
Abstract
In order to determine the behavior of cocoa hybrids to the attack of Steirastoma breve in its larval phase, the evaluation of damage intensity (DI) was carried out in a trial set up at the Padrón field experimental station in Miranda state, Venezuela. Sixty hybrid progenies were used, obtained through crossings of 27 parentals selected from among “criollo", "trinitario” and “forastero” types. The hybrids were planted in a randomized block design with six plants and three to ten replicates, depending on the availability of material. In the first evaluation the damage produced by larvae of the insect was recorded according to the presence or absence of affected hybrids. In the second evaluation a nominal scale from one to five was used to measure the DI in 57 hybrid progenies. Highly significant differences were observed (P <0.001; Kruskal-Wallis test) in DI among hybrids in the second evaluation. Multiple comparisons of the average ranges separated the hybrids into five groups, with DI extremes from one (11 hybrids) and between three and four (five hybrids that were the most susceptible); the rest formed intermediate groups. With the exception of the materials (PV-1 x IMC-11) x OC-61 and SJU-4 x (PV-1 x IMC-11), most of the hybrids that did not present damages tended to have a genotype of Amazonian origin. The results indicate a differential behavior of the evaluated hybrids to insect attack.
Introducción
La "gota" del cacao, Steirastoma breve (Sulzer, 1776) (Coleoptera: Cerambycidae) (Fig. 1D) es una de las principales plagas del cultivo en algunos países productores de este rubro. Su presencia ha sido señalada en el neotrópico desde Argentina hasta Venezuela en América del Sur, y en las islas Trinidad y Tobago, Grenada, Martinica, Puerto Rico, Jamaica y otros países (Entwistle 1972; Sánchez y Capriles de Reyes 1979). Las larvas y los adultos de este insecto, producen graves daños en tronco y ramas. Al emerger la larva se alimenta de la corteza y capas del cambium vascular. A medida que avanza este proceso, construyen galerías irregulares en espiral que pueden ocasionar el anillado de tallos y ramas, lesión que causa la muerte del área apical afectada o toda la planta según sea la edad y ubicación del daño (Entwistle 1972).
Por otro lado, existe poca información de evaluaciones de daño de este insecto en materiales de cacao provenientes de selecciones de este cultivo. La mayor parte de la información está referida a la resistencia de los materiales genotípicos de cacao a los principales agentes causales de enfermedades, como Crinipellis perniciosa (Stahel) Singer, Ceratocystis fimbriata Ellis y Halsted, Phytophthora palmivora (Butler) Butler (Desrosiers et al. 1955; Delgado y Echandi 1965; Capriles de Reyes et al. 1966; Capriles de Reyes y Reyes 1968; Esquivel 1973; Fonseca y Albuquerque 1996). Con relación a los insectos, se han registrado algunos clones de cacao con cierto grado de resistencia a Conopomorpha cramerella (Snellen, 1904) (Lepidoptera: Gracillariidae), Glenea aluensis Gahan, 1897 (Coleoptera: Cerambycidae) y S. breve, en Malasia, Guinea y Trinidad y Tobago, respectivamente (Azhar et al.1995; Azhar 2001; Efron y Epaina 2003; Maharaj et al. 2004).
En 1997 el Instituto Nacional de Investigaciones Agrícolas (INIA), Estación Experimental del Estado Miranda, se insertó en el Proyecto Internacional “Utilización y Conservación del Germoplasma de Cacao: Un enfoque Global" (Eskes 1999) el cual se afianza en tres criterios fundamentales: (1) selección de materiales nacionales de buena productividad y calidad (sobresalientes índices de mazorca y de almendra), (2) introducción de materiales foráneos y (3) evaluación de los materiales seleccionados e introducidos y los cruces entre ellos. La evaluación se basa en los aspectos agronómicos, fenotípicos y fitosanitarios. En este último aspecto se estudia el efecto de las principales enfermedades e insectos plaga del cacao en el país. El presente trabajo, tiene por objeto evaluar el comportamiento de 60 familias híbridas de cacao, generadas a partir de cruces entre materiales nacionales e introducidos al ataque de S. breve en su fase larval.
Materiales y Métodos
El trabajo se realizó en el Campo Experimental Padrón, adscrito al INIA-Miranda, ubicado en el sector Tapipa, Municipio Acevedo, estado Miranda, Venezuela, área cacaotera de bosque húmedo tropical según las zonas de vida de Holdridge (1978), con coordenadas geográficas 10°13′36″N, 66°18′30″W y una altitud de 20 msnm. El ensayo se instaló en el año 2000. Para ello se utilizó un diseño en bloques al azar, con 60 familias híbridas como tratamientos, generadas a partir del cruce de 27 clones parentales: 15 materiales nacionales y 12 introducidos, entre criollos, trinitarios y forasteros (Tabla 1), con unidad experimental de seis plantas por híbrido, de tres a diez repeticiones, según la disponibilidad de material de siembra, y una distancia de siembra de 3 x 3 m. Se usó como sombra permanente el bucare anauco (Erythrina glauca Willd.) (Leguminosae) a una distancia de 21 x 21 m, y como sombrío temporal el topocho pelipita (Musa sp. Grupo ABB subgrupo Topocho) (Musaceae) sembrado a una distancia de 3 x 3 m.
Clones parentales de cacao utilizados en los cruces para la obtención de las progenies híbridas.
Son materiales nacionales; algunos son derivados de cruces entre clones realizados y evaluados hace varios años en Venezuela.
La primera evaluación se realizó en el año 2002 mediante el uso presencia ó ausencia de plantas dañadas por larvas del insecto. La segunda evaluación se realizó en el 2003 para 57 híbridos y se empleó la siguiente escala de intensidad de daño (ID):
1 = Planta aparentemente sana.
2 = Ligeramente dañada (primeros síntomas de daño larval con presencia de sustancia gelatinosa).
3 = Moderadamente dañada (presencia de larvas y perforaciones o galerías en la mitad del perímetro alrededor del tallo o parte afectada) (Fig. 1A).
4 = Altamente dañada (presencia de larvas y perforaciones o galerías en casi todo el perímetro alrededor del tallo o parte afectada) (Fig. 1B).
5 = Extremadamente dañada (parte afectada totalmente anillada, región apical de la parte afectada muerta) (Fig. 1C).
Todas las evaluaciones se realizaron entre los meses de junio y julio. Los análisis estadísticos se realizaron mediante la prueba binomial y la prueba de Kruskal-Wallis (Siegel y Castellan 2003), equivalente a un ANAVA en diseños completamente aleatorizados pero cuando existe desigualdad en el número de repeticiones por tratamiento.

Tipos de daño ocasionado por S. breve. (A, B, C) fase de larva e intensidad de daño nivel tres, cuatro y cinco, según escala propuesta en plantas de cacao. (D) insectos adultos de S. breve.
Resultados y Discusión
En total, 42 progenies híbridas presentaron daños de larvas de S. breve, mientras que 18 de ellas (30% de los híbridos) no presentaron daños, siendo estas diferencias significativas (P ≤ 0,0015; prueba Binomial) entre presencia o ausencia de daño en las plantas híbridas de cacao. El mayor número de híbridos atacados mantuvo un rango de 0,1 a 14,9% (34 híbridos), los materiales más afectados oscilaron entre 15 y 29,9% (ocho híbridos; Fig. 2). Los híbridos más afectados fueron el SJU-3 x EEM-001, SJU-4 x OC-61, SJU-4 x EEM-001, EEM-001 x OC-61 y EEM-001 x ICS-6.
Se detectaron diferencias significativas entre los híbridos en la intensidad de daño (H=113,6; P < 0,001; Tabla 2). La comparación múltiple separa claramente cinco grandes grupos: extremos A y C con valores de mediana de 1 y 3 ≤ x ≤ 4, respectivamente y tres grupos intermedios no diferenciados, con valores de mediana entre uno y tres, inclusive. Solo 11 materiales genéticos presentaron valores de tres a cuatro en intensidad de daño y cinco de ellos representa al grupo C, que son los híbridos más atractivos para el insecto estudiado (Fig. 3; Tabla 3).
Comparación múltiple en la intensidad de daño de S. breve en híbridos de cacao.
Filas con la misma letra no presentaron diferencias significativas (P<0,05).
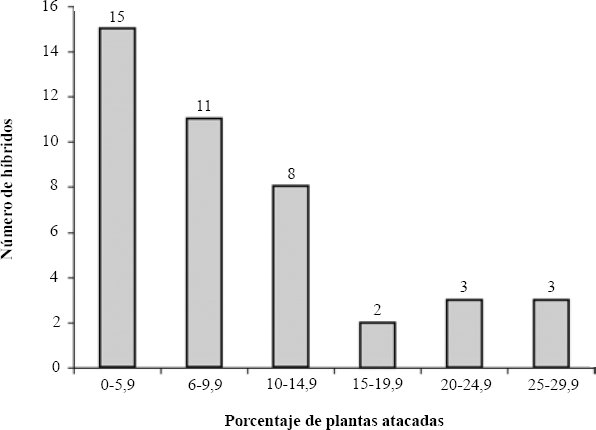
Porcentaje de plantas atacadas en la primera evaluación. Año 2002.
Al discernir el comportamiento de las progenies híbridas se observa una alta variabilidad que puede ser debida a la complejidad de los cruces (Tabla 3). Los materiales más susceptibles de hospedar larvas del insecto fueron los híbridos SJU-3 x EEM-001, SJU-4 x OC-61, SJU-7 x EET-332, (PV-1 x IMC-11) x EEM-001, (PV-1 x IMC-11) x OC-61 (todos con un componente de cacao tipo criollo en calidad de madre) con una ID de 3 (Tabla 3).
Intensidad de daño (ID) causada por S. breve en las progenies híbridas de cruces entre 27 materiales genéticos de cacao.
Valores de intensidad de daño (ID) referidos a la mediana en cada familia híbrida.
CM: Comparación múltiple. Letras distintas indican diferencias significativas (P < 0,05). NI: No Incluido en la evaluación.
Sin embargo, de 11 progenies híbridas que no presentaron daños, seis provienen del cruce de cacao criollo puro con materiales conocidos como trinitarios o forasteros: SJU-3 x ICS-6, ICS-6 x SJU-3, SJU-4 x (PV-1 x IMC-11), SJU-3 x (OC-77 x EET-400), EET-400 x PORCELANA VERDE, La Concepción-164 x SJU-3. Los otros cinco materiales están relacionados con el cacao de origen amazónico. Al parecer la posible resistencia o tolerancia al insecto es independiente del origen del tipo de cacao parental que participa en los cruces.
Se ha registrado que la presencia de polifenoles y otras sustancias como el ácido clorogénico, actúan como el principal mecanismo de resistencia en plantas de cacao para ciertas enfermedades fúngicas (C. fimbriata y C. perniciosa), y que la concentración aumenta a medida que aumenta la resistencia de los diferentes clones a tales enfermedades (Capriles de Reyes et al. 1966; Capriles de Reyes y Reyes 1968). Efron y Epaina (2003) encontraron una respuesta diferencial de materiales genéticos de cacao al ataque del cerambícido G. aluensis, en clones provenientes de cruces entre plantas de origen Trinitario, por ejemplo, clones derivados del cruce de los clones K-20 y KA2-101, muestran el más bajo promedio de daño (6,0) mientras que el más alto promedio de daño (31,8) fue obtenido en clones provenientes del cruce entre los Trinitarios KT-140 x KA5-201. Similares resultados reportan en Trinidad y Tobago con el insecto S. breve (Maharaj et al. 2004), donde ciertos clones locales como TSH-1102, 1188 y 1364 son los más susceptibles, mientras que los clones TSH-730, 1077, 1313, 1344, 1350 y 1362 son resistentes. Es probable que el mecanismo de resistencia o tolerancia de las progenies de cacao en el presente estudio se encuentre bajo control genético, asociado a sustancias o metabolitos producto de la defensa química de las plantas. Es necesario profundizar los estudios y evaluar las sustancias químicas y derivados presentes en las familias híbridas del bioensayo, según los parentales involucrados en los cruces, de tal manera que se pueda dilucidar porqué unos híbridos expresan comportamiento de resistencia y otros parecieran ser más susceptibles al insecto, aún con parentales de un mismo origen o tipo genético.
Los resultados indican que existe un comportamiento diferencial de los híbridos que al parecer es independiente del origen genético de los padres. El carácter criollo, trinitario o forastero en determinados cruces, aumenta considerablemente la preferencia de S. breve por hospederos que favorecen la oviposición, el desarrollo y el crecimiento de la fase larval del insecto.
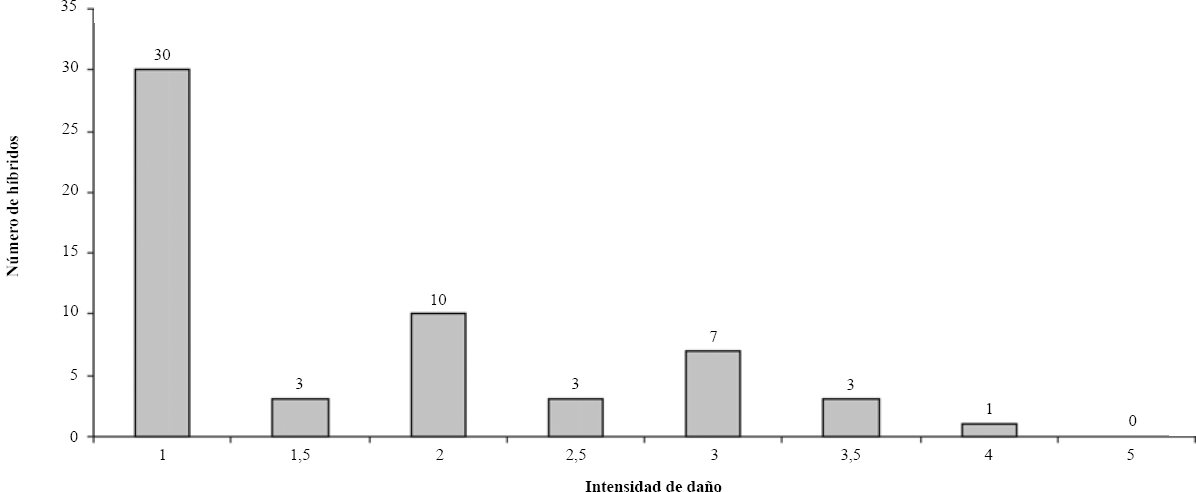
Frecuencias de la intensidad de daño de S. breve en híbridos de cacao. Valores relativos a la mediana. Año 2003.
Footnotes
Agradecimientos
Los autores desean expresar su más sincero agradecimiento a las instituciones CFC/ICCO/IPGRI (ahora BIOVERSITY) por el financiamiento de la presente investigación a través del Proyecto "Cocoa germoplasm utilization and conservation: a global approach". También desean agradecer a Bertus Eskes (CIRAD), Ventura González (INIA) por el minucioso trabajo de revisión del manuscrito y Carlos Marín (INIA) por la revisión de los análisis estadísticos.