo, es importante destacar a Ectatomma ruidum (Roger, 1860), Odontomachus cf. bauri (Emery, 1892) y Pachycondyla harpax (Fabricius, 1804), como las especies más comunes en los tres tipos de hábitats (Tabla 2), en las cuales se observan dos tendencias: O.cf. bauri y P. harpax aumentan su abundancia con la cobertura arbórea, mientras que en E. ruidum disminuye. A pesar de que las dos primeras especies han sido reportadas en hábitats boscosos, áreas abiertas, plantaciones y ambientes urbanos (Rodríguez 2007; Mackay et al. 2007), también O. bauri ha mostrado dependencia de hábitats con cobertura arbórea y hojarasca (Fontalvo-Rodríguez y Domínguez-Haydar 2009). Es posible que en el bosque seco del norte del Tolima, tanto O.cf. bauri como P. harpax prefieran hábitats con cobertura arbórea, ya que la disponibilidad de sitios de anidamiento y de alimento en estos tipos de cobertura es mayor que en hábitats abiertos, así como una menor competitividad (Torres 1984; Bustos y Ulloa-Chacón 1996). De este modo, O. cf. bauri y P. harpax fueron las especies más abundantes en bosque con 25,5% y 39,8% de las capturas, respectivamente. Por el contrario E. ruidum, encontrada tanto en bosques como en áreas abiertas (Chacón de Ulloa et al. 2007; Serna y Vergara-Navarro 2007; Fontalvo-Rodríguez y Domínguez-Haydar 2009), en esta región prefiere hábitats con poca cobertura arbórea, es una especie dominante en la matriz con el 77,9% de los eventos de captura y con una abundancia representativa (38,8% de los eventos) en parches de matorrales, similar a lo observado por (Fontalvo-Rodríguez y Domínguez-Haydar (2009). Sin embargo, es necesario tener en cuenta que su distribución en diversos tipos de hábitats puede variar con la zona de vida y las características del ambiente, ya que sus estrategias de alimentación, tamaño y carácter generalista permiten su adaptación a las alteraciones en su hábitat y, por ende, de la disponibilidad de los recursos (Santamaría et al. 2009a, 2009b).
De otra parte, de acuerdo con Halffter y Ezcurra (1992), las especies se distribuyen según jerarquías de abundancia como resultado de la abundancia relativa de cada una de ellas, por lo que es posible que la presencia de especies dominantes como E ruidum, O.cf. bauri y P.harpax en los tipos de hábitats evaluados estén determinando la estructura de la comunidad de hormigas cazadoras.
Composición y tipos de hábitats
Al analizar la composición de especies de hormigas cazadoras a través de agrupamiento usando el índice de Bray-Curtis, se encontró que la composición y estructura de esta comunidad en fragmentos de bosque y parches de matorrales son similares, pero a su vez se diferencian de la matriz. El nivel de similitud entre bosques y parches de matorrales es de 0,62, mientras que la matriz se asemeja a los elementos de cobertura arbórea con un valor de 0,51 (Fig. 4A). Es necesario mencionar que al encontrar una riqueza similar a nivel local entre los tipos de hábitat no se indica que la comunidad de hormigas cazadoras esté conformada por las mismas especies en cada uno de ellos. La composición de especies en cada hábitat y localidad no evidenció agrupación entre los bosques con su respectiva matriz, por lo que es posible que el impacto de sistemas agrícolas en esta región sea generalizado y esté limitando la movilidad de las especies de hormigas cazadoras desde los bosques hacia la matriz, contrario a lo encontrado por Domínguez et al. (2007). Sin embargo, en la mayoría de los estudios, se reporta que niveles más contrastantes de tipos de hábitats difieren marcadamente en relación a la composición de especies (Armbrecht y Ulloa-Chacón 1999; Carvalho y Vasconcelos 1999; Vasconcelos 1999; Vasconcelos et al. 2001; Arcila-Cardona et al. 2007). De esta manera es posible que el tipo de matriz influya sobre este resultado. En la Figura 4B se observan dos grupos que separan matrices de hábitats con cobertura arbórea (fragmentos de bosque y matorrales), al interior de los cuales se muestran similitudes mayores al 55%. Este tipo de agrupamiento coincide con el encontrado por Arcila-Cardona et al. (2007), al evaluar diferentes elementos del paisaje en el bosque seco de la cuenca alta del río Cauca. De otra parte, es posible que la agrupación de el parche de matorral ubicado en Guayabal (Mt3) con las matrices, sea el resultado de un paisaje más degradado en el que predominan pastizales expuestos con algunos árboles dispersos lo que podría hacer que este hábitat presente condiciones similares a las de las matrices.
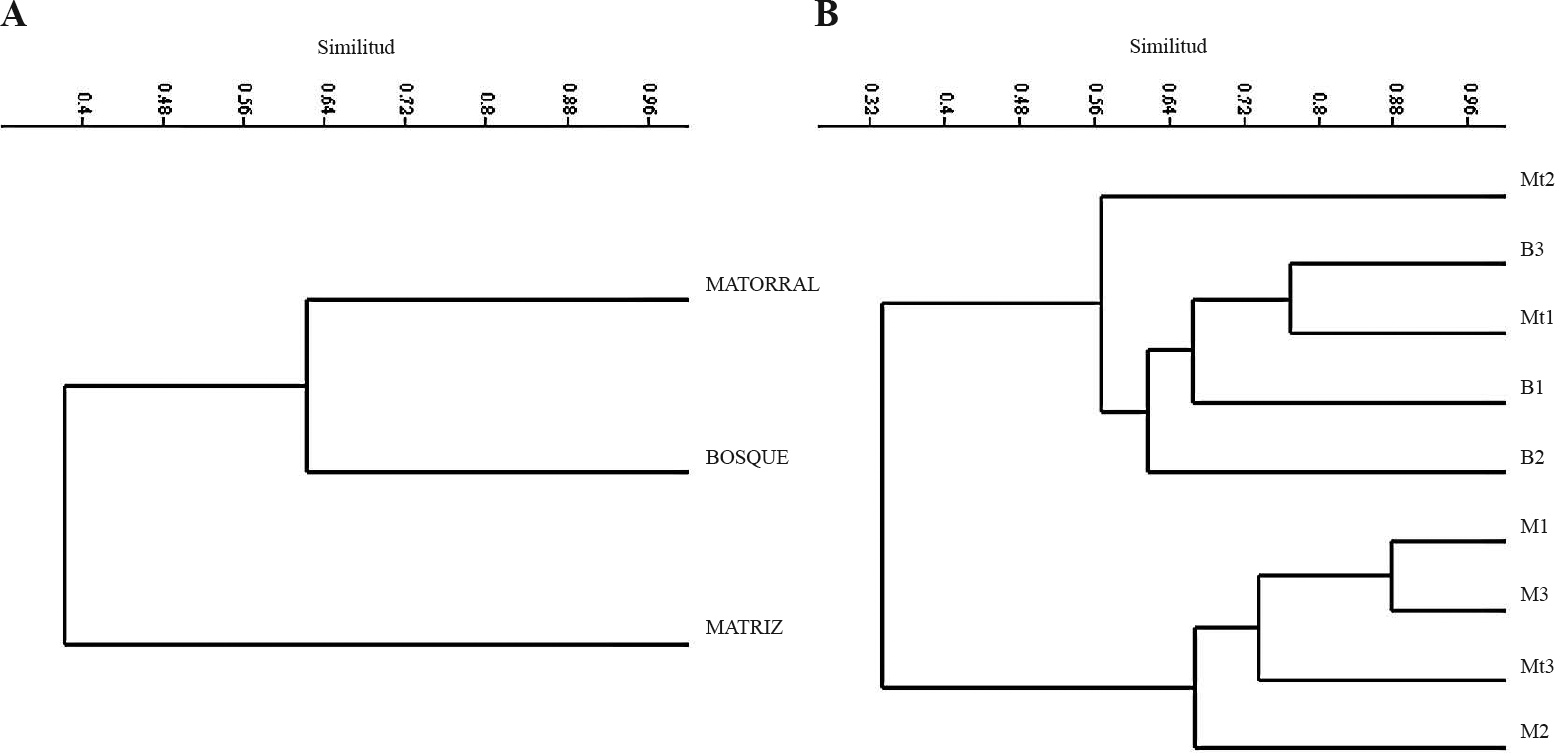
Dendrogramas de similitud de Bray-Curtis para las hormigas encontradas en el bosque seco tropical del norte del Tolima. A. Discriminado por hábitat. B. Discriminado por habitat y localidad. Bosque = B; Matriz = M; Matorrales Mt.
El análisis de ordenamiento (NMDS) en dos dimensiones permitió describir los datos de manera eficiente (stress = 0,062) y resultó en la agrupación de los tres tipos de hábitats. La Figura 5 presenta los tres grupos que corresponden a fragmentos de bosque, parches de matorrales y matrices, al tiempo que se observa cómo los hábitats bosque y matorrales son más similares entre sí comparados con la matriz, la cual formó un grupo aislado. El ANOSIM, mostró diferencias significativas entre los grupos de especies en cada tipo de hábitat (R = 0,61; P = 0,007). Esto confirma lo expuesto anteriormente mediante del índice de Bray-Curtis.
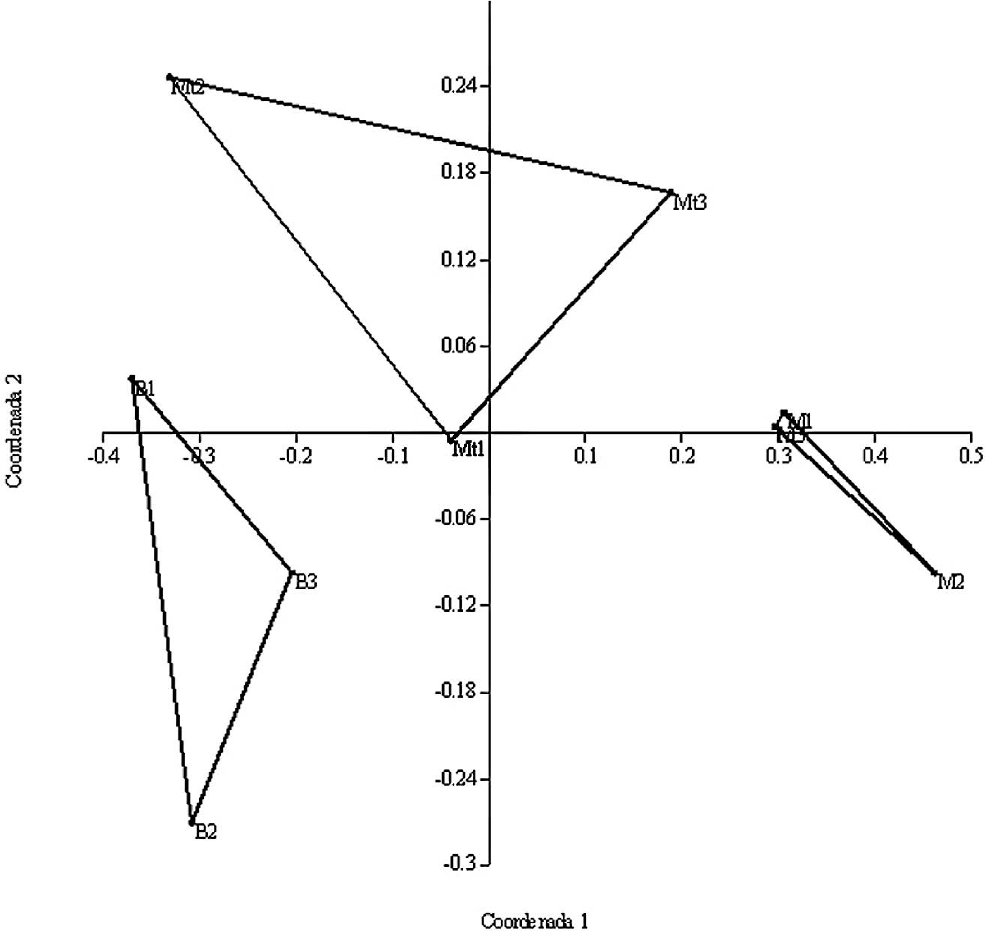
Ordenamiento de tres tipos de hábitats, bosque (B), matriz (M) y matorrales (Mt), discriminado por localidad, mediante NMDS usando el índice de Bray-Curtis.
La presencia de L. pubiceps cobra importancia ya que de acuerdo con Jiménez et al. (2007b), es una especie prioritaria por características como distribución en 1 ó 2 ecorregiones, en áreas transformadas por actividades humanas en un 71-100%, reportada en al menos un área protegida en el Sistema Nacional de Áreas Protegidas (SINAP) y aunque los autores registran que está asociada a bloques grandes de bosque (>1000 ha), en este estudio se registró en los matorrales de Guayabal (Mt3), hábitat que fue similar en composición a las matrices.
Finalmente, es necesario resaltar que a pesar de la distancia entre las ventanas y, por ende, los sitios evaluados, las similitudes son considerables en cuanto a la comunidad de hormigas cazadoras en los tipos de hábitats, especialmente las matrices, posiblemente por su homogeneidad.
Conclusiones
El bosque seco tropical del norte del Tolima presenta una fauna pobre de hormigas cazadoras en comparación con los registros para el Valle del Cauca y el Atlántico, sin embargo, alberga una parte importante de los registros para el departamento. Es posible que la transformación de hábitats naturales en áreas de uso agrícola desarrollada en la región, se constituya en un factor que modifica la estructura de las comunidades de hormigas cazadoras, ya que aunque en algunos lugares la riqueza sea similar en bosques y matrices, la composición de especies difiere entre ellos y de esta manera la comunidad responde a dicho cambio. En este sentido, los bosques y los parches de matorrales pueden comportarse como hábitats similares en términos de riqueza y composición y por lo tanto, adquieren mayor importancia de conservación, adicional a la presencia de especies consideradas como raras demográficamente, debido a que pueden presentar vulnerabilidad en la medida en que avanza el deterioro de los bosques, como consecuencia de la transformación de hábitats naturales.
Footnotes
Agradecimientos
Las autoras expresan sus agradecimientos al Fondo Regional de Investigaciones, Red de Universidades Públicas del Eje Cafetero Alma Mater, Programa Jóvenes Investigadores convocatoria regional AMFRI-JI-10. Al Comité Central de Investigaciones de la Universidad del Tolima y al Grupo de Investigación en Zoología de la Universidad del Tolima por su colaboración en la ejecución de este trabajo. Al director del CURN-UT, los dueños y trabajadores de las haciendas El Chaco y Guananí, a los auxiliares de campo por su colaboración y apoyo durante la fase de campo. Al revisor del abstract y, finalmente, a los evaluadores anónimos por las observaciones y sugerencias sobre el manuscrito.