
Abstract
An adequate agroecological management of ranching systems benefits from the associated entomafauna. The aim of this research was to determine the species of staphylinids in five production systems established along a gradient of plant recovery in the Colombian Andes. The hypothesis was that the diversity was positively associated with the recovery of the arboreal component. The survey was made over two rainy and one dry seasons between 2004 and 2006. Using pitfall traps, 272 beetles were collected, which belonged to five subfamilies and 45 morphospecies. The subfamilies Aleocharinae and Staphylininae were the most diverse, while Anotylus (Oxytelinae) was the most abundant. Pastures with trees exhibited high species richness, which was comparable to that of natural ecosystems such as forests and “guadua” (bamboo-like) forests. Homogeneous vegetation was highly related to a low diversity of staphylinids, and so the components of production ranching systems influence diversity. The different elements of the landscape, however, contribute as a whole to the regional diversity of staphylinids.
Introducción
La agricultura y ganadería reemplazan los bosques naturales por sistemas productivos abiertos implicando pérdida o alteraciones de la biodiversidad (Altieri y Nicholls 2002). En Latinoamérica, la ganadería corresponde al 82% de los agroecosistemas y se caracteriza por la baja diversidad vegetal representada por pasturas extensivas. Por ello se considera que un manejo ambiental adecuado, utilizando el componente arbustivo y arbóreo, puede favorecer la biodiversidad silvestre (Murgueitio et al. 2003).
Las prácticas agrícolas y ganaderas han transformado el paisaje de los Andes Colombianos en mosaicos complejos de potreros, campos agrícolas, fragmentos de bosque y otros tipos de cobertura arbórea (IAvH 1998). A pesar de que estos elementos del paisaje proporcionan una amplia diversidad de hábitats, generalmente son considerados de poco valor en conservación y escasamente son incluidos en estudios de biodiversidad (Fajardo et al. 2006). Estudios recientes indican que una considerable proporción de la biodiversidad original puede persistir dentro de dichos elementos del paisajes y que el grado de cobertura arbórea remanente juega un papel fundamental (Harvey et al. 2004). Sin embargo, la información sobre biodiversidad en paisajes agrícolas es escasa (Hernández et al. 2003) pero indudablemente útil para el diseño de planes de conservación eficaces y sostenibles.
Los insectos constituyen una porción muy importante en los agroecosistemas y bosques, tanto en abundancia como en diversidad. Dentro de los coleópteros, se destaca la familia Staphylinidae por ser altamente diversa y dominante en la fauna del suelo en muchos ecosistemas a nivel mundial (Obrtel 1968; Bohac 1999; Navarrete-Heredia et al. 2002). La mayoría de las especies son depredadoras de otros artrópodos, muchas son saprófagas o se alimentan de hongos o polen. Sólo pocas especies son de importancia económica actuando como enemigos de plagas de cultivos (Andersen 1999); por ejemplo, algunas especies del género Aleochara, quienes han sido reportadas como predadores de segundo orden sobre plagas en sistemas manejados (Haynes et al. 1980).
Los estafilínidos han sido propuestos como potenciales bioindicadores en zonas seminaturales y cultivadas en Europa Central (Bohac 1999). Presentan susceptibilidad a las prácticas de agricultura extensiva; la riqueza y diversidad de especies aumentan con la reducción de la labranza y poco uso de pesticidas, lo que permite el crecimiento de malezas y por tanto favorables condiciones microclimáticas para estos organismos (Krooss y Schaefer 1998). Asimismo, la distribución y abundancia de estafilínidos parece estar influenciada por la temperatura (Hoffman y Mason 2006) y la humedad (García 2001) del suelo. Además, se han encontrado diferencias a nivel de comunidad ligadas principalmente a las características estructurales de la vegetación (e.g. altura) (Hoffman y Mason 2006) y a la composición de la comunidad de plantas (Woodcock et al. 2005).
Colombia cuenta con al menos 796 especies de estafilínidos, lo que representa el 25% del potencial de su riqueza (Gutiérrez 2003; Newton et al. 2005). Los estudios sobre Staphylinidae en Colombia han sido desarrollados principalmente en bosques, desde altamente conservados en gradientes altitudinales en los Andes Occidentales (Gutiérrez-Chacón y Ulloa-Chacón 2006) hasta pequeños fragmentos de Bosque Seco (García et al. 2001; García y Chacón de Ulloa 2005) y sus matrices (Bermúdez et al. 2006) en el Sur Occidente del país. En general, se ha encontrado que los estafilínidos poseen estrechos rangos de distribución, exhibiendo altas tasas de recambio de especies a través del gradiente de elevación y entre localidades (Gutiérrez-Chacón y Ulloa-Chacón 2006) y que la diversidad de estos coleópteros se incrementa en elementos del paisaje con mayor heterogeneidad vegetal como bosques de galería y fragmentos de bosque (Bermúdez et al. 2006).
El objetivo de este estudio fue entonces determinar la composición y diversidad de estafilínidos en cinco sistemas productivos en la zona Andina Colombiana definidos con base en el uso de suelo en fincas ganaderas. Además se relaciona la biodiversidad con el grado de recuperación del componente arbóreo. Este trabajo constituye el primer estudio comparativo de la familia Staphylinidae en sistemas manejados en Colombia y es un aporte al conocimiento taxonómico de la entomofauna colombiana.
Materiales y Métodos
Área de estudio
El presente estudio se desarrolló en fincas ganaderas de cinco municipios en los departamentos de Quindío y Valle del Cauca, Colombia (Tabla 1, Fig. 1). Dichas fincas están incluidas en el proyecto: “Enfoques Silvopastoriles Integrados para el Manejo de Ecosistemas” (Rivera 2005).
Mapa de ubicación geográfica del área de estudio, Departamento del Quindío y Valle del Cauca, Colombia. Fuente: CIPAV (2003).
Localidades Muestreadas y descripción de la vegetación presente en cada uso de tierra.
Diseño del muestreo
Se seleccionaron cinco sistemas productivos con base en el uso de tierra y el orden creciente de recuperación vegetal. De cada sistema se escogieron tres lotes, cada uno considerado como pseudoréplica (Tabla 1). En cada una de las pseudoréplicas se trazó un transecto de 100 m donde se instalaron 10 trampas de caída o Pitfall (equidistantemente) (Vásquez 2008) y se dejaron actuar por 24 horas. Se usó este tiempo porque en un estudio piloto, usando 24, 48 y 76 horas, el más eficiente en términos de relación costo-beneficio fue 24 h. Cada trampa consistió en un vaso plástico de 6 cm x 6 cm semilleno con agua jabonosa y enterrado a ras de suelo. En total se realizaron tres muestreos: el primero entre noviembre y diciembre de 2004, el segundo entre mayo y julio de 2005 y el tercero entre diciembre de 2005 y enero de 2006.
Determinación taxonómica
Los estafilínidos se separaron de los demás artrópodos y se montaron en seco en el laboratorio de Entomología, Universidad del Valle. La determinación taxonómica se realizó a nivel de género (a excepción de Aleocharinae) siguiendo las claves de Navarrete-Heredia et al. (2002) y García (2004) (Tabla 2). La colección resultante fue depositada en el Museo de Entomología de la Universidad del Valle (MEUV).
Especies de la familia Staphylinidae coleccionadas en cinco usos de tierra en el Valle del Cauca y Quindío. Pm-a: Pastura sin árboles, Slv: Silvopastoril, Pm+a: Pastura con árboles, Gd: Guadual, Bs: Bosque.
Análisis de datos
Para evaluar la efectividad de los muestreos en cada uso de la tierra, se realizaron curvas de acumulación de especies con el programa EstimateS (Colwell 2000) utilizando los estimadores no paramétricos ICE y Jack 2 (Colwell y Coddington 1994; Villareal et al. 2004). Se calcularon índices de diversidad de Shannon-Wienner (H') de acuerdo con la ecuación H'= -ΣPilnPi, donde Pi= (ni/N), siendo n = No. de individuos de la especie i y N = No. total de individuos en el sistema i; el índice de Equitatividad (J'), se estimó de acuerdo con J=Hobs-(Hmin/Hmax)-Hmin, cada índice se cálculo para cada localidad (Moreno 2001). Se calcularon índices de similitud de Jaccard, que se estima en función de los datos presencia-ausencia, así: Cj = c/(a+b-c), donde, a = No. de especies en sistema i, b = No. de especies en el sistema j, c = No. de especies comunes entre los usos de la tierra con base en el número de especies en común (Moreno 2001). Para determinar la distribución de la abundancia de las especies en cada uno de los usos de la tierra se construyeron curvas de dominancia-diversidad usando la formula pi*ln(pi) (Magurran 1987).
Resultados
Se capturaron 272 especímenes de la familia Staphylinidae agrupados en cinco subfamilias y 45 morfoespecies. Las subfamilias Aleocharinae y Staphylininae presentaron la mayor riqueza con 21 y 10 morfoespecies respectivamente, valores que representan más del 50% del total, seguidas por Paederinae (5), Tachyporinae (5), Oxytelinae (4). (Tabla 2).
El sistema de pastura con árboles presentó la mayor riqueza y diversidad de especies seguido por el guadual. El uso de la tierra con menor riqueza fue la pastura sin árboles (Tabla 3). En cuanto a la distribución de abundancia de especies, el guadual fue el más equitativo mientras que la pastura sin árboles y el silvopastoril fueron los usos de tierra menos equitativos. Las curvas de acumulación de especies con los dos estimadores (ICE y Jack 2) son consistentes entre si indicando que el bosque fue el uso de la tierra donde se recolectó una menor proporción de las especies esperadas (35-44%), mientras que en la pastura con árboles las especies colectadas representaron entre el 57 y 61% de las especies esperadas (Tabla 3; Fig. 2).
Riqueza observada y esperada con el estimador ICE, índice de diversidad de Shannon-Wienner (H΄) y equitabilidad de Pielou (J).
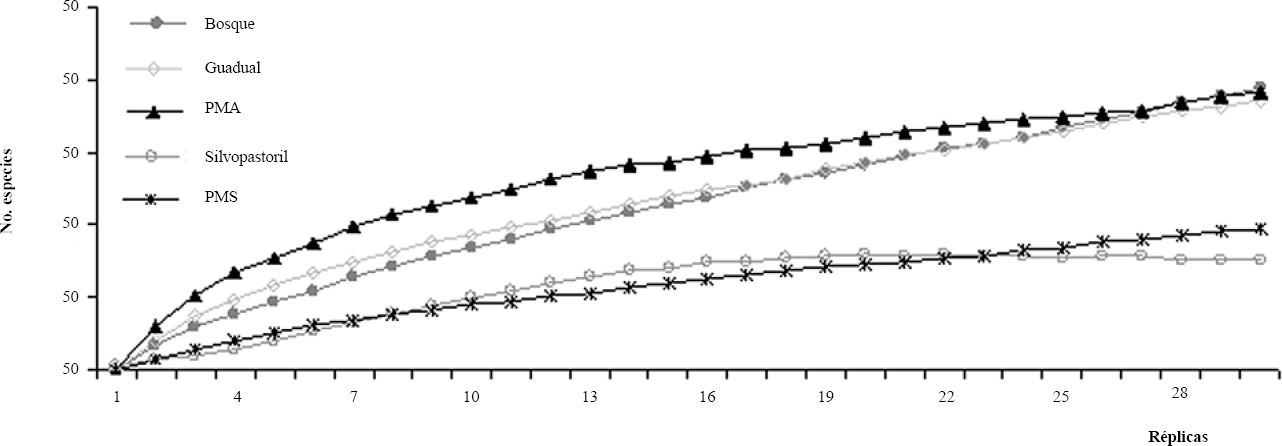
Curvas de acumulación de especies para todos los usos de suelo muestreados mediante el estimador no paramétrico Jack 2.
Los usos de la tierra fueron altamente disímiles entre si, con valores menores a 0,3 (Tabla 4). La menor similitud se presentó entre el bosque vs. pastura sin árboles y el bosque vs. silvopastoril, es decir entre el sistema natural y los usos de la tierra más intensivos (Tabla 4). Las curvas de dominancia-diversidad para todos los usos de la tierra (excepto el silvopastoril) mostraron que tres especies dominan numéricamente. Las pendientes son similares y abruptas, indicando la dominancia de unas pocas morfoespecies y muchas especies de aparición única. En estos casos, las morfoespecies dominantes pertenecen principalmente a las subfamilias Aleocharinae (Aleocharinae sp. 19 y Aleocharinae sp. 14) y Oxytelinae (género Anotylus) (Fig. 3).
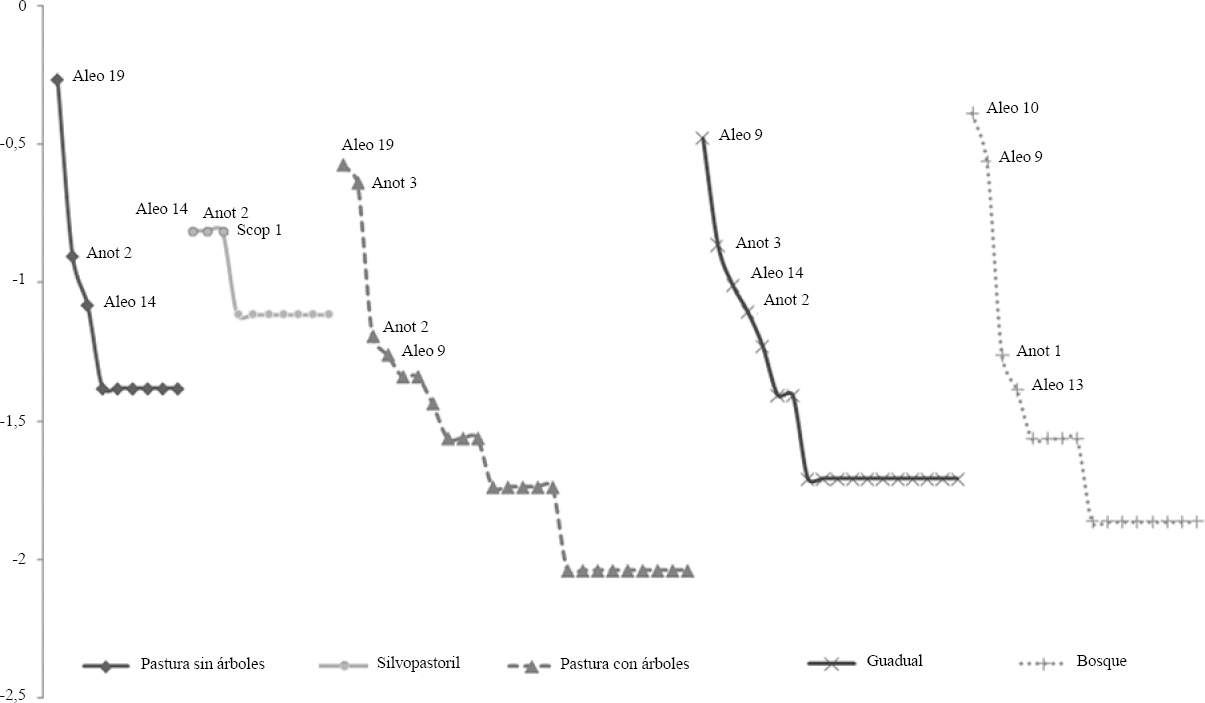
Relación de dominancia-diversidad de estafilínidos en cinco usos de tierra en el Valle del Cauca y Quindío-Colombia. Aleo: Aleocharinae; Anot: Anotylus; Scop: Scopaeus. Los códigos se refieren a las morfoespecies correspondientes en la tabla 2.
Índice de similitud de Jaccard entre sitios.
Discusión
La proporción de cada subfamilia dentro del estudio refleja en parte su tamaño dentro de la familia Staphylinidae a nivel global. Las dos subfamilias mejor representadas fueron Aleocharinae seguida por Staphylininae cuyos miembros son considerados generalistas y depredadores dominantes en la fauna del suelo (Navarrete-Heredia et al. 2002), esto contrasta con los estudios en paisajes fragmentados del Bosque Seco Tropical del Valle del Cauca y Quindío (García et al. 2001; García y Chacón de Ulloa 2005), en donde se registraron 78 especies agrupadas en ocho subfamilias, de las cuales Paederinae, Staphylininae y Aleocharinae fueron las de mayor riqueza específica. De manera similar, los resultados obtenidos en el estudio de Gutiérrez-Chacón y Ulloa-Chacón (2006), en tres localidades de la cordillera occidental, se registraron 11 subfamilias y 293 morfoespecies encontrándose una proporción similar, pero en este último caso la subfamilia Pselaphinae estuvo en segundo lugar de riqueza debido al tipo de muestreo (sacos Winkler), con los cuales se recolecta una mayor proporción de artrópodos de hojarasca (Olson 1994). Los bajos valores de especies observadas en relación con las especies esperadas podrían atribuirse a que en este estudio se utilizaron exclusivamente trampas de caída, las cuales tienden a excluir animales con poco movimiento y generalmente submuestrean depredadores que cazan al acecho y aquellas especies que pueden sostenerse en superficies verticales (Longino et al. 2002). Longino y colegas (2002), en Costa Rica, utilizaron cebos, fumigación de dosel, trampas Malaise muestras Berlese y Winkler y captura manual y encontraron 437 especies de hormigas con un inventario casi completo de acuerdo con los estimadores de riqueza. Por tanto, es recomendable complementar diferentes métodos de muestreo dirigidos a los diversos sustratos donde potencialmente habitan estos coleópteros. Al no tener en cuenta un método especializado en artrópodos de hojarasca (e.g sacos Winkler) se excluye la recolección de grupos altamente diversos pero de limitada movilidad como Pselaphinae, cuya representación en otros estudios (e.g. Bermúdez et al. 2006; Gutiérrez-Chacón y Ulloa-Chacón 2006) ha sido sustancial.
Diferentes entidades de Aleocharinae (e.g. Aleocharinae sp.19, Aleocharinae sp. 14) y Anotylus spp fueron los que exhibieron dominancia en todos los usos. En un estudio que evaluó el impacto de los cultivos en los artrópodos del suelo, Holland y Reynolds (2003), encontraron que de los coleópteros recolectados, el 10% perteneció a la familia Staphylinidae y que de este porcentaje, el 72% correspondió a individuos del género Anotylus, consistente con el resultado de este estudio donde Anotylus fue el más abundante. Se esperaba que la biodiversidad se incrementara en relación con la recuperación del componente vegetal, sin embargo, a pesar de que el bosque presenta la mayor cobertura y diversidad vegetal respecto a los demás usos de la tierra, éste exhibió valores de diversidad relativamente bajos, posiblemente como consecuencia del reducido tamaño de los fragmentos, el aislamiento y el alto grado de intervención al que son sometidos (Kattán 2002). Por otro lado, los bosques en general, aún perturbados, presentan alta diversidad vegetal, por lo cual las relaciones ecológicas se tornan complejas, pueden entrar a ser importantes los efectos indirectos y otros factores que podrían limitar o estimular la diversidad de estafilínidos.
Adicionalmente, en los bosques, la proporción de especies recolectadas en relación a las especies esperadas fue la más baja en comparación con los otros usos de la tierra, lo cual dificulta una comparación equiparable en términos de diversidad y riqueza. En general, la eficiencia del muestreo fue mayor en los sistemas con baja heterogeneidad vegetal (silvopastoril y pastura sin árboles) respecto a aquellos con mayor componente arbóreo (bosque, guadual y pastura con árboles) (Tabla 1). Este resultado es razonable en tanto que se espera mayor diversidad en los bosques, que generalmente son más heterogéneos que los sistemas productivos.
Los bajos índices de similitud de Jaccard en general reflejan el carácter más bien exclusivo o singular la fauna en cada uso de la tierra, demostrando que los diferentes elementos del paisaje aportan fuertemente a la diversidad regional de estafilínidos. Los índices que presentaron valores más bajos correspondieron al bosque en comparación con los sistemas ganaderos de manejo intensivo como lo son el silvopastoril y la pastura sin árboles (Sanabria 2006). Estos últimos tienen una menor cobertura arbórea y su fauna es muy diferente de aquella en sistemas más arbolados como el bosque y el guadual. Mientras que la cobertura de dosel del guadual y bosque puede alcanzar 15-20 m o más de altura con este dosel generalmente cerrado a estas alturas, la cobertura del sistema silvopastoril intensivo con Leucaena alcanza máximo 2,5 m, pues el ganado forrajea directamente sobre él. La pastura con árboles alcanza alturas similares a las del bosque pero las copas no se unen en la zona de dosel. Estas diferencias cualitativas, sumadas a los recursos que ofrecen los diferente sistemas (alimenticios, de refugio, de hojarasca, entre otros) podrían influir en que la composición de los estafilínidos difiera en los sistemas de uso de la tierra.
En Colombia los trabajos realizados con la familia Staphylinidae se han concentrado en ecosistemas naturales como bosques. Este estudio se realizó en un paisaje heterogéneo de los Andes colombianos, involucrando usos de la tierra con diferentes intensidades de manejo antrópico, enfocadas principalmente a la ganadería. Teniendo en cuenta que el muestreo se realizó en sistemas altamente intervenidos y que los estimadores indican baja representatividad del muestreo en algunos casos, se puede afirmar que los agroecosistemas del paisaje ganadero de esta región aportan alta diversidad de fauna asociada (e.g. estafilínidos) y que este estudio es una contribución al conocimiento a la diversidad de estos sistemas. Los resultados demuestran que de los sistemas ganaderos, la pastura con árboles es el más favorable para la conservación de la fauna de estafilínidos. Este hallazgo es consistente con el comportamiento observado en hormigas de suelo, en las cuales se encontró una correlación positiva altamente significativa con la cobertura vegetal (Rivera 2005). Por tanto, se resalta especialmente que el sistema productivo con elementos arbóreos presenta riquezas comparables con ecosistemas naturales como bosques y guaduales.
Agradecimientos
Las autoras agradecen a Leonardo Rivera, María Botero y Selene Escobar quienes contribuyeron con apoyo logístico, trabajo de campo y separación de muestras. A la fundación CIPAV-Cali, por proporcionar la financiación para el trabajo de campo, la Universidad del Valle, Facultad de Ciencias y al Grupo de Investigación en Biología, Ecología y Manejo de Hormigas, para el trabajo de laboratorio y logística.